同域分布水鹿和毛冠鹿活动节律的比较研究
——基于红外相机数据
2022-05-07贾国清杨旭李永东王宇郑笑傲蒋勇杨彪李生强
贾国清, 杨旭, 李永东, 王宇, 郑笑傲, 蒋勇, 杨彪, 李生强
1. 四川贡嘎山国家级自然保护区管理局,四川 康定 626000;
2. 成都兴艾信息技术有限公司,四川 成都 610051;
3. 西华师范大学,西南野生动植物资源保护教育部重点实验室,四川 南充 637002;
4. 四川省大熊猫科学研究院,四川 成都 610057;
5. 广西师范大学,广西珍稀濒危动物生态学重点实验室,广西 桂林 541004
同域物种的共存机制研究一直是群落生态学中的重要内容,开展同域物种的比较研究有助于进一步分析种间行为、生态适应与共存机制等[1]。根据生态位理论,同域分布且生态需求相近的物种,需要通过生态位分化才可有效减少或避免有限资源条件下的竞争[2],且必须通过一定的生态位分化才可达到稳定共存[3]。生态位分化往往涉及栖息地利用、食性、行为节律等多个维度[4],尤其对于同域分布的近缘物种上表现得更为明显,由于具有相似的身体结构、行为活动和资源需求等,往往成为潜在的竞争者[5-6]。因此,同域物种要想实现长期共存,往往需要调节各自的栖息地利用模式、食性选择与活动节律以促进生态位分化[7]。
动物的活动节律是行为生态学研究中的重要内容,其中日活动节律是动物应对昼夜更替(包括温度、湿度、光照强度等)长期适应而逐渐形成的在一天中不同时间段下的活动强度和变化规律[8]。目前,关于活动节律在同域物种共存机制中的作用研究已有不少文献报道,包括同域分布的地栖性雉类[4,9-10]、有蹄类[6,11]、食肉类[12,13]、啮齿类[14-15]等方面,研究表明同域分布且生态需求相近的物种可以通过调节活动节律来减少资源竞争从而达到共存。
作为一种新型的野生动物监测技术,红外相机技术自上世纪90年代应用以来,经过近三十年的发展,目前已广泛应用到野生动物生态学与保护学研究中[16-17]。近年来,利用该技术来探究野生动物活动节律和生态位分化的研究已有不少报道,相关应用也证实了该技术可以作为分析同域物种共存机制的有效技术途径[6,9-11,13-15]。
水鹿(Cervus equinus)隶属偶蹄目(Artiodactyla),鹿科(Cervidae),鹿属[18],在我国广泛分布于南部和西南部热带及亚热带地区,包括云南、四川、重庆、青海、贵州、广西、海南、湖南、广东、江西、台湾等地[19],被IUCN濒危物种红色名录列为易危(Vulnerable, VU)等级[20],属于国家二级重点保护野生动物[21]。毛冠鹿(Elaphodus cephalophus)隶属偶蹄目,鹿科,毛冠鹿属[18],广布于我国南部并延伸至缅甸,在我国的分布区包括湖南、浙江、安徽、福建、江西、湖北、广东、广西、四川、贵州、云南、西藏、陕西、甘肃、青海、重庆等地[19],被列为近危(Near Threatened, NT)等级[20],属于国家二级重点保护野生动物[21]。这2种鹿科动物在分布海拔上较为接近,均为植食性,生态位相近[22],很可能成为潜在竞争关系,因此可以作为研究同域分布有蹄类物种生态位分化的理想选择。在以往的研究中,欧善华等从食性角度比较了毛冠鹿和黑麂(Muntiacus crinifrons)的差异[23];孙佳欣等简单描述了同域分布水鹿和毛冠鹿在内的9种有蹄类动物日活动节律模式,但缺乏具体的(如季节间)差异性比较[24];邹启先等分析了麻阳河同域分布毛冠鹿和小麂(Muntiacus reevesi)的时空分布格局[6]。然而,关于同域分布水鹿和毛冠鹿针对性的活动节律比较研究尚未见报道,这2种鹿科动物在活动节律上究竟存在何种分化以实现共存?
为了回答以上问题,本研究于2017年5月—2018年4月,在四川贡嘎山国家级自然保护区内布设160个红外相机监测位点,利用红外相机数据来探究这2种鹿科动物在活动节律上的分化程度,以期为同域分布下近缘物种的共存机制研究和针对性保护管理策略的制定提供重要的数据参考。
1 研究方法
1.1 研究区概况
四川贡嘎山国家级自然保护区位于青藏高原东南缘(101°29'—102°12'E, 29°1'— 30°5'N),涉及甘孜藏族自治州的康定市、泸定县、九龙县和雅安市的石棉县,保护区面积为4 091.4 km2。保护区以保护高山生物多样性及多元生态系统为主,主要保护对象为大雪山系以贡嘎山为主的森林生态系统、各类珍稀野生动植物资源、低海拔现代冰川。保护区内山体高大,谷岭高差悬殊,主峰海拔7 556 m,是青藏高原东部的最高峰和东亚地区的第一高峰。区内年降水量800~900 mm,多集中在7、8、9三月。该地区属亚热带季风气候区,但因区内高耸于对流层中的巨大山岭对气流的阻挡,影响并改变了环流形势,形成了较为独特的气候特点,大致可以将全年分为旱季和雨季,其中旱季为11月至翌年5月,雨季为每年6—10月[25-26]。
1.2 相机布设与数据收集
2017年5月—2018年4月,结合当地以往监测、野生动物的活动痕迹、人力可到达性、红外相机在野外环境的布设效果[27]以及红外相机自身的安全性等因素,重点在保护区内的老榆林村、贡嘎山村周边区域选择合适的位置安放红外相机(Ltl 6 210)。研究期间共计160个红外相机监测位点,覆盖海拔区间为2 672~4 620 m(见图1)。所有红外相机监测位点的选择尽量兼顾不同生境和海拔,同时保证相邻红外相机位点间距至少300 m,安装高度控制在0.5~1.3 m,统一不使用引诱剂。相机安装时需对拍摄区域正前方的杂草等进行适当清理以保证拍摄聚焦区域不被遮挡,最佳聚焦拍摄点距离相机镜头约4~6 m。相机统一设置为混合模式(3张照片+1段视频),视频长度设置为10 s,拍摄时间间隔为2 s。详细记录每个相机位点的编号、生境、坐标等信息。每3~5个月收取一次数据,同时检查相机性能和更换储存卡。

图1 研究区位置及红外相机布设位点Fig. 1 Location of the study area and distribution of infrared camera traps during the study period
将研究期间回收的所有红外相机数据与点位信息进行系统归类整理,统一利用四川自然保护红外相机数据管理信息化平台(http://www.datawild.cn:9090/Bioplatform/, CDMS)[28]完成监测数据的录入、影像数据的物种识别与相关信息挖掘,最后从平台上导出所有数据Excel汇总表用于后期分析。
1.3 数据分析
本研究中关于活动节律的所有数据分析均以独立有效记录为基础,即同一相机位点下同一物种连续30 min(±1min)的拍摄记录(包括照片或视频),同时规定每次独立有效记录与红外相机拍到的动物个体数量无关[29]。
采用核密度估计方法(Kernel density estimation)[30]来分析水鹿和毛冠鹿的日活动节律,该方法假设目标物种的行为活动呈24 h周期性连续分布,其行为事件的有效记录为连续分布中获得的随机样本,该方法从数据样本本身出发分析数据分布特征,而在数据分析中不对数据分布外加任何假定。核密度曲线图使用R软件中的overlap包[31]和activity包[32]进行绘制。其中,横轴代表时间,纵轴(密度)代表该时间点上目标物种被探测到的概率[11,30,33]。采用条件密度等值线法来确定物种的活动高峰,通常以50%的内核阈值来计算活动比例集中的周期并以此来表征活动高峰期,条件密度等值线图利用R中的circular包“modal. region”函数来实现[34]。采用重叠指数(Coefficient of overlap)[35-36]来分析比较同一季节不同物种以及单一物种不同季节的日活动节律重叠程度,利用activity包“overlapEst”函数来绘制日活动节律的重叠图,并利用该函数的分布曲线重叠面积比Δ(当Δ=0时,表示完全分离;当Δ=1时,表示完全重叠)来表示具体的重叠程度。利用activity包中的“compareCkern”函数的Wald test对水鹿和毛冠鹿日活动节律进行概率检验,循环检测设定为1 000次[32],以此来分析日活动节律的差异性。重叠指数根据成对物种较小的样本数选取,当样本量小于50时采用Dhat 1值,当样本量大于75时采用Dhat 4值[33,35]。在对比分析水鹿和毛冠鹿日活动节律的重叠程度时,剔除仅记录到水鹿或者毛冠鹿活动(相机位点)的数据,保留拍摄到两个物种活动(共计105个相机位点)的数据[11,13]。所有检验的差异显著水平设定为P<0.05,统计分析和作图均在R 4.1.0中完成[37]。
2 结果与分析
研究期间共在110个相机位点拍摄到水鹿活动,在138个相机位点拍摄到毛冠鹿活动,共计获得两种野生动物的独立有效记录4 827次,其中水鹿1817次,毛冠鹿3 010次。
2.1 水鹿和毛冠鹿的日活动节律
整体来看,水鹿和毛冠鹿的日活动节律存在极显著差异(Δ=0.80,P<0.01),表明二者的日活动节律模式相似度较低(见图2)。其中,水鹿的日活动节律出现了3个活动高峰,分别在03:30—04:00、06:00—08:00、18:00—23:30,3个活动峰值大小和持续时间呈递增趋势。然而,08:00—18:00很少有水鹿的活动迹象,尤其是12:00几乎不活动。根据贡嘎山地区的气象数据,当地2017年5月—2018年4月的日出时间为06:10—08:09,日落时间为18:11—20:17,分析当地水鹿偏向于晨昏活动习性(见图2 A)。毛冠鹿的日活动节律呈现出明显的双峰趋势,较低的活动高峰出现在07:00—12:00,较高的活动高峰出现在17:00—21:30,两次活动高峰的持续时间均较长(接近5 h),21:30—07:00仅有少许活动迹象,表明当地毛冠鹿偏向于昼行性,且具有一定的晨昏活动现象(见图2 B)。

图2 水鹿(A)和毛冠鹿(B)的日活动节律分析图Fig. 2 Daily activity rhythms of Cervus equinus (A) and Elaphodus cephalophus (B)
2.2 日活动节律的季节间差异
水鹿在旱季(994次)与雨季(823次)间的日活动节律存在极显著差异(Δ=0.93,P=0.03)。其中,雨季03:50—04:30比旱季多1个小的活动高峰,雨季早晨(07:00)活动峰值出现时间比旱季(07:40)要早,而夜间活动峰值出现时间基本一致(20:30)。旱季早晨(06:30—09:00)和傍晚(17:30—23:30)的活动高峰期比雨季(06:00—08:00, 18:00—23:10)长至少半小时。旱雨季间活动最高峰值相近(见图3)。
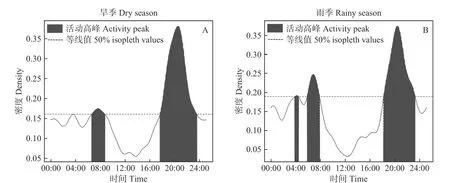
图3 水鹿旱季和雨季的日活动节律比较Fig. 3 Comparison of the daily activity rhythms of Cervus equinus in the dry season (A) and rainy season (B)
毛冠鹿在旱季(1 304次)与雨季(1 706次)间的日活动节律存在极显著差异(Δ=0.88,P<0.01)。旱季早晨(09:00)活动峰值出现时间比雨季(08:30)要晚半个小时,但傍晚(19:00)活动峰值出现时间比雨季(20:00)要提前1h。旱季早晨(07:30—11:00)的活动高峰期要比雨季(07:00—13:00)短2.5h,但傍晚(16:30—21:30)的活动高峰期要比雨季(18:00—21:30)长1.5h。雨季的活动最高峰值要低于旱季(见图4)。
在旱季,水鹿和毛冠鹿均在傍晚的活动高峰值更大、活动高峰期更长;而在雨季,二者的活动高峰值和活动高峰期呈现出相反的趋势(见图3;图4)。

图4 毛冠鹿旱季和雨季的日活动节律比较Fig. 4 Comparison of the daily activity rhythms of Elaphodus cephalophus in the dry season (A) and rainy season (B)
2.3 水鹿和毛冠鹿的日活动节律重叠
研究期间,共计有105个相机位点均拍摄到水鹿和毛冠鹿,占相机布设位点总数的65.63%。其中,有5个相机位点仅拍摄到水鹿,占比3.13%;有33个相机位点仅拍摄到毛冠鹿,占比20.63%。
对105个相同相机位点下水鹿和毛冠鹿的日活动节律进行Wald test分析,发现存在极显著差异(Δ=0.80,P<0.01),说明二者的日活动节律模式相似度较低(见图5)。整体上看,水鹿和毛冠鹿活动重叠时间主要集中在早晨和傍晚,而07:00-17:00期间分离程度较大(见图5)。二者在旱季和雨季的日活动节律同样表现出显著差异性(旱季Δ=0.83,P<0.01, 雨季Δ=0.74,P<0.01),但旱季的重叠指数以及最大活动高峰值均高于雨季(见图6)。

图5 水鹿和毛冠鹿日活动节律的重叠Fig. 5 Overlapping of the daily activity rhythms of Cervus equinus and Elaphodus cephalophus
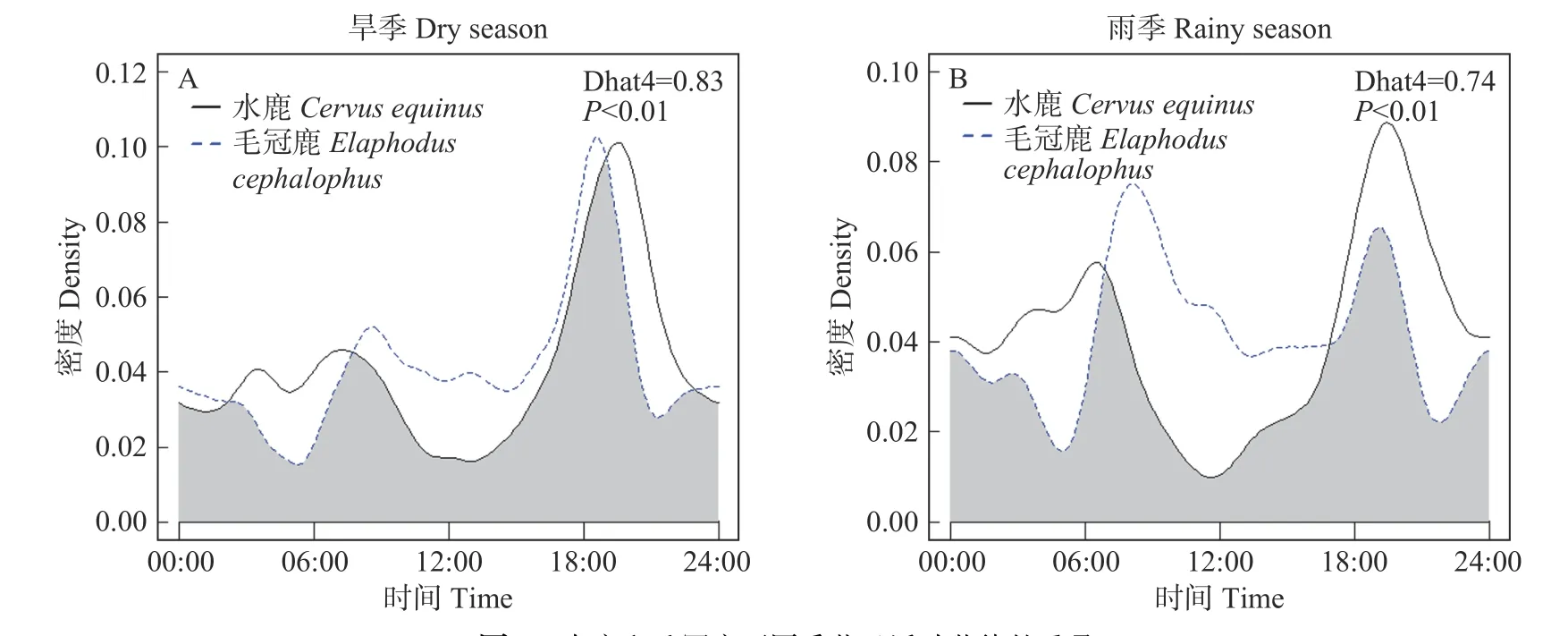
图6 水鹿和毛冠鹿不同季节日活动节律的重叠Fig. 6 Overlapping of the daily activity rhythms of Cervus equinus and Elaphodus cephalophus in the dry season (A) and rainy season (B)
3 讨论
3.1 关于日活动节律
活动节律是动物在长期的生存进化过程中应对环境变化而采取的一种生存适应[4,11,13]。通常认为,活动节律会受到动物本身生物钟、食物资源、能量代谢、性别差异、社群结构、种间竞争、天气情况等多方因素影响[38],且在长期的生存进化中,每种动物都会形成各自最佳的活动节律以适应当地特定的生存环境[39]。
本研究中,水鹿的活动节律出现了3个活动高峰,分别在03:30—04:00、06:00—08:00、18:00—23:30,但整体偏向于晨昏活动(见图2 A)。晨昏活动节律的研究结果与以往的部分研究类似[40-41],然而与孙佳欣等[24]综合评估四川王朗、四姑娘山、贡嘎山及亚丁4个自然保护区中水鹿活动曲线呈U型且属于特殊的单峰模式的研究结果差异较大,这可能与研究对象所在的区域空间尺度差异较大有关。另外,不同地区研究结果中水鹿的活动高峰有差异,如孙佳欣等[24]综合评估四川王朗、四姑娘山、贡嘎山及亚丁4个自然保护区水鹿的活动高峰出现在22: 00—24: 00;王盼[41]研究发现四川卧龙自然保护区水鹿的活动高峰出现在17:00且在22:00出现全日活动最低峰;何兴成等[40]研究发现四川鞍子河保护区水鹿日活动节律在不同季节具有较大差异:春季日活动高峰出现在17: 00—20: 00,夏季无明显日活动高峰,秋季日活动高峰出现在17: 00—19: 00,冬季日活动高峰分别出现在08: 00—10: 00、17: 00—19: 00、23: 00—02: 00。这表明同一物种的活动节律会存在不同程度的地域性差异,这种差异除了可能受不同地区气候条件、食物资源、种间竞争等多方因素影响外[38],还可能与不同研究中关于活动高峰的判断方式有关。如孙佳欣等[24]以活动峰值左右的一定时间段作为高峰,王盼[41]直接以活动峰值出现时间作为高峰,而本研究以50%的内核阈值来计算活动比例集中的周期并以此来表征活动高峰,目前已有不少研究证实本研究采用的方法可能更可靠[6,9-11,13-14]。毛冠鹿的日活动节律呈现出明显的双峰型,活动高峰出现在07:00—12:00与17:00—21:30,属于昼行性且具有一定的晨昏活动现象(见图2 B),这与以往其他地区关于毛冠鹿活动节律的研究结果类似[6,24,42]。
整体上看,水鹿和毛冠鹿的日活动节律相似度较低(Δ=0.80, P<0.01),尤其是活动峰值出现时间以及活动高峰期持续时间差异较大,如水鹿早上的活动高峰明显比毛冠鹿提前,而晚高峰结束时间比毛冠鹿晚2h。这可能是二者在同域分布情况下为了减少资源竞争实现长久共存而做出的适应性活动节律调整。刘鹏等[11]在对比贺兰山同域分布阿拉善马鹿(Cervus canadensis)和岩羊(Pseudois nayaur)的活动规律时认为,岩羊活动节律的调整很可能受种群数量占优势的马鹿种间竞争影响而做出的主动回避性适应。本研究中,从独立有效记录数(水鹿1817次,毛冠鹿3 010次)推测,毛冠鹿的相对种群数量明显较多,因此,我们同样认为种群数量的差异可能会迫使同域分布下种群数量较少的水鹿更早做出时间分化调整来减少种间资源竞争。
3.2 关于活动节律的季节间差异
本研究中,水鹿和毛冠鹿在旱季和雨季之间的日活动节律差异显著,表明二者应对季节变化而做出的日活动节律调整较大,这可能与季节变化引起的气候条件、食物资源等差异有关。已有部分研究指出,气候条件与食物资源可能是影响动物活动节律及季节性变化的主要驱动因子,而繁殖需求、捕食风险、干扰因子等其他因素同样会影响动物的活动节律[43-44]。贡嘎山保护区旱雨季间气温和食物资源差异较大,旱季(11月至翌年5月)气温整体较低,食物资源相对短缺;而雨季气温相对较高,许多植物会长出新叶和新芽,食物资源变得相对丰富。受旱季较低气温条件的影响,可能迫使动物避开低温时间段活动,这可能是水鹿和毛冠鹿早上的活动高峰较雨季延迟大约半小时以及毛冠鹿傍晚(19:00)活动峰值比雨季(20:00)提前1 h的重要原因。旱季里气温低,许多植被处于枯黄期,食物资源少且质量较差,加之地面积雪影响而不利于动物的取食活动,为了获得足够的食物量以满足自身能量需求(包括自身保暖以及其他活动等),动物不得不在可能暖和的时间开展取食活动,尤其是傍晚时间的活动,必须采集足够的食物才能有效满足较长夜晚时间的能量消耗,这可能是旱季水鹿和毛冠鹿均在傍晚的活动高峰值更大、活动高峰期更长的重要原因。相关研究发现水鹿和毛冠鹿均在旱季发情,雨季产仔(水鹿:冬季发情,8月左右产仔;毛冠鹿:9—12月发情,4—7月产仔)[22]。旱季里,动物需要适当增加活动强度来满足繁殖需求(包括寻找食物和配偶等),这可能是水鹿和毛冠鹿在旱季的活动高峰期均有适当延长的重要原因。雨季里,二者均会产仔、育幼,需要尽量保持活动隐秘以保护自身和幼崽的安全,气温明显暖和,食物资源丰富且质量高,食物资源的获取变得相对容易,毛冠鹿在较低活动强度下依旧能够满足自身能力需求;而水鹿种群数量相对较少,为了尽量减少种群间的资源竞争可能不得不采取适当的行为调整,这可能是水鹿在旱雨季间活动最高峰值相近、在03:50—04:30比旱季多1个小的活动高峰以及其活动高峰值和活动高峰期与毛冠鹿呈现相反趋势的重要原因。
3.3 关于日活动节律的重叠
同域分布物种的生态位格局可以看着是一种特殊的相互补偿机制,而物种之间通常会在食物、空间和时间尺度上表现出对资源利用的差异以实现共存[45-46]。
本研究中,有105个相机位点均拍摄到水鹿和毛冠鹿,相同位点数分别占所有水鹿发现位点数和所有毛冠鹿发现位点数的96.87%与79.37%,说明二者的活动空间区域存在较大重叠。然而,日活动节律分析发现二者在全年和不同季节下均存在显著差异,日活动节律模式相似度较低(见图5),说明二者在时间生态位上的分化程度较高,一定程度上减少了彼此间的竞争。这可能主要受当地气候条件、食物资源等方面的季节性差异影响,而旱季二者的繁殖需求同样会影响其日活动节律的调整。因此,二者在旱季的日活动节律重叠指数和活动强度略高于雨季。
生物对不同资源的选择利用是相互影响的,并非单一因子决定,因此仅凭二者在日活动节律模式上的分化尚不足以全面科学地评估二者在时间生态位上的分化与竞争[4,45-46]。已有研究发现,同域分布物种在资源选择上的分化首先体现在食物与生境的利用上且对物种间生态位分化产生重要影响[47]。如骆颖等[48]对比了贺兰山岩羊与马鹿的食性和生境选择,发现二者存在明显的分离,表明二者很大程度上可以同时对食物资源和生境选择利用,二者间不会出现干扰和竞争。本研究中,即便在水鹿和毛冠鹿均出现的相机点位下均未发现二者同时出现的画面。这与以往的部分研究结果类似,如胡磊[43]研究发现在内蒙古赛罕乌拉国家级自然保护区内同域分布的马鹿和狍(Capreolus pygargus)不会同时出现在某个相机位点;刘鹏等[11]研究发现贺兰山同域分布马鹿和岩羊在不同的相机位点上二者均未同时出现。因此,这一现象可能表明水鹿和毛冠鹿为了减少资源上的竞争而采取了主动回避彼此,在微生境的利用上,其时间生态位发生了完全分离。
综上所述,贡嘎山保护区水鹿和毛冠鹿日活动节律模式相似度较低,二者在时间生态位上分化程度较高且在微生境中产生了完全分离。加之水鹿对日活动节律模式的调整,扩大与毛冠鹿在时间生态位上的分化,表明同域分布水鹿和毛冠鹿可以通过时间生态位上的分化来减少二者在食物、空间与时间资源上的竞争以实现共存。
