睡莲品种‘公牛眼’和‘泰国王’授粉后子房的发育差异研究
2022-04-27唐毓玮李佳慧毛立彦黄秋伟龙凌云於艳萍苏群
唐毓玮 李佳慧 毛立彦 黄秋伟 龙凌云 於艳萍 苏群









摘 要:睡蓮属()植物是我国新兴的水生花卉,其中重瓣型睡莲的花态丰满艳丽,一直倍受睡莲爱好者、育种者的喜爱和关注,在水生园林景观中的应用日渐广泛,但在育种的过程中,育种者经常遇到杂交不结实的现象,导致重瓣型睡莲品种难以培育。为探索重瓣睡莲难以结实的原因,以重瓣睡莲中较具代表性的品种‘泰国王’为实验组,以育性较好的品种‘公牛眼’为对照组,利用石蜡切片技术对其授粉后不同时期的子房进行显微结构比较,采用扫描电镜进一步观察样本的柱头和胚珠,通过RNA-seq测序分析发育与败育子房的基因表达差异。结果表明:2个睡莲品种的柱头表面均分布着多细胞单列乳突,细胞间的连接处形成一圈圈凹槽,乳突数量庞大、排列紧密,配合自身的结构能更容易捕获外来的花粉。‘公牛眼’授粉7 d后子房中大部分胚珠发育成红色种子,10 d后子房膨大成果实,种皮由红色转黑色,形成成熟的种子;重瓣睡莲‘泰国王’授粉7 d后无种子形成,子房中有部分胚珠发育,其表皮变为红色,形态特征与‘公牛眼’授粉4 d后的红色胚珠相似,由此推测少量胚珠可能完成了受精,但无法进一步发育,10 d后子房完全败育。显微结构观察可知,授粉7 d后2个品种的胚珠发育产生明显差异,‘公牛眼’的胚珠明显增大,合子开始分裂形成胚,而‘泰国王’胚珠内珠心萎缩,胚珠逐步败育消解,授粉10 d后,‘公牛眼’胚珠的珠被增厚形成种皮,胚开始发育,进一步向种子形态转变。由转录组测序分析可知,差异基因显著富集在光合作用——天线蛋白、光合作用、卟啉和叶绿素代谢、核糖体、淀粉和蔗糖代谢等通路,这些通路可能是影响睡莲子房发育的关键途径,推测‘泰国王’子房败育可能是因为自身光合作用受到抑制,进而影响了蔗糖、淀粉等营养物质的生成。
关键词:睡莲;杂交授粉;子房发育;胚珠;转录组中图分类号:S682.32 文献标识码:A
Differences in Ovary Development Between ‘Bull’s Eye’ and ‘King of Siam’ after Pollination
TANG YuweiLI JiahuiMAO LiyanHUANG QiuweiLONG LingyunYU YanpingSU Qun
1. Guangxi Subtropical Crop Research Institute, Nanning, Guangxi 530001, China; 2. Flower Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
(water lily) is an aquatic flower that has been introduced in recent years in our country. In particular, double petal water lily are very popular and full of love and attention by water lily lovers and breeders. And it has been increasingly widely used in the field of aquatic garden landscapes. But in the process of breeding, breeders often encounter the situation of failed hybridization. This makes it hard for breeders to breed double flower. In order to study on the reason that double petal water lily is difficult to set fruit, we selected the more representative cultivar ‘King of Siam’ from double petal water lily as the experimental group, and the better fertile cultivar ‘Bull’s Eye’ as the control group. Firstly, the microstructure of ovaries at different stages after pollination was compared by paraffin section technique. Secondly, the stigmas and ovules were further observed by scanning electron microscope. Finally, the differences of gene expression between developing and abortive ovary were analyzed by RNA-Seq. The results showed that multicellular uniserial papillae were distributed on the stigma surface of the two water lily varieties, and a circle of grooves were formed at the junction between single cells. The number of papillae was large and closely arranged. We think this structure can more easily capture foreign pollen. Meanwhile, after 7 days of . ‘Bull’s Eye’ pollination, most of the ovules developed into red seeds, 10 days later, the ovary expanded into fruit, and the seed coat changed from red to black to form mature seeds. On the other hand, after 7 days of pollination, no seeds were formed in . ‘King of Siam’. However, some ovules in its ovary develop, and its epidermis turns red. The morphological characteristics are similar to the red ovules after 4 days of . ‘Bull’s Eye’ pollination. Therefore, we speculate that a small number of ovules may have completed fertilization, but can not develop further. Than, the ovary was completely aborted after 10 days. Microstructural observation showed that there were significant differences in ovule development between the two varieties after 7 days of pollination. The ovules of . ‘Bull’s Eye’ increased significantly, and the zygotes began to form embryos, while the nucellus in the ovules of . ‘King of Siam’ shrank and the ovules gradually disappeared. After 10 days of pollination, the embryo of . ‘Bull’s Eye’ began to develop and further changed to seed morphology. In additional, the enrichment analysis of differential genes showed that photosynthesis antenna protein, photosynthesis, porphyrin and chlorophyll metabolism, ribosome, starch and sucrose metabolism may be the key pathways affecting the development of waterlily ovary. Consequently, we speculate that the abortion of the ovary of . ‘King of Siam’ may be due to the inhibition of its own photosynthesis, which affects the production of nutrients such as sucrose and starch.
; cross pollination; ovary development; ovule; transcriptome
10.3969/j.issn.1000-2561.2022.04.019
授粉是种子植物生殖过程中的重要阶段,授粉成功的子房會进一步发育成果实,而个体间不亲和的授粉会影响花粉与雌蕊的识别、受精以及胚珠的发育,进而导致子房败育无法结实。
睡莲是睡莲科(Nymphaeaceae)睡莲属()植物的通称,花色丰富,形态多样,在水生园林景观中应用广泛。国内育种研究始于1999年,育种目标以花色特异性为主。截止2019年,我国已培育成新品种205个,其中国内鉴定18个、国际登陆187个。重瓣型睡莲的花态丰满艳丽,一直倍受睡莲爱好者、育种者的喜爱和关注,但在育种的过程中,育种者经常遇到杂交不结实的现象,限制了重瓣型热带睡莲的杂交育种研究。
‘泰国王’是重瓣型热带睡莲中较具有代表性的品种,本课题组前期进行了杂交试验,发现‘泰国王’授粉以后常出现假膨大的现象,子房略微膨大后衰败,难以结实。本研究选择睡莲‘泰国王’品种和育性较好的‘公牛眼’品种为材料,采用组织学、转录组学等手段研究其授粉后子房发育的差异,探讨重瓣型睡莲授粉后子房败育的成因,以期为睡莲的杂交育种提供理论基础。
材料与方法
材料
实验材料为栽培多年的热带睡莲品种‘公牛眼’和‘泰国王’,种植于广西壮族自治区南宁市兴宁区亚热带作物研究所睡莲种质资源圃(108°38¢E,22°87¢N),实验地属于亚热带季风气候。课题组前期研究发现,‘泰国王’的雄蕊完全瓣化,无法产生花粉,因此不能进行自交授粉,仅能作为母本材料进行杂交,经人工杂交授粉后难以结实,而‘公牛眼’自交结实率较高。
方法
1.2.1 人工授粉 选择健康的‘泰国王’和‘公牛眼’品种植株,以‘公牛眼’为父本,‘泰国王’和‘公牛眼’分别作为母本,采用睡莲的免去雄人工杂交技术进行授粉。
1.2.2 授粉后子房发育的组织学观察 收集授粉后6 h、4 d、7 d和10 d的子房,分别命名为6HAP、4DAP、7DAP和10DAP,用FAA固定液对样本进行固定,采用二甲苯和乙醇进行梯度脱水干燥后用常规石蜡法包埋。用PAS+萘酚黄S试剂进行染色,将切片置于显微镜(Zeiss Primostar 3)下观察拍照记录。
1.2.3 柱头和胚珠的扫描电镜观察 立即采集授粉后的柱头以及采集授粉后7 d的子房,迅速投入电镜固定液,转移至4℃保存。将样本依次于不同梯度酒精脱水后,用临界点干燥仪进行干燥,最后将样本喷金做导电处理,在扫描电镜下观察拍照。
1.2.4 RNA的提取及转录组测序 授粉7 d后分别剪下母本的子房,‘公牛眼’样本为子房发育组(ovary development, OD),‘泰国王’样本为子房败育组(ovary abortion, OA),立即放入液氮中速冻,在转移至超低温冰箱中–80℃保存,用于RNA提取。每个样品取3个生物学重复,共6份材料。用TRIzol试剂盒法(invitrogen, CA, USA),对子房的RNA进行分离和纯化,检测RNA的纯度与完整性。样品RNA合格后,委托联川生物技术股份有限公司构建cDNA文库,使用illumina Novaseq6000 (LC Bio Technology CO., Ltd. Hangzhou, China)平台进行转录组测序。
1.2.5 生物信息学分析 转录组测序后得到的原始数据Raw reads,使用cutadapt软件对原始数据进行去除接头处理,然后对数据进行去除低质量序列和重复序列后得到Valid reads。具体数据预处理步骤如下:(1)去除带接头(adaptor)的reads;(2)去除含有N(N表示无法确定碱基信息)的比率大于5%的reads;(3)去除低质量reads (质量值Q≤10的碱基数占整个read的20%以上);(4)统计原始测序量、有效测序量、Q、Q、GC含量。使用ballgown包进行FPKM定量,FPKM表示在每百万测序碱基中每千个转录子测序碱基中所包含的测序片断数。使用R包edgeR对样本之间进行显著差异分析,将差异倍数FC>2倍或FC<0.5倍且<0.05的基因定义为差异基因,并对其进行GO和KEGG富集分析。
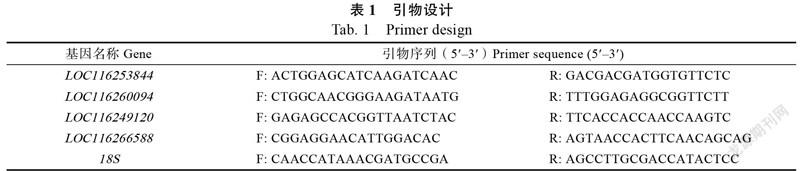
1.2.6 qRT-PCR验证 依据富集结果筛选出差异表达的基因,从中筛选4个设计特异性引物(表1),以基因为内标参照,进行qRT-PCR验证分析,引物由擎科生物技术有限公司合成。qRT-PCR反应体系为10 μL:2×SYBR Green Supermix 5 μL,Forward primer与Reverse primer各0.5 μL,cDNA模板1 μL,ddHO 3 μL。每个样品设置3个生物学重复。利用2方法计算各个样品的基因相对表达量。
结果与分析
‘公牛眼’与‘泰国王’品种子房发育的形态差异
‘公牛眼’具有较好的育性,如图1所示,授粉4 d后(4DAP)的子房略微膨大,靠近柱头处的胚珠最先发育,形成红色的表皮,这些可能是最先成功受精的胚珠。授粉7 d后(7DAP)子房中的大部分胚珠发育成为红色的种子,授粉10 d后(10DAP)子房膨大呈果實(果实横径约3 cm),种皮由红色转为黑色,形成成熟的种子。‘泰国王’育性较差,难以结实,授粉后4 d(4DAP)的子房略微膨大,7 d后(7DAP)柱头表面开始褐变,子房中有部分胚珠发育,其表皮变为红色,形态特征与‘公牛眼’4DAP的红色胚珠相似。‘泰国王’6HAP和4DAP的子房大小与同期‘公牛眼’的无明显差异,直到授粉7 d后子房产生明显的形态差异,‘泰国王’子房中无种子形成,10DAP的花朵完全腐烂。
‘公牛眼’与‘泰国王’品种子房发育的组织学观察
如图2所示,睡莲的胚珠在子房壁上的排列无明显规律。授粉6 h的‘公牛眼’胚珠与‘泰国王’的胚珠发育无明显差异。授粉后4 d,‘公牛眼’的胚珠发育成为长卵圆形,其珠心的面积也随之增加,而‘泰国王’胚珠的珠心开始萎缩。授粉后7 d,2个品种的胚珠发生了明显的差异,‘公牛眼’的胚珠明显增大,此时的珠心充满整个胚珠内部,与珠被紧紧贴合,合子开始分裂形成胚,而‘泰国王’胚珠的珠心继续萎缩呈“十”或“丫”字型,胚珠败育消解。10 d后,‘公牛眼’胚珠的珠被增厚形成种皮,珠心萎缩,胚开始
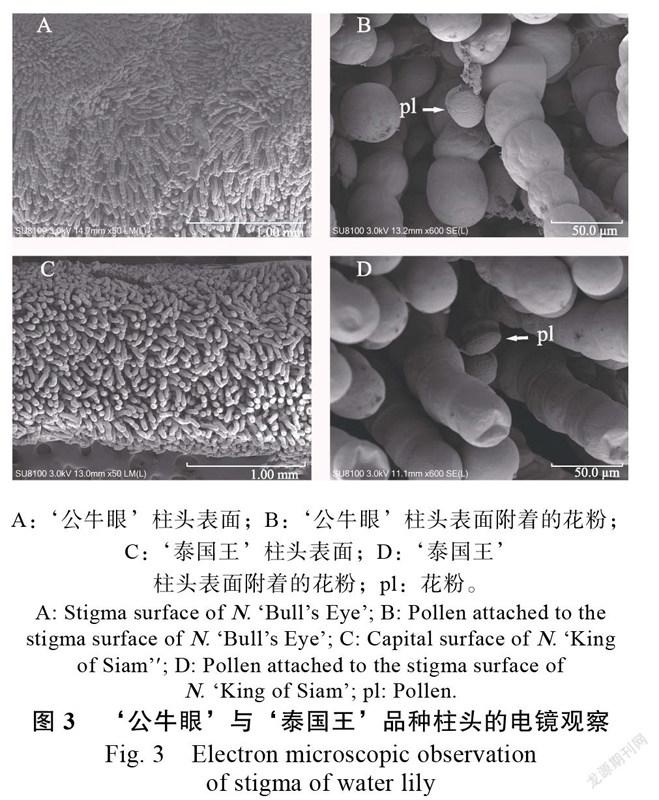
授粉后立即采集的柱头经电镜扫描发现(图3),‘公牛眼’与‘泰国王’柱头表面的显微结构相似,柱头上均分布着大量的乳突,每个乳突由一单列的单细胞组成,基部的细胞体积较大,顶部的细胞体积较小,顶部第一个细胞的表面略微凹陷。乳突上一列单细胞之间的连接处形成了一圈圈的凹槽,花粉易附着在凹槽处。乳突数量庞大、排列紧密,配合自身的结构能更容易捕获外来的花粉。
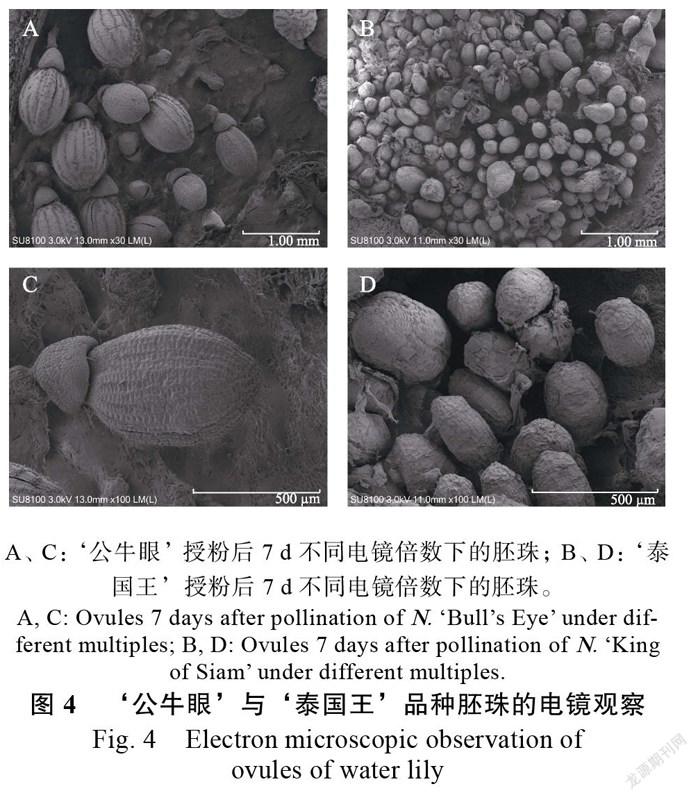
如图4所示,‘公牛眼’授粉7 d后,子房中正常发育的胚珠膨大成为种子,呈卵圆形,悬挂于子房壁上,珠被增厚初步发育成种皮,表面均匀分布着条纹,条纹的方向与胚珠的长轴平行。胚珠与珠柄连接处,存在一个冠状组织盖着胚珠的顶端。而在‘泰国王’的子房中,胚珠的发育程度参差不齐,有的呈卵圆形,有的较为狭长,也有呈近似圆形的,体积大小不一。
‘公牛眼’与‘泰国王’品种子房的转录组分析
2.3.1 转录组测序质量评估及基因表达量分析 经转录组测序共获得282 800 654条原始序列(raw reads),去除低质量序列和重复序列后得到有效序列(valid reads)277 620 180条。从表2
可知,有效序列所占比例均大于97%,质量值不小于20的碱基所占比例均大于99.98%,质量值不小于30的碱基所占比例均大于97%,碱基G和C的数量总和占总碱基数量的48%~49%,说明6个样品的测序质量很高,可进行后续的生物学分析。
将两组样本的基因表达量差异倍数(fold change,FC)大于2或FC小于0.5,且<0.05的基因定义为子房败育组(OA组)与子房发育组(OD组)的差异基因。从结果中可以看出,共有6033个差异基因,其中上调表达的基因3104个,下调表达的基因有2929个(图5)。
2.3.2 差异基因GO富集分析 Gene Ontology(简称GO)基因功能分类是国际化标准的分类体系之一,GO总共有3个ontology(本体),分别描述基因的分子功能(molecular function)、所处的细胞位置(cellular component)、参与的生物过程(biological process)。‘泰国王’和‘公牛眼’被授粉以后,对子房差异表达基因(OA vs OD)进行GO功能分类,可以分为生物过程、细胞组分和分子功能3个大类,3416个小类,图6分别展示的是生物过程、细胞组分和分子功能中基因数量前25、15、10的小类。在生物过程中,差异基因参与数量排名前五的是生物过程(biological process)、转录调控与DNA模板化(regulation of transcription, DNA-templated)、转录与DNA模板化(transcription, DNA-templated)、氧化还原过程(oxidation-reduction process)、蛋白质磷酸化(protein phosphorylation),基因数量分别是656、359、300、248、192个。在细胞组分中,差异基因参与数量排名前五的是细胞核(nucleus)、叶绿体(chloroplast)、细胞质(cytoplasm)、膜的组成部分(integral component of membrane)、质膜(plasma membrane),基因数量分别为1362、800、769、755、737个。在分子功能中,差异基因参与数量排名前五的是分子功能(molecular function)、蛋白质结合(protein binding)、DNA结合转录因子活性(DNA-binding transcription factor activity)、ATP结合(ATP binding)、DNA结合(DNA binding),基因数量分别为648、585、334、325、297个。
如图7所示,在处理组OA和对照组OD中,差异基因GO富集显著性排在前五的分类是叶绿体类囊体膜(chloroplast thylakoid membrane, rich factor = 0.45)、叶绿体(chloroplast, rich factor =0.33)、叶绿体包膜(chloroplast envelope, rich factor=0.40)、光合作用与光系统中的光捕获(photosynthesis, light harvesting in photosystem I, rich factor=0.78)、叶绿素结合(chlorophyll binding, rich factor=0.71)。差异基因富集显著的GO类目均与光合作用息息相关。
2.3.3 差异基因KEGG富集分析 KEGG(kyoto encyclopedia of genes and genomes)全称是京都基因和基因组百科全书,是基因组破译方面的公共数据库。在生物体内,不同基因相互协调行使其生物学功能,基于Pathway的分析有助于更进一步了解基因的生物学功能。如图8所示,在处理组OA和对照组OD中,差异基因富集显著性-value排在前五的KEGG pathway分别为光合作用-天线蛋白(photosynthesis-antenna proteins, rich factor=0.77)、光合作用(photosynthesis, rich factor=0.50)、角质,软木和蜡的生物合成(cutin, suberine and wax biosynthesis,rich factor=0.49)、核糖体(ribosome, rich factor=0.37)、卟啉和叶绿素代谢(porphyrin and chlorophyll metabolism, rich factor=0.48)。在显著富集的pathway(≤0.05)中,差异基因数量最多的3条pathway为核糖体(ribosome)、淀粉和蔗糖代谢(starch and sucrose metabolism)、苯丙烷类生物合成(phenylpropanoid biosynthesis),差异基因的数量分别为122、111和94个。在参与光合作用-天线蛋白、光合作用途径相关的差异基因中,OA组绝大多数基因的表达量下调,仅LOC116246649、LOC116263852、LOC116266250、LOC116260662这4个基因表达量上调(图9),可见OA样本的光合作用受到抑制。
相关基因的表达分析
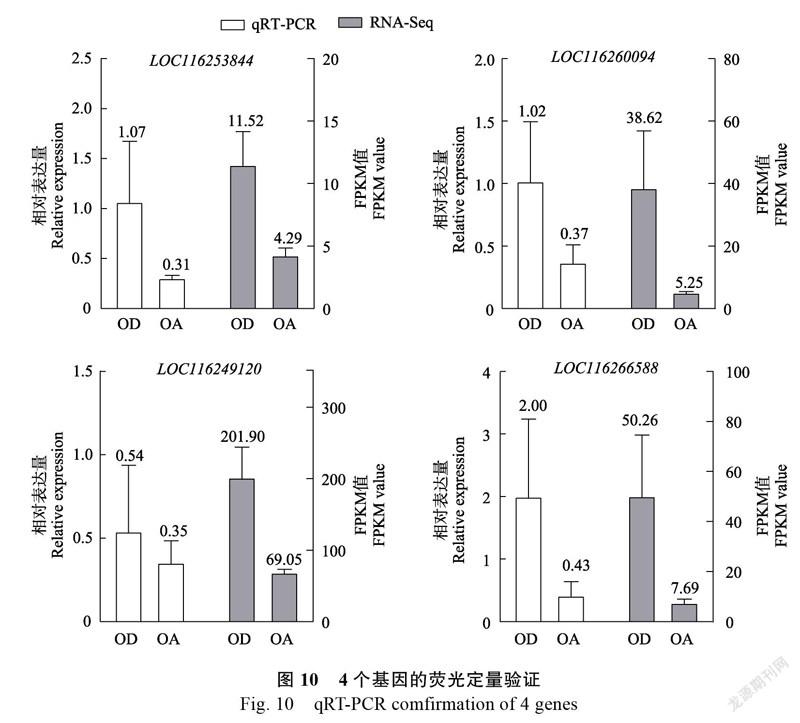
结合相关文献资料发现,PRPL1、L4、L21、L35对拟南芥的胚珠发育至关重要,依据差异基因KEGG富集分析结果,选择相关基因LOC116253844、LOC116260094、LOC116249120、LOC116266588进行验证(表3)。在OA组中,相关基因的表达量均较低,qRT-PCR结果与转录组的基本一致(图10)。
讨论
花粉与柱头间的识别作用是确保亲和花粉萌发、完成受精并保持后代遗传稳定性的重要环节,睡莲的柱头呈盘状,成熟后能分泌大量富含营养物质的液体,为花粉的水合萌发提供了必要的基质,也在花粉与柱头的相互识别过程中起着重要作用。从花朵盛开与闭合的时间上分类,可将睡莲分为夜间开花和白昼开花两大生态类型,夜间开花的热带型睡莲,如印度红睡莲、埃及白睡莲Nymphaea rubra等柱头表面分布着浓密的乳突组织,且每个乳突由一单列的多个细胞组成,属于多细胞单列乳突的湿柱头类型。本研究中,对白天开花的热带型睡莲‘公牛眼’和‘泰国王’品种的柱头表面进行电镜观察,得到了较一致的结果,柱头表面分布着浓密的多细胞单列乳突,形似“葫芦状”,许多花粉很容易附着于多个细胞的连接处。芡实、萍蓬草等植物的水生植物柱头上仅仅隆起单个乳突,柱头结构可能为原始类型,而睡莲属的多细胞单列乳突的柱头可能为进化类型,本文认为后者能更容易附着外来花粉。
通过对‘公牛眼’和‘泰国王’子房的形态观察发现,结实率低的‘泰国王’子房中有少量胚珠发育成熟,其表型与‘公牛眼’红色胚珠相似,由此推测少量胚珠可能完成了受精,但‘泰国王’的子房最终败育腐烂,受精的胚珠无法进一步发育为成熟的种子。DENG等在菊花杂交中发现大约有75%的胚珠能完成受精,但最终只有17.2%的胚珠形成成熟的胚胎,高比例的胚胎败育是种子产量低的主要原因。胚珠受精后形成胚胎,由于营养供给不足等原因,很容易发生胚胎和内外胚乳组织发育异常、细胞核降解等情况,导致胚胎败育。睡莲子房中的胚珠多达上千粒,对于少量成功受精的胚珠,幼胚拯救是目前常用的技术方法。
本研究中,‘泰国王’和‘公牛眼’授粉后7 d的子房表型和子房內部的胚珠发育均发生了较大差异,因此选择授粉后7 d的子房进行转录组测序,构建了发育组(OD)和败育组(OA)睡莲子房cDNA文库。
光合作用是植物重要的生理过程,对许多作物早期的生殖发育也起到重要的作用。榛子发育子房中光合作用相关的差异表达蛋白均显著上调表达,表明光合作用对子房的发育有所促进。光合作用对番茄果实内种子的发育也发挥了重要作用,LYTOVCHENKO等发现,叶绿素合成受到抑制的转基因番茄无法形成健康的种子。PATTISON等也证实在番茄果实早期发育时,大量与光合作用相关的基因在番茄胎盘和隔膜中表达,同时通过淀粉染色发现淀粉颗粒主要集中在胎盘和隔膜中。GAO等发现,光合作用相关基因的启动与拟南芥胚胎发育早期有关,贮藏、能量相关蛋白的激活与胚胎后期发育相关。光合产物主要以蔗糖和淀粉的形式存在,蔗糖和淀粉的有效利用对于配子体发育、受精以及子代和母体的协调发育至关重要。在玉米子房中,蔗糖为子房壁和花梗中的淀粉积累提供碳源,并且被花梗中的细胞转化酶(CWIN)和珠心组织中的液泡转化酶(VIN)水解,产生高浓度的葡萄糖流入胚囊,促进胚珠的发育。本研究中差异表达基因的GO和KEGG富集结果显示,差异表达基因在光合作用-天线蛋白、光合作用、卟啉和叶绿素代谢、核糖体、淀粉和蔗糖代谢等通路上显著富集,表明这些通路可能是影响睡莲子房发育的重要途径,‘泰国王’子房的败育可能是因为自身光合作用受到抑制,进而影响了蔗糖、淀粉等营养物质的生成。
核糖体蛋白PRPS20、L1、L4、L27和L35是维持核糖体活性所必需的,这在拟南芥胚胎发育从球形向心形阶段的过渡中变得至关重要,缺乏这些PRP蛋白会导致细胞分裂模式的改变,拟南芥胚胎发育到心形阶段之前就会停止。YIN等发现,叶绿体核糖体蛋白L21基因也是拟南芥胚胎发生所必需的。本研究选择了4个与PRPL1、L4、L21和L35蛋白相关的基因进行验证工作,4个基因在败育子房中均显著下调,由此可见,这些蛋白对睡莲胚的发育可能也有重要的调控作用。
本研究采用形态学方法确定了子房败育的关键时期,进一步利用RNA-seq测序分析初步探讨导致睡莲子房败育的可能原因,但仅仅从基因表达层面仍然不能充分阐明睡莲子房败育的机理,后续研究可采用多组学联合分析,挖掘并验证影响子房发育的关键基因,以系统阐明睡莲子房败育的机制。
参考文献
- 黄 岩, 邢世岩, 付兆军, 李 真. 叶籽银杏授粉期胚珠的形态结构变化[J]. 园艺学报, 2013, 40(2): 205-212.HUANG Y, XING S Y, FU Z J, LI Z. Observation on changes of morphological structure of the ovules during pollination period in L. var. [J]. Acta Horticulturae Sinica, 2013, 40(2): 205-212. (in Chinese)
- 齐秀娟, 徐善坤, 张威远, 林苗苗, 方金豹. 美味猕猴桃‘徐香’与长果猕猴桃远缘杂交亲和性的解剖学研究[J]. 园艺学报, 2013, 40(10): 1897-1904.QI X J, XU S K, ZHANG W Y, LIN M M, FANG J B. Studies on compatibility of interspecific hybridization between actinidia diliciosa ‘Xuxiang’ and by anatomy[J]. Acta Horticulturae Sinica, 2013, 40(10): 1897- 1904. (in Chinese)
- 王文鹏, 周莉花, 刘华红, 包志毅, 赵宏波. 夏蜡梅与美国蜡梅属间杂交障碍的组织学机理[J]. 园艺学报, 2013, 40(10): 1943-1950.WANG W P, ZHOU L H, LIU H H, BAO Z Y, ZHAO H B. Histological reproductive barriers for intergeneric cross between and var. [J]. Acta Horticulturae Sinica, 2013, 40(10): 1943-1950. (in Chinese)
- 解玮佳, 李世峰, 瞿素萍, 彭绿春, 张 露, 杨秀梅, 王继华. 常绿杜鹃不同杂种群间杂交的可育性分析[J]. 园艺学报, 2019, 46(5): 910-922.XIE W J, LI S F, QU S P, PENG L C, ZHANG L, YANG X M, WANG J H. Cross fertility of the crosses between different hybrids groups[J]. Acta Horticulturae Sinica, 2019, 46(5): 910-922. (in Chinese)
- 牛红云, 王 臣, 薛贵彬, 胡宝忠. 四种寒带睡莲的杂交亲合性研究[J]. 北方园艺, 2011, 233(2): 100-102.NIU H Y, WANG C, XUE G B, HU B Z. Study on affinity of sexual hybridization in four taxa of northeast[J]. Northern Horticulture, 2011, 233(2): 100-102. (in Chinese)
- ISIDORA R, LUCA T, FABIO R, SIMONA M, MATHIAS P, PETER J, MARTIN K, DARIO L, PAOLO P. Versatile roles of plastid ribosomal proteins in plant growth and development[J]. The Plant Journal, 2012, 72(6): 922-934.
- YIN T Z, PAN G, LIU H, WU J, LI Y P, ZHAO Z X, FU T D, ZHOU Y M. The chloroplast ribosomal protein 21 gene is essential for plastid development and embryogenesis in [J]. Planta, 2012, 235(5) : 907-921.
- 程 茵, 马光耀, 赵 莹, 盛玉辉, 李雪青, 周 扬, 宋希强, 王 健. 睡莲花露主要成分及抗氧化作用[J]. 食品研究与开发, 2019, 40(8): 108-114.CHEN Y, MA G Y, ZHAO Y, SHEN Y H, LI X Q, ZHOU Y, SONG X Q, WANG J. The main metabolites and antioxidation effect of waterlily stigmatic exudata[J]. Food Research and Development, 2019, 40(8): 108-114. (in Chinese)
- DENG Y M, TENG N J, CHEN S M, CHEN F D, GUAN Z Y, SONG A P, CHANG Q S. Reproductive barriers in the intergeneric hybridization between (Ramat.) Kitam. and Poljak (Asteraceae)[J]. Euphytica, 2010, 174(1): 41-50.
- 孙春青, 陈发棣, 房伟民, 刘兆磊, 马 靜, 滕年军, 佘琳芳. 栽培菊花‘奥运天使’与野路菊杂交生殖障碍的细胞学机理[J]. 中国农业科学, 2009, 42(6): 2085-2091.SUN C Q, CHEN F L, FANG W M, LIU Z L, MA J, TENG N J, SHE L F. Cellular mechanism of reproductive barrier during cross breeding between cvAoyuntianshi and [J]Scientia Agricultura Sinica, 2009, 42(6): 2085-2091. (in Chinese)
- GAO P, XIANG D, QUILICHINI T D, VENGLAT P, PANDEY P K, WANG E, GILLMOR C S, DATLA R. Gene expression atlas of embryo development in [J]. Springer Berlin Heidelberg, 2019, 32(1): 93.
