没食子酸的生物学特性及其在畜禽生产中的应用进展
2022-04-29陈志龙黄运茂王一冰蒋守群
■陈志龙 黄运茂 王一冰 蒋守群*
(1.仲恺农业工程学院,广东 广州 510225;2.广东省农业科学院动物科学研究所,畜禽 育种国家重点实验室,农业农村部华南动物营养与饲料重点实验室,广东 省畜禽育种与营养研究重点实验室,广东 广州 510640)
近年来,我国畜禽养殖集约化水平日益提高,随着饲料全面禁抗时代的到来,畜禽肠道健康和免疫机能改善愈发成为养殖行业关注的热点。实际生产中,饲粮抗生素禁用导致畜禽疫病发生率提高、生长性能降低,临床治疗用药物用量增加,严重影响养殖效益和产品安全品质。因此,安全、高效、新型、替代抗生素饲料添加剂产品的研发迫在眉睫。
从植物中提取的酚酸,因为其特有的共轭结构和强大的给电子能力,具有较强抗氧化和抗菌等作用,其中部分还具有改善畜禽生长性能的作用[1]。没食子酸(Gallic acid,GA)是酚酸的一种,广泛存在于五倍子、锁阳、地榆、刺三加等草药中和山葡萄、石榴等水果中[2-10]。目前关于没食子酸抗菌、抗炎、抗氧化及抗癌特性在人和小鼠等模式动物上的研究较深入,但在畜禽生产上的研究和应用报道较少。
文章介绍了没食子酸的理化特性,对其发挥生物学活性的分子机理进行了讨论分析,并针对其在畜禽生产上的应用进行总结与展望,旨在为没食子酸对畜禽健康调控的进一步研究和生产上的科学应用提供参考。
1 没食子酸的理化特性和吸收利用
没食子酸又名五倍子酸、棓酸,学名为3,4,5-三羟基苯甲酸(C7H6O5),分子质量为170.12 u,是一种白色或淡黄色针状结晶或粉末,通常以一水化合物(C7H6O5·H2O)的形式存在。没食子酸含有3个酚羟基和1个羧基,其分子结构如图1所示[1]。
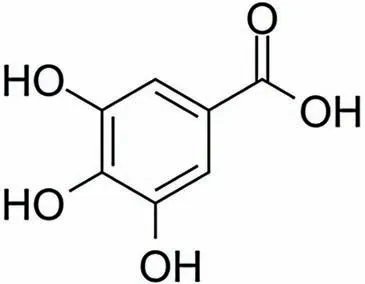
图1 没食子酸的分子结构式
没食子酸在自然界中来源广泛,五倍子、锁阳、地榆、刺三加等草药和芒果、石榴等水果中均含有没食子酸。没食子酸在我国已经有几十年的生产历史,可以从五倍子等植物中直接提取,从五倍子和石榴皮等浸出液中分离的棓单宁在酸、碱、酶等催化作用下也可生成没食子酸[9]。表1展示了近年来各种植物提取物中没食子酸的含量,其中,马桑皮提取物中没食子酸含量最高,可达到10.79%左右[3],五倍子提取物中棓单宁的含量高达18.3%,也可以作为没食子酸的主要来源[6]。

表1 各种植物提取物中没食子酸的含量
没食子酸衍生物包括两种类型:酯衍生物和儿茶素衍生物。没食子酸最常见的酯衍生物是烷基酯,主要由没食子甲酯(MG)、丙酯(PG)、辛酯(OG)、十二烷酯(DG)、十四甘酯(TG)和十六酯(HG)组成;主要的儿茶素衍生物是表儿茶素(EC)、表儿茶素没食子酸酯(ECG)、表没食子儿茶素(EGC)、加酸酯(GCG)和表没食子儿茶素没食子酸酯(EGCG)[11]。其中,EGCG 已被观察到具有强大的抗癌活性和对心血管和代谢性疾病的保护作用,具有多种机制。由于强效抗氧化剂清除活性氧的特性,没食子酸的衍生物如DG、PG、OG、TG 和HG 等,被广泛应用于食品制造和制药行业[11]。
没食子酸直接以中药或者提取物形式口服时溶解度低,吸收效果差,首过效应强,其在不同原料中的吸收效果也不同,所以在人中往往通过制成纳米制剂药片来提高其吸收率和生物利用度[12]。用结肠癌细胞Caco-2 单层膜进行体外试验,模拟纯没食子酸在细胞屏障上的跨膜转运,发现其质子梯度下的表观通透性系数为0.20×10-6cm/s[11]。没食子酸的代谢部位在肾脏,在肾脏经过甲基化、硫基化或者糖苷酸化后,原型和代谢产物于全身分布,最后经胆汁和尿液排出[11]。
2 没食子酸的生物学作用与机制
作为一种有机酚酸,没食子酸因为其较强的还原性,易与蛋白质、酶以及细胞膜上受体发生反应,激活或抑制一系列分子信号,发挥抑菌、抗氧化、抗炎及促进物质转运等生物学作用,前人对其功能的研究见表2[1]。
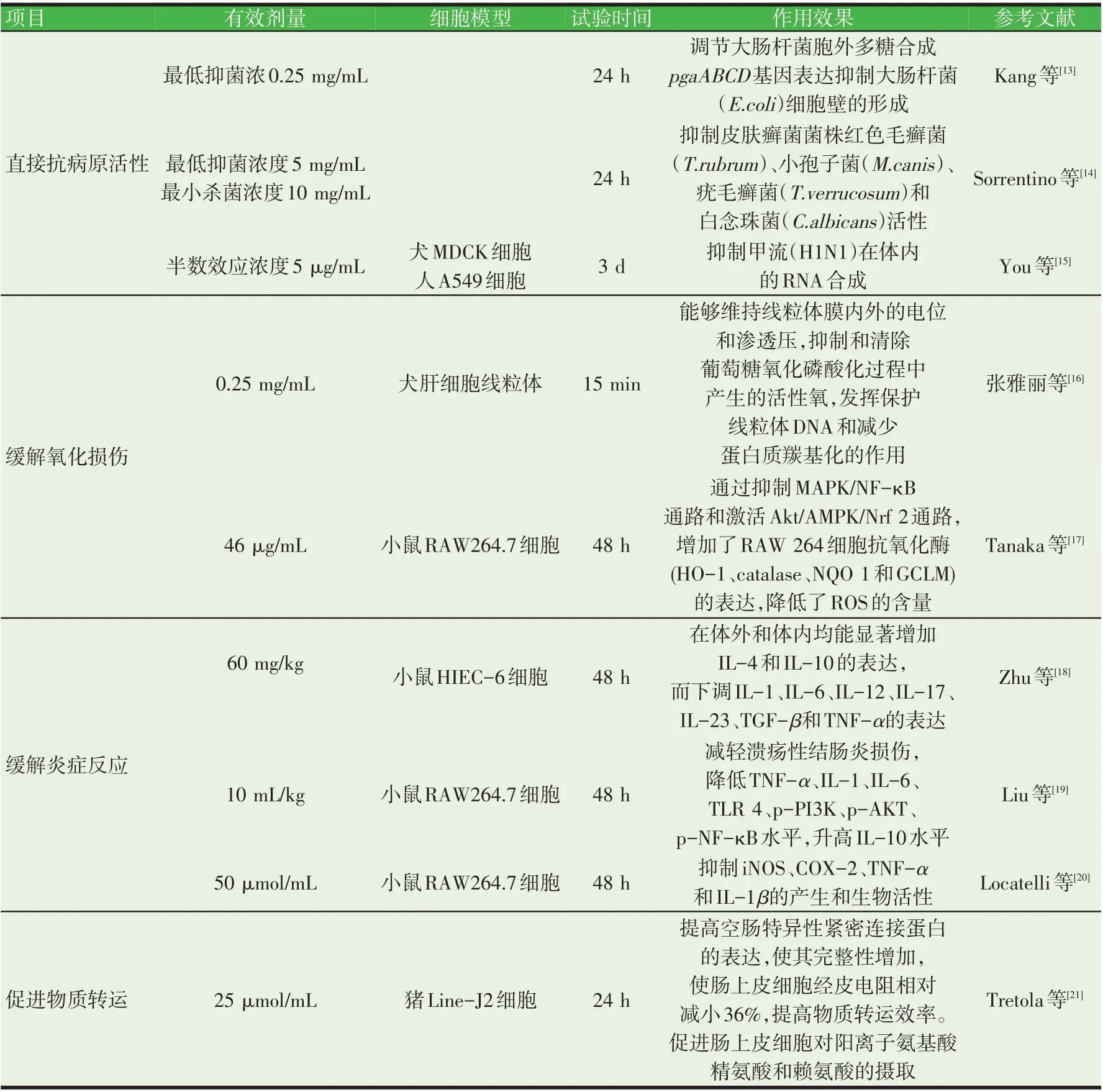
表2 没食子酸的生物学功能
2.1 杀灭/抑制病原菌的作用
没食子酸在体外有广泛的抗菌活性。体外研究表明,没食子酸通过调节大肠杆菌胞外多糖合成pgaABCD基因表达抑制大肠杆菌(E.coli)细胞壁的形成[14];没食子酸通过调节mdoH基因和OpgH蛋白的表达,有效抑制福氏志贺菌(Sh.flexneri)细胞膜的形成和活性[22],并通过调节ICA操作子的表达,对金黄色葡萄球菌(S.aureus)有特异性的抗生物膜作用[23]。体内研究证明,没食子酸可抑制幽门螺杆菌生长(H.pylori)[24]。
没食子酸不仅可以抑制、杀灭细菌,而且也具有很强的抗真菌活性[14]。对新鲜黑块菌进行贮藏试验,观察到没食子酸的体外抗菌活性,假单胞菌属、肠杆菌科和伞状菌的存活率显著下降,并且没食子酸对所有被测的皮肤癣菌菌株红色毛癣菌(T.rubrum)、小孢子菌(M.canis)、疣毛癣菌(T.verrucosum)和白念珠菌(C.albicans)均显示抗真菌活性[25]。
You 等[15]研究表明,甲型H1N1 流感病毒对没食子酸敏感,其在人体内感染的过程会被没食子酸抑制。没食子酸也具有抗乙肝病毒活性[26]。GutiéRrez等[27]以没食子酸强大的抗菌活性为理论基础进行改良,合成了甲氧苄啶等抗菌剂,用于治疗某些微生物传染病。
2.2 没食子酸的抗氧化作用机制
核因子E2相关因子2(Nrf2)是一种转录因子,负责调节哺乳动物细胞的氧化还原平衡和抗氧化及Ⅱ期解毒反应[28]。应激条件下,Keap 1中的——SH基团的修饰或Nrf2 的磷酸化促进了Nrf2 与Keap 1 的解离以及Nrf2 向核的移位。在与Maf 肌肉腱膜纤维肉瘤癌基因同源物重组蛋白(MAF)结合后,Nrf2激活ARE(抗氧化反应元件,antioxidant response element),增加Nrf2 调控抗氧化酶基因[例如血红素氧化酶(oxygen⁃ase-1,HO-1)、谷胱甘肽巯基转移酶(Glutathione Stransferase,GST)、磷酸酰胺腺嘌呤二核苷酸醌氧化还原酶-1(NADPH quinineoxidoreductase-1,NQO-1)]的转录[29]。Zhang 等[30]用没食子酸预处理人肝脏癌细胞HepG 后使用H2O2建立氧化损伤模型,发现没食子酸处理组的Nrf2和HO-1表达水平显著上升,从而提高了下游抗氧化酶谷氨酸半胱氨酸连接酶(glutamatecysteine ligase modifier,GCLM)和过氧化氢酶(cata⁃lase,CAT)的表达。当抑制HepG2细胞的Nrf2基因转录时,没食子酸引起的HO-1和NQO-1表达上调被抑制。Tanaka等[17]研究发现,通过敲除Nrf2可显著降低没食子酸诱导的抗氧化酶NQO-1、HO-1、CAT 和GCLM 的表达,提示没食子酸对Nrf2靶向抗氧化基因的上调作用,从而提高抗氧化酶的活性。Pandurangan等[31]研究表明,没食子酸可导致Nrf2 下游靶点的NQO-1 和二磷酸尿苷葡糖醛酸转移酶(uridine di⁃phosphate-glucuronosyltransferases,UDP-GT)的表达增加,从而保护结肠黏膜,缓解结肠炎,降低结肠癌的发病风险。没食子酸可能是通过激活Keap1-Nrf2 通路,但在畜禽上的具体作用机制未见报道。
2.3 没食子酸缓解炎症作用机制
核转录因κB信号(Nuclear factor kappa beta,NFκB)通路在免疫调节、炎症、应激反应和细胞凋亡等方面发挥着重要作用。在病原刺激过程中,白细胞介素-1β(interleukin,IL-1β)和肿瘤坏死因子-α(tumor necrosis factor,TNF-α)等炎症因子刺激髓核细胞激活I-κB激酶,使I-κB蛋白在泛素化和磷酸化后降解,并暴露于NF-κB 二聚体,特别是二聚体p65-p50。然后,p65 被磷酸化,p65-p50 被导入细胞核,二聚体p65-p50 在进入细胞核后激活转录因子,同时在细胞核内被组蛋白乙酰转移酶(HATS)乙酰化。乙酰化的p65抑制了其与IκBα的结合能力,导致NF-κB信号通路的持续激活[32]。有研究发现,没食子酸可以抑制NF-κB途径上游的促炎因子TNF-α和IL-1β/6诱导的p65磷酸化,抑制p65-p50向细胞核的转移,阻抑NFκB 途径的激活,降低血栓形成素4、分解素和金属蛋白酶的分泌,防止腰椎间盘退变[33],抑制IL-4/10 释放,则可以降低结肠炎肺炎和肺气肿,酒精性肝炎和酒精性胃溃疡的发病风险[17]。Yang等[34]发现5 μg/mL没食子酸可通过抑制A549 肺癌细胞的p65 乙酰化抑制LPS诱导的NF-κB信号转导从而抑制诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)和环氧化酶-2(cyclooxygenase-2,COX-2)的表达。Liu等[19]使用5%右旋糖酐硫酸钠建立溃疡性结肠炎模型的大鼠经1、2 mL/kg和10 mL/kg没食子酸处理后,发现没食子酸处理减缓了右旋糖酐硫酸钠诱导的TLR依赖的磷酸化PI3K/AKT/NF-κB 的表达水平升高,从而减轻溃疡性结肠炎损伤,降低丙二醛(Malondialdehyde,MDA)、肿瘤坏死因子α、IL-1、IL-6、TLR-4(Toll 样受体4,Toll like reactor 4)、p-PI3K(磷酸化磷酸肌醇-3-激酶,phosphorylated phosphoinositide-3-kinases)、p-AKT(磷酸化蛋白激酶A,phosphorylated protein kinases A)、p-NF-κB 水平,升高SOD(超氧化物歧化酶,superoxide dismutase)、GSH-Px(谷胱甘肽过氧化氢酶,glutathi⁃one peroxidase)、CAT 和IL-10 水平,其中10 mL/kg 没食子酸处理组效果最佳。可见,没食子酸调节炎症反应的作用对维持肠道健康的策略具有重要意义。因此,进一步研究揭示其在特定动物及其不同生理阶段下抑制动物肠道炎症的机制十分必要。
3 没食子酸在畜禽养殖中的应用
目前关于没食子酸在畜禽生产上的研究报道较少,现有研究显示,没食子酸在改善动物生长性能和健康方面具有显著作用。
3.1 没食子酸改善畜禽生长性能和肉、蛋品质的作用
在饲料中添加没食子酸,能通过改善动物机体的代谢能力,提高生长性能和改善肌肉和蛋的品质。Samuel 等[35]在1 日龄爱拔益加肉鸡饲粮中添加75~100 mg/kg 纯度为90%的没食子酸提高了饲料转化率,当没食子酸的添加量为100 mg/kg和150 mg/kg时可提高42日龄肉鸡胸肌率。Starčević等[36]在罗斯鸡饲粮中添加5 g/kg没食子酸后发现,35日龄罗斯鸡血浆n-3多不饱和脂肪酸含量和胸肌脂肪沉积增加,同时降低肝脏胆固醇含量。Lee 等[37]研究发现,饲粮添加1%没食子酸和亚麻油(1:1)混合物的35 日龄罗斯鸡大腿中总酚含量、1,1-二苯基-2-吡啶酰肼(DPPH)自由基清除活性、2,2-偶氮-(3-乙基苯并噻唑啉-6-磺酸)(ABTS+)还原活性和还原力显著提高,降低贮藏第7 d 的胸肌挥发性盐基氮含量,同时大腿肌肉储存时细菌丰度增加慢于对照组,说明饲料添加没食子酸后的鸡肉更不易腐败而耐储存。Jung 等[38]在蛋鸡饲粮中添加1%没食子酸和亚麻油(1:1)的混合添加剂可以提高鸡蛋的抗氧化能力,降低蛋黄的MDA含量,提高蛋清的总抗氧化能力,提高蛋黄的脂肪酸含量,降低胆固醇水平。
3.2 没食子酸对畜禽机体抗氧化作用
饲料添加没食子酸可以提高动物机体抗氧化能力,减轻氧化损伤。Samuel等[35]在1日龄爱拔益加肉鸡饲粮中添加75~100 mg/kg纯度为90%的没食子酸时,发现其上调21日龄肉鸡血浆总抗氧化能力和总超氧化物歧化酶活性,降低了MDA含量。Al-sagheer 等[39]在新西兰白兔饲粮中添加500 mg/kg没食子酸,有效改善了热应激对白兔生长性能和血液生化特性的负面影响,显著增加了白细胞、淋巴细胞和异嗜细胞的数量,提高过氧化氢酶活性,降低谷胱甘肽过氧化物酶和丙二醛的含量,还显著提高了其对粗蛋白的消化率。以上研究表明,日粮中添加没食子酸能增加畜禽体内抗氧化酶活性,清除过量自由基类物质,减少机体氧化损伤。
3.3 没食子酸改善肠道健康的作用
肠道健康是动物正常生长的重要保证。除了抗氧化作用,没食子酸还能通过改善肠道微生物群落和缓解肠道炎症来改善肠道健康。Mašek 等[40]发现,饲粮添加5 g/kg 没食子酸能改善35 日龄罗斯鸡盲肠微生物发酵状态,使得盲肠内容物pH 降低,甲酸、乙酸和丁酸等短链脂肪酸含量增加,盲肠抗病能力增强。Lawal 等[41]研究则发现,饮水中添加1~3 ml/L 没食子酸,能有效改善黄曲霉毒素导致的肉鸡回肠黏膜结构的破坏和炎性浸润,从而缓解了黄曲霉素攻毒导致的肉鸡肠道消化吸收功能的损伤和生长性能的下降。
没食子酸可直接作用于肠道上皮,促进动物肠道对其他营养物质的吸收。在猪小肠上皮细胞IPEC-J2上的试验表明,没食子酸可能提高空肠特异性紧密连接蛋白的表达,使其完整性增加,50 μmol/L处理细胞时经皮电阻相对减小36%,有利于物质转运[41]。此外,没食子酸对肠上皮细胞对阳离子氨基酸、精氨酸和赖氨酸的摄取有积极影响,其机理可能为介导了阳离子氨基酸转运蛋白-1(CAT-1)的表达[21]。没食子酸对阴离子氨基酸的转运有积极影响,但具体的作用机制尚待进一步研究[11]。
综上所述,没食子酸可在饲料和饮水中添加提高动物生长性能和机体抗氧化能力,缓解炎症损伤和改善盲肠微生物群落来改善肠道健康,改善肉蛋品质,延长产品保存时间。欧盟食品安全局(EFSA)推荐饲料中添加没食子酸的安全剂量为不大于5 mg/kg[40],而实际生产中作为饲料添加剂没食子酸应用研究报道均显示有效剂量往往在此之上,且针对具体动物品种和日龄的剂量选择存在差异。
4 总结与展望
目前没食子酸在人和小鼠等模式动物上已经有了广泛的研究。没食子酸本身吸收效果较差,但是对促进其他营养物质的转运有积极效果,在促进负电氨基酸的转运机制还需进一步研究。
没食子酸有广泛的抗菌抗炎抗氧化作用,同时也具有促生长作用,但是作为饲料添加剂长期使用中,为了达到理想的效果可能会超过安全标准,对动物肠道和机体的不利影响并没有得到有效评估。未来需要在特定动物及其不同条件下研究没食子酸对动物肠道作用的具体机制,以便得出科学的添加量和添加剂型。
