猪血浆半胱氨酸蛋白酶抑制肽的筛选及其机制
2022-04-27王嘉敏王逐鹿常思佳刘汉雄王震宇
王嘉敏,王逐鹿,常思佳,刘汉雄,王震宇
(大连工业大学 食品学院,国家海洋食品工程技术研究中心,辽宁 大连 116034)
我国猪血资源非常丰富,作为我国一大禽畜类血液,我国对动物血液的使用率较低,基本都是被用来制作成一些血肠、血豆腐等食品,但这些传统食品的加工效率低、带来的综合效益也较低[1]。近年来对动物血液的研究才逐渐丰富起来,在食品加工和生物医药中的应用也逐渐增多,如制作一些新型血液制品等,提高了我国血液的开发利用程度[2-4]。猪血作为畜产品加工的副产物,因其血腥味较重、呈现的鲜红色或者深红色的色泽差、储藏期短,限制了对猪血的利用,诸多因素导致目前我国血液加工利用率水平低、产品附加值低,宝贵的血资源得不到充分的利用,更难以发挥其蛋白的价值。已有研究表明血浆能够抑制鱼糜凝胶劣化过程中组织蛋白酶活性、增强凝胶强度[5-7]。但是,血浆直接添加可能会导致产品颜色、风味的显著变化,影响鱼糜品质,因此开发基于血浆的组织蛋白酶抑制肽能够有效避免上述劣势,为猪血浆的综合利用及其制品在水产加工中的应用提供了理论基础。
1 材料
1.1 原料与试剂
新鲜猪血(已加入抗凝剂),购买于大连市仟和市场;胰蛋白酶与组织蛋白酶L 购买于江苏南京沃瑞达斯有限公司;其余化学试剂均为分析纯。
1.2 仪器
超高效液相四极杆飞行时间串联质谱(UPLCQ-TOF Impact II),德国Bruker 公司;分子对接软件Discovery Studio v2017,法国Dassault Systèmes 公司;AKTA avant-25 蛋白纯化仪,美国Cytiva 公司。
1.3 方法
1.3.1 猪血浆蛋白的制备
新鲜猪血浆在4 000 g、10 min 条件下进行离心,取上清液。加入硫酸铵至30%饱和度,4 ℃静置6 h后,8 000 g 离心20 min 取上清,在上清液中继续添加硫酸铵至60%饱和度,4 ℃静置6 h 后10 000 g 离心 20 min 取沉淀的蛋白,加入3 倍(v/w)的PB 缓冲液 (20 mmol·L-1,pH 6.8),在3 500 Da 透析袋中透析12 h,去除其中多余的硫酸铵,4 ℃保存用于后续酶解反应。
1.3.2 多肽的酶解制备
在透析过的蛋白溶液中,按200 ∶1(原料蛋白∶胰蛋白酶)(w/w)的比例加入胰蛋白酶,在45 ℃,pH 8.0 的条件下酶解3 h,每隔30 min 用0.5 mol·L-1的NaOH 溶液进行调节维持pH 恒定,采用pH-state法对酶解过程中的水解度进行测定[8],水解度的计算公式为
式中:h为酶解液中生成的游离氨基的数量,mmol·g-1protein;hhot为底物中蛋白质中肽键的总数,mmoL·g-1protein,在本实验中根据原料种类选取8.8 mmoL·g-1protein[9];B为滴定消耗NaOH 的体积,mL;Nb为用于滴定的NaOH 浓度,mol·L-1;a为氨基的解离度,其计算参考公式(2);Mp为底物中蛋白质的总含量,g。
式中:pH取酶解体系pH 值8.5;碱滴定的情况下pK为氨基解离常数,取值7.5。
1.3.3 多肽混合物的弱阴离子交换层析
酶解后的多肽混合物使用AKTA avant-25 仪器以及DEAE-Sepharose 柱进行分离纯化[10],用含有不同氯化钠浓度的流动相进行梯度洗脱,梯度设置为 0.1 mol·L-1、0.2 mol·L-1、0.3 mol·L-1、0.4 mol·L-1和 0.5 mol·L-1氯化钠浓度,每个浓度梯度冲洗一个柱体积,流速为1 mL·min-1,按峰收集,并测定抑制活性。
1.3.4 多肽混合物的超滤分级
取抑制活性最强的阴离子交换组分,分别采用 3 kDa 和10 kDa 的超滤膜进行分级,分别得到分子量为0 ~3 kDa、3 ~10 kDa 以及10 kDa 以上3 个分子量组分。分别进行冻干,超纯水复溶后测定抑制活性。
1.3.5 多肽组织蛋白酶抑制活性的测定
组织蛋白酶L 活性的测定按照Barrett 方法[11-12],并略微有改动。
(1)组织蛋白酶L 活力的测定。一个酶活力单位(U)定义为在反应温度37 ℃及条件下,1 min 内能够水解底物并释放出1 nmol AMC 所需的酶量(1 nmol AMC·min-1)。①溶液配制。反应终止液:0.1 mol·L-1乙酸-乙酸钠缓冲液,含0.1 mol·L-1氯乙酸(pH为6.0);反应缓冲液:0.34 mol·L-1乙酸钠、0.06 mol·L-1冰乙酸、4 mmol·L-1EDTA-2Na、8 mmol·L-1DTT;底物:Z-苯丙胺酰-精氨酸-7-氨基-4-甲基香豆素配制成40 μmol·L-1的溶液,作为组织蛋白酶L 的专一性底物。②组织蛋白酶L 活力的测定。组织蛋白酶L 与PB 缓冲液按照5 ∶2 充分混合,取混合液100 μL,加入底物Z-苯丙胺酰-精氨酸-7-氨基-4-甲基香豆素25 μL,加入反应缓冲液50 μL,充分混匀37 ℃水浴15 min,然后再加入200 μL 反应终止液,混匀后终止反应,用荧光分光光度计测定其荧光强度。测定所用的激发波长和发射波长分别为380 nm和460 nm。
(2)猪血浆对组织蛋白酶L 的抑制活性的测定。样品1:0 ~3 kDa;样品2:3 ~10 kDa,样品3:10 kDa 以上。测样溶液按照表1 进行配制,混匀后37 ℃水浴15 min。

表1 测样制备
(3)抑制率测定。在激发波长380 nm,发射波长460 nm 条件下用荧光分光光度计测定其荧光强度,设置对照组为A1,空白组为A2,样品为A3。抑制率测定公式为
1.3.6 超高效液相四极杆飞行时间串联质谱测定多肽氨基酸序列
通过用超高效液相四极杆飞行时间串联质谱联用方法对多肽溶液进行质谱分析鉴定,具体方法参考文献[13],在NCBI 网站上的数据库检索猪(S.scrofa)的基因库[14],进行氨基酸序列比对。
1.3.7 分子对接
将比对完的多肽氨基酸序列进行筛选,用分子对接软件Discovry Studio 2017 模拟抑制肽与组织蛋白酶L(PDB ID:3BC3)相互作用,并分析具体的相互作用结合的具体氨基酸,作用活性位点[15-16]。
2 结果与分析
2.1 猪血浆蛋白的水解过程
由图1 可知,酶解前期,在0 ~1.0 h,由于底物蛋白浓度较高,胰蛋白酶活旺盛,酶解体系pH基本不变,酶解程度较高,蛋白酶水解较快,水解度高幅度增长;随着酶解的持续进行,从进行酶解 1.0 h 后胰蛋白酶酶解速度开始降低,胰蛋白酶降解,酶解体系pH 变化大,酶解速度降低,水解度虽然一直有提升趋势但增长变得逐渐缓慢。胰蛋白酶酶解 3.0 h 的水解度为3.65%。
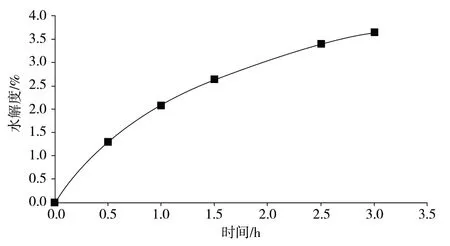
图1 酶解反应中蛋白水解度变化曲线
对血浆全蛋白、盐析得到的蛋白沉淀以及酶解后得到的多肽混合物进行SDS-PAGE凝胶电泳分析,结果如图2 所示,猪血浆蛋白分子量大小基本集中在50 kDa 以上,少部分为15 kDa 和25 kDa,由此可得出猪血浆中的蛋白大多都可被胰蛋白酶水解为肽;用硫酸铵盐溶液盐析后的血浆蛋白分子量大小集中于50 ~75 kDa,少部分为25 kDa 和15 kDa,比盐析之前的血浆蛋白量有所减少,说明盐析使得大部分蛋白沉淀下来;血浆蛋白酶解后,大蛋白几乎被降解,肽的分子量主要集中分布在20 kDa 以下,呈弥散条带,分布范围广,说明大部分蛋白基本都被酶解为肽,且酶解后的肽其中的成分复杂,由多种小分子肽组成。

图2 猪血浆蛋白及盐析和酶解后的SDS-PAGE 凝胶电泳分析
2.2 猪血浆半胱氨酸蛋白酶抑制肽的分离纯化
根据目标多肽的性质预测,选用阴离子交换层析作为第一步纯化。大量呈正电荷的多肽没有被填料吸附,在梯度洗脱程序前即流出。使用含有氯化钠的缓冲液,通过改变进入柱子的缓冲液的电导率,将不同结合力的蛋白冲洗下来并进行收集。根据出峰结果适当调整氯化钠浓度梯度,最终得到如图3所示的4 个分离完全、不拖尾的洗脱峰,按出峰位置分别收集洗脱液。

图3 猪血浆酶解物DEAE-弱阴离子交换层析分离纯化
从表2 中可以看出经过阴离子交换层析后,带有负电荷的多肽组分普遍具有更高的酶活抑制率,其中洗脱峰1 的抑制率最高,为26.83%。虽然其体现出了较显著的组织蛋白酶抑制活性,但抑制率仍较低,需要对其组分进行进一步分离纯化。

表2 猪血浆酶解物阴离子交换层析后组分的酶活抑制率
进一步采用超滤分级的方式对洗脱峰1 中的组分进行分离,并对回收的多肽组分进行SDS-PAGE电泳分析,如图4 所示。由图4 可知,10 kDa 以上超滤截留液中含有大量分子量15 kDa 左右以及超 35 kD 的蛋白,而0 ~3 kDa 超滤截留液中几乎不含有大分子蛋白条带,仅显示了最下端的多肽条带,3 ~ 10 kDa 超滤截留液中大蛋白含量明显减少,小分子蛋白和多肽的条带明显。在对上述3 个组分进行组织蛋白酶抑制率分析后(表3),可见小分子量组分普遍具有更高的酶抑制活性,其中洗脱峰1 中分子量0 ~3 kDa 的组分其酶活抑制率最高,达35.15%。该组分具有一定量的负电荷,同时分子量较小,能有效与组织蛋白酶相结合,进而达到竞争性抑制的效果,但具体多肽的氨基酸序列情况以及发挥功效的作用机制还有待探究。因此有必要对其中的多肽氨基酸序列进行测定分析。
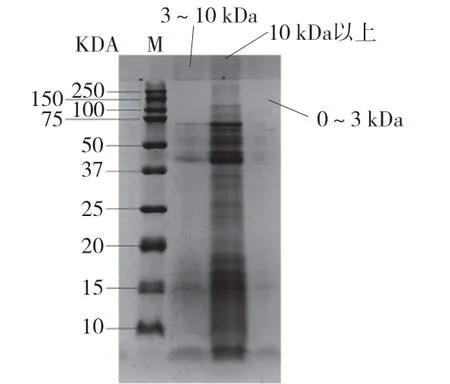
图4 猪血浆蛋白酶解超滤后肽的SDS-PAGE 凝胶电泳分析

表3 洗脱峰1 中不同分子量组分的酶活抑制率
2.3 组织蛋白酶抑制肽组分中的多肽氨基酸序列
将超滤分级后分子量为0 ~3 kDa 的肽溶液进行质谱分析。在Mascot 软件上对UPLC-Q-TOF 联用质谱后的质谱图进行分析,再利用NCBI 网站数据库中猪的基因库进行多肽氨基酸序列的比对,一共比对出35 条肽段,如表4 所示,长度范围为16 ~40 aa,分子量为2 013.9 ~4 306.7 Da,来源蛋白包括假定蛋白质、组蛋白乙酰转移酶、转录激活蛋白Pur-alpha、 N-甲基转移酶、细胞周期素依赖性激酶1、KH 结构域RNA 结合蛋白、琥珀酰辅酶A 合成酶、血红蛋白、肌联蛋白、纤溶酶原、未表征蛋白、包膜糖蛋白、假定转甲状腺素蛋白、唾液酸胎球蛋白、烯醇化酶1(α)、内源性因子钴胺素、F-box 蛋白、假定胰蛋白酶以及二酰甘油酰基转移酶1 等。根据目标蛋白来源、细胞定位进行筛选,例如琥珀酰辅酶A 合成酶、组蛋白乙酰转移酶转录激活蛋白Pur-alpha、N-甲基转移酶都是在酶解血浆蛋白时细胞所产生的作用酶,用以提供酶解过程的需要或者其他活动,不是猪血浆本身所含有物质酶解之后产生的,是细胞生理活动中所产生的。因此,此类多肽氨基酸序列不做进一步研究。根据上述超滤分级后酶活性测定等实验的分析得到的分子量为0 ~3 kDa 的多肽氨基酸序列抑制率最高,筛选出分子量为3 kDa 以下的多肽氨基酸序列。再如,某些蛋白在血浆中的含量较少的也不作进一步研究。因此本实验共筛选出分子量小于3 kDa 的两个肽段,如表4 所示,将序号24 和25 的两条多肽氨基酸序列命名为PP1、PP2,PP1 蛋白来源为血红蛋白,其氨基酸序列分别为EAVLGLWGKVNVDEVGGEALGR,长度为22 aa,分子量为2 267.2 Da,PP2 蛋白来源为纤溶酶原,其氨基酸序列为VILGAHEEYHLGEGVQEIDVSK,长度为22 aa,分子量为2 421.2 Da,然后进一步通过分子对接模拟探索半胱氨酸蛋白酶抑制剂的作用机制。
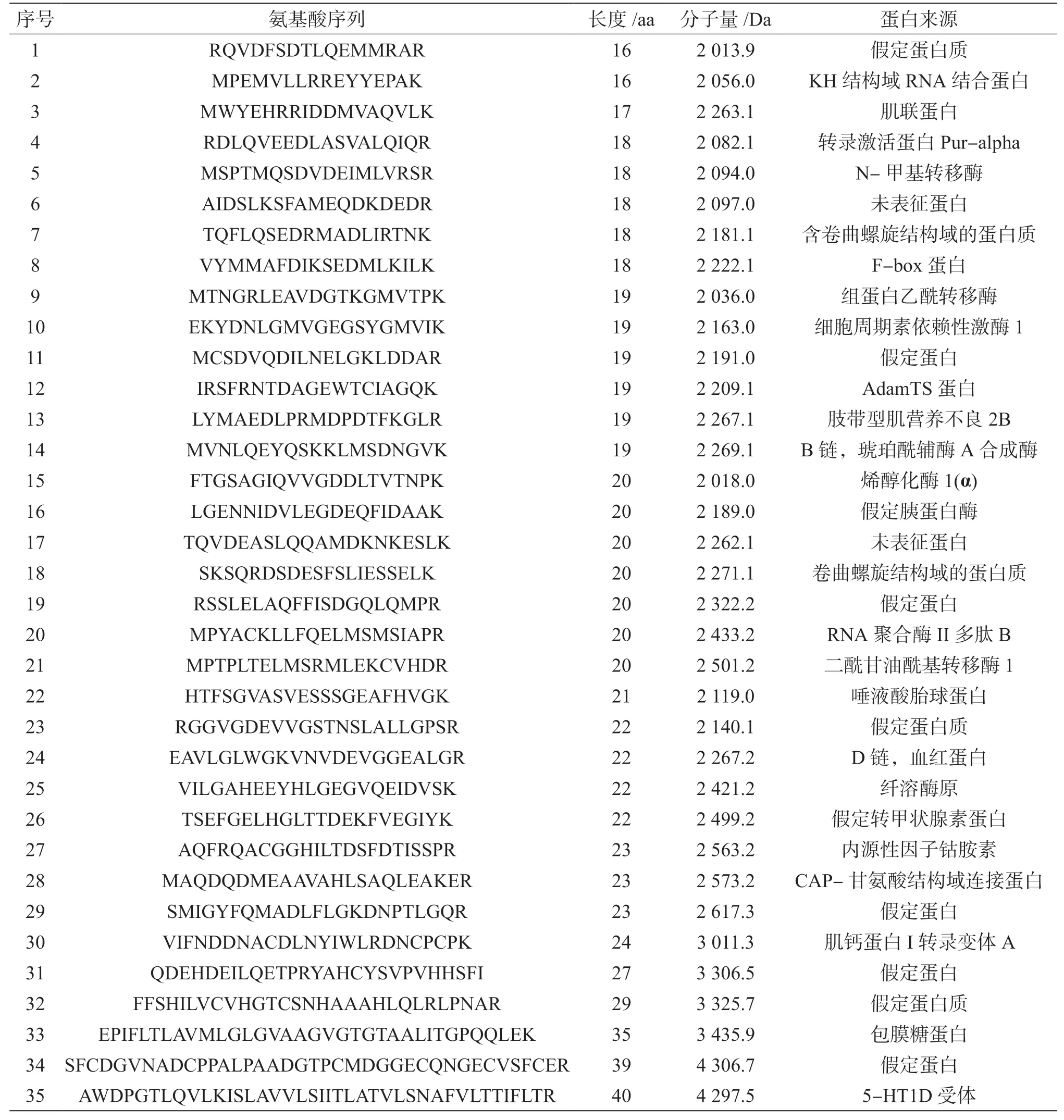
表4 猪血浆中0 ~3 kDa 肽溶液质谱分析后氨基酸序列检索结果
2.4 抑制肽PP1 和PP2 与组织蛋白酶L 相互作用的机制
如图5(a)所示,红色球部为组织蛋白酶L 的作用部位,图5(b)中球棍分子模型为PP1,条带分子模型为组织蛋白酶L,其钩状凹下的部位为作用活性部位催化中心,PP1在组织蛋白酶L 的活性位点作用,阻止了组织蛋白酶L 与其他蛋白等物质的结合作用,起到了抑制其作用活性的目的。图6 所展示的是分子模拟分析二者相互作用位点附近关键氨基酸。表5 可以看出抑制剂与组织蛋白酶L 之间的相互作用情况,其分子间作用力有静电力、氢键,其中氢键为二者发生相互作用的主要作用力,分子间作用力键长范围为1.961 04 ~5.018 80 Å。分子模拟结果表明组织蛋白酶L 作用的作用位点为H166、H234、H239、H244、H256、H313,其氨基酸位点为LYS117、SER158、GLU159,肽作用位点为H3、H4、H6、H14、N15,其氨基酸位点为GLY20、GLN21、GLY23,在二者的混合体系中,两者通过活性位点紧密贴合,并相互作用。

表5 PP1 与组织蛋白酶L 相互作用的关键氨基酸化学作用力类型及键长
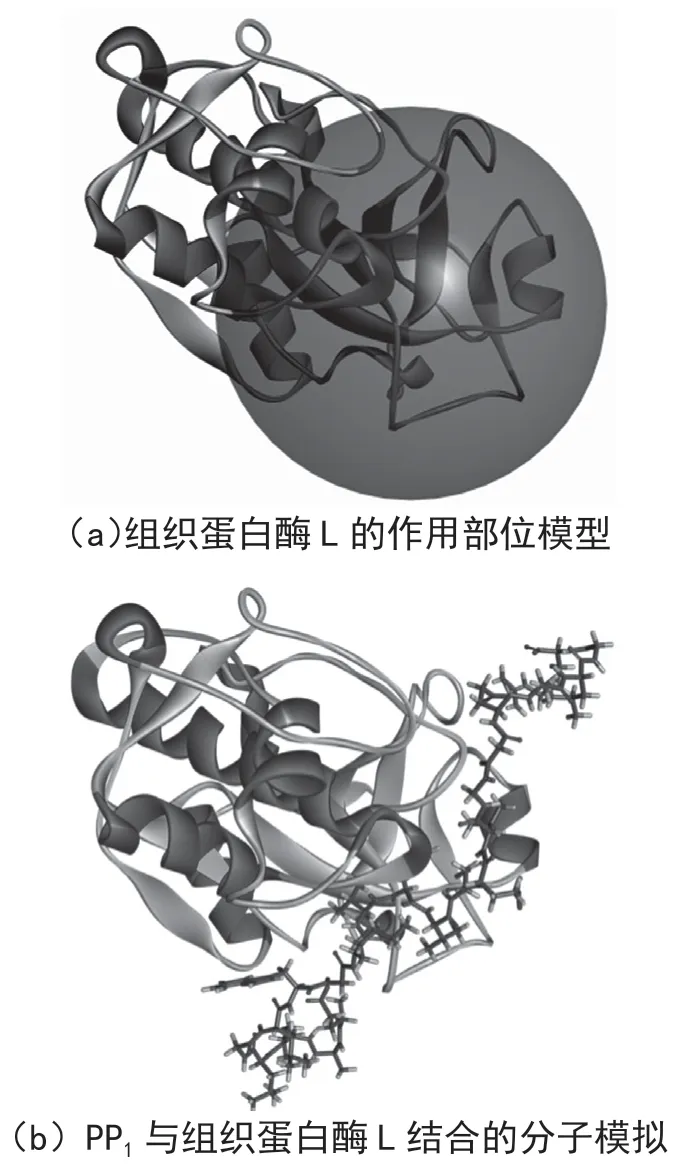
图5 组织蛋白酶L 的作用部位及其与PP1 相互作用的分子模拟
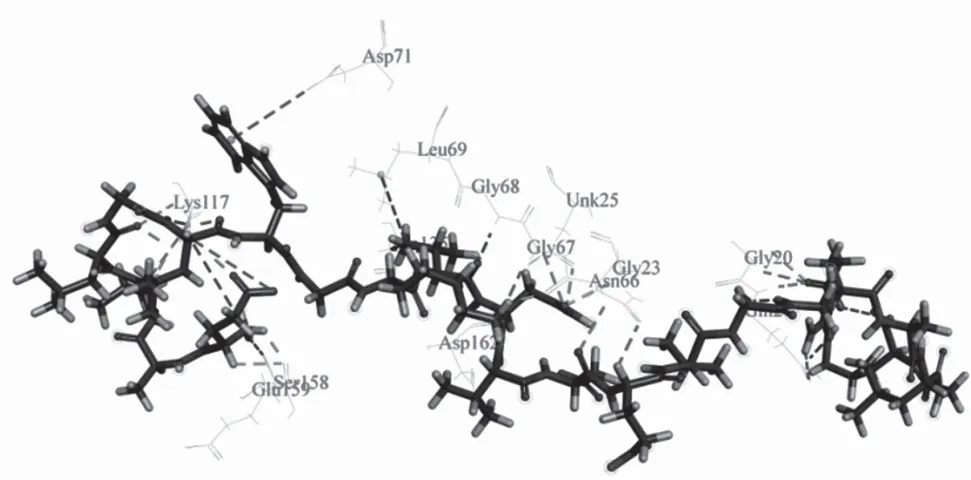
图6 PP1 与组织蛋白酶L 相互作用位点附近的关键氨基酸
如图7 所示,图中条带模型分子为组织蛋白酶L,球棍模型分子为PP2,以抑制剂纤溶酶原作为供体,组织蛋白酶L 为受体,钩状凹下的部位为作用活性部位催化中心,PP2 在组织蛋白酶L 的活性位点作用,阻止了组织蛋白酶L 与其他蛋白等物质的结合作用,起到了抑制其作用活性的目的,与PP1 对组织蛋白酶L 的抑制作用原理是相同的。图8 所显示的是二者结合相互作用位点附近的关键氨基酸。抑制剂与蛋白酶之间相互结合的分子间作用力只有氢键,键长为2.443 32 Å,肽作用位点氨基酸为GLN21,组织蛋白酶L 作用位点为H243,作用活性位点只有一个的原因可能是PP2 与组织蛋白酶L 的相互结合的关系不紧密或PP2 对组织蛋白酶L 的抑制作用不明显,作用力较弱,需要进一步的实际实验来进行验证和深入的研究。
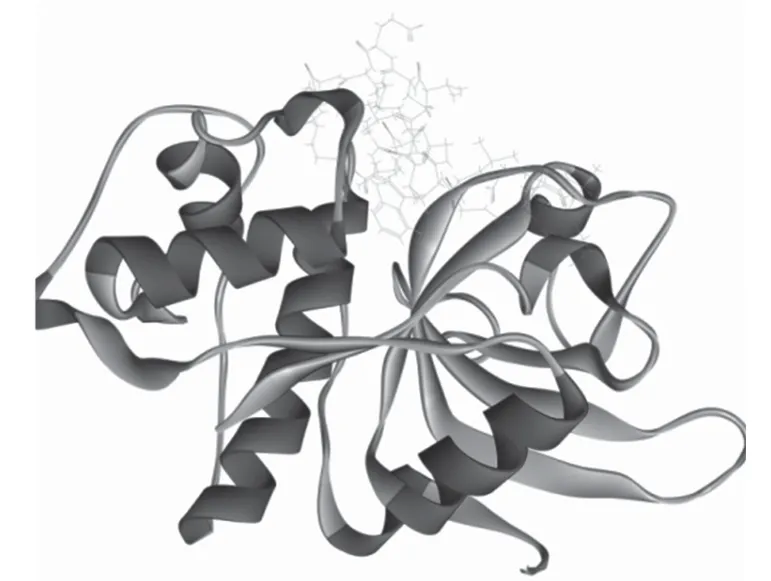
图7 PP2 与组织蛋白酶L 相互作用的分子对接模型
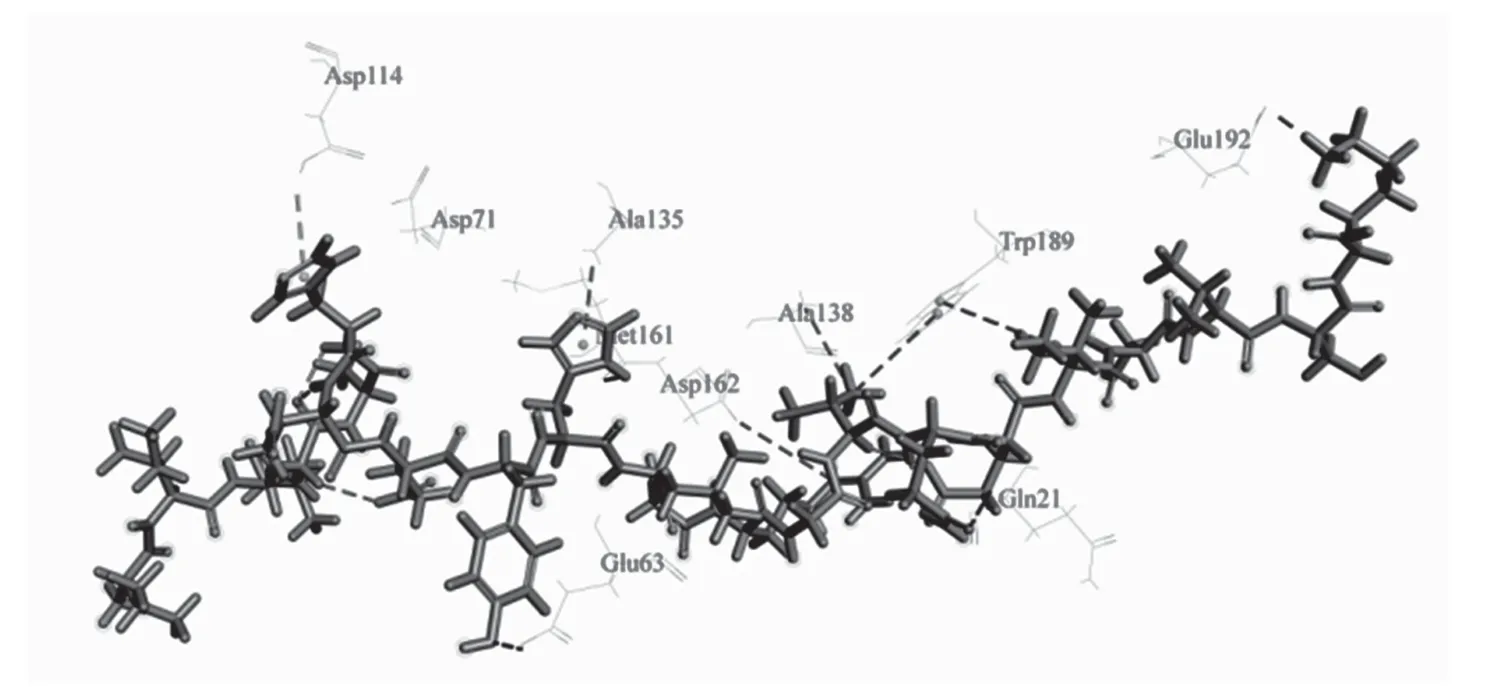
图8 PP2 与组织蛋白酶L 相互作用位点附近的关键氨基酸
3 结论
本研究以猪血浆为研究对象,将其血浆蛋白经过硫酸铵盐溶液分级沉淀、胰蛋白酶酶解,再利用阴离子交换层析和超滤分级的方法对其中的半胱氨酸蛋白酶抑制肽组分进行分离纯化,发现具有少量负电荷的多肽组分、分子量小于3 kDa 的组分具有更高的酶活抑制率,最高抑制率达35.15%,再通过UPLC-QTOF 联用质谱鉴定分析,然后在NCBI 中检索比对抑制肽的氨基酸序列,并从中筛选出具有有效成分的多肽氨基酸序列PP1 和PP2,通过抑制肽与组织蛋白酶L 模拟分子对接分析抑制肽和组织蛋白酶L 的结合作用机制,发现PP1 抑制肽可与组织蛋白酶L 通过活性中心紧密结合,模拟结果显示PP1 与组织蛋白酶L 之间存在13 个作用位点,且分子间作用力主要为氢键,极少部分为静电力。上述研究结果为猪血浆蛋白高效综合利用提供了一定的理论基础。
