乳源外泌体研究进展
2022-04-24付子琳和俊豪段晓微卜登攀
丁 军,付子琳,和俊豪,段晓微,马 露,卜登攀,2*
(1.中国农业科学院北京畜牧兽医研究所 动物营养学国家重点实验室,北京 100193;2.中国农业科学院与世界农用林业中心农用林业与可持续畜牧业联合实验室,北京100193)
外泌体(exosomes)存在于正常生理或者病理条件下的各种体液中,如血液[1]、尿液[2]、脑脊液、支气管灌洗液、精液[3]、腹水、汗液、唾液、泪液、乳汁和淋巴液等[4-5]。它是由细胞分泌到胞外的纳米级囊状小泡,外包磷脂双分子层,内含核酸、蛋白质和磷脂,呈类细胞的拓扑结构,大小在30~150 nm之间[6]。与其他类型的细胞外囊泡(微囊泡、凋亡小体)相比,外泌体在大小、生物合成和内含物上存在不同[7]。外泌体,最早(1983年)是由Pan等[8]研究体外培养的绵羊网织红细胞向成熟红细胞分化过程中,在细胞上清液中发现并分离得到的一种30~150 nm的细胞外膜性小囊泡。之后研究发现,这种细胞外囊泡是受调控的,非随机地包裹mRNAs、miRNAs、DNA、脂质和蛋白质等物质[9-12],并通过胞吐、融合等方式把内含物释放到细胞外,然后由临近的或远隔的受体细胞接受信号而发挥生理维护或者病理修复的作用[13]。外泌体作为细胞间信号标志物之一,在细胞间信号传导上拥有极大的生物学价值[14-15],同时在疾病的诊断、治疗和预防上也发挥极大的作用。例如在阿尔茨海默病和帕金森综合征的诊断中,外泌体参与了β-淀粉样蛋白和α-突触核蛋白的传播[16-18];在癌症的研究上,外泌体还具有抗肿瘤[19]及肿瘤生物标记作用[20]。
1 外泌体的产生
几乎所有的细胞都可以产生细胞外囊泡(extracellular vesicles,EVs),但是外泌体不同于其他的细胞外囊泡,其大小在30~150 nm,呈囊泡状。外泌体的生成是从细胞的内出芽(胞吞)开始,然后依次形成早期内吞小体(early endosome)、晚期内吞小体(late endosome)。伴随着细胞的代谢,晚期内吞小体的脂膜内陷,开始有选择地分拣RNAs、miRNAs、脂质和蛋白质等内含体[21-22]。目前的研究发现,外泌体的产生主要有两种途径(图1):1)晚期内吞小体在囊泡分拣蛋白如转运必需核内复合物(endosomal complex required for transport,ESCRT)的作用下通过识别、分类、挑选内含物而生产多囊泡体(multivesicular body);2)在神经酰胺的协助作用下晚期内吞小体生成多囊泡体。有的多囊泡体会和溶酶体融合,然后被分解,有的和细胞膜融合,释放到细胞外形成外泌体[6, 23-25]。在乳中,外泌体来源于乳腺上皮细胞和一些免疫细胞[26]。日本科学家Hata等[27]通过乳脂肪球表皮生长因子8(milk fat globule epidermal growth factor 8,MFG-E8)确定了乳源外泌体来源于乳腺上皮细胞。最近,中国学者分离获得牛乳腺上皮细胞来源的外泌体,并利用蛋白组学技术结合现有牛乳源外泌体库进行了鉴定[28]。

图1 外泌体的生物合成和释放[6]
2 乳源外泌体的结构和功能
哺乳动物的乳汁是新生动物最主要的营养来源,乳汁中含有新生动物所需的所有营养物质,包括蛋白质、脂肪、矿物元素、维生素及各种活性成分等,能够促进动物个体生长发育和机体免疫力等功能[29]。有研究发现,在哺乳动物的乳汁中存在外泌体,并已证实其具有生物活性[6]。乳源外泌体在疾病诊断、治疗、预防以及人和动物的生长发育等方面都具有重要的研究价值。牛乳是外泌体的重要来源[30],牛奶样品容易获取,并且不会对奶牛产生负面影响,这为乳源外泌体的研究奠定了基础。
乳源外泌体和其他来源外泌体在结构上相似,都具有与细胞相同的拓扑结构和脂质双分子层,膜上具有膜蛋白,内含物中存在大量的蛋白质、核酸和脂质[31-32]。
乳源外泌体中的蛋白质大致分为两类,第一类是膜上的膜蛋白。其膜表面既有与其他源外泌体相同的保守膜蛋白,如CD6、CD81、CD9等[33],又有与其起源细胞密切相关的相对特异性蛋白,如MFG-E8[27]。目前研究发现,在外泌体表面还有一类蛋白,其功能是阻止巨噬细胞识别和清除外泌体,如跨膜蛋白CD47[34]。第二类是内含物蛋白,外泌体内部通过非随机、受调控的方式可以包裹原细胞的蛋白,如热休克蛋白和肌动蛋白等。目前,国际上外泌体的统计数据库主要是ExoCarta[35]和Vesiclepedia[36]。根据ExoCarta统计,从不同来源的外泌体中共检测发现了41 860种蛋白[37]。现已发现,人乳外泌体中含1 963种蛋白,牛乳外泌体中含有1 351种[38],值得注意的是,牛乳中有198种蛋白在人乳中未被检测到[39]。上述研究表明,乳源外泌体中存在大量的蛋白质,这些蛋白质可能会发挥某些生理或病理作用,需要更深入研究。
乳源外泌体中另一类重要组分是核酸,包括DNA、mRNA、miRNA和其他非编码RNA[40]。由于脂质双分子层的保护,乳源外泌体中的核酸可以稳定存在。目前,乳源外泌体中DNA的功能还不清楚,而大量的研究集中在乳源外泌体RNA的研究上。Izumi等[41]在牛乳乳清外泌体中提取鉴定了大约19 000种mRNA和2 838种miRNAs,约占乳源外泌体总RNA的13%[42]。另有研究显示[43],牛乳中大约有400种miRNA的核苷酸序列和人乳外泌体相同,且牛乳来源的外泌体可以被人体巨噬细胞吞噬[41,44],表明牛乳外泌体来源的miRNA可能会被人体吸收而发挥作用。林德麟等[45]统计分析了不同动物乳中miRNAs的含量和种类,发现miR-148a、let-7家族、miR-182-5p、miR-378、miR-141、miR-30a-5p、miR-27b-3p、miR-146b-5p、miR-26a等能在动物乳中高表达,尽管这些乳源miRNAs的生物学功能尚未证实,但Gu等[46]在人、猪和牛乳源外泌体中发现了包括miR-148a和let-7a等与免疫相关的miRNA。
关于乳源外泌体脂质类内含物的报道较少。目前有研究对来源于早产儿和足月儿健康母亲的乳源外泌体进行了分离,并通过LC-MS阐明了其脂质表达谱,共鉴定出395种脂质,共15个脂质亚类,包括磷脂酰胆碱(PC)、磷脂酰丝氨酸(PS)和磷脂酰乙醇胺(PE),通过生物信息学分析发现其中有50种脂质(PC(18:0/18:2)、PC(18:1/16:0)、PS(18:0/18:1)、PS(18:0/22:6)等)通过细胞外信号调节丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)途径调节肠上皮细胞功能[47]。
3 乳源外泌体的分离鉴定
关于外泌体的分离依然缺乏公认方法。目前常用的分离方法有超速离心法[48]、尺寸排阻法、免疫亲和捕获法、试剂盒法[49]和微流体技术等[50-52]。这些分离方法得到的外泌体纯度和浓度不高[48],同时均存在一定的优缺点[49]。超速离心法和试剂盒法是外泌体分离提取最常用的方法。超速离心法操作难度低,分离耗时短[53],该方法利用乳汁样品中外泌体、蛋白质、脂肪、细胞和细胞碎片等物质沉降速度的差异, 通过控制不同的离心力和离心时间, 将其划分为上清液和沉淀,然后进行分离。试剂盒法又被称为聚合物沉淀法,其主要是利用聚乙二醇或其类似物将外泌体沉淀,然后使用低速离心分离外泌体,这样避免了因差速离心耗时而提高了外泌体的分离效率[54]。范士杰等[48]比较了差速离心法和试剂盒法提取娟珊牛乳外泌体,发现试剂盒法提取的外泌体数量多且无细胞毒性。一般的单一分离方法并不能准确获得单一的外泌体,因此,多技术联合使用,会更容易得到单一的外泌体。Abramowicz等[50]通过研究发现,超速离心结合碘克沙醇密度梯度离心或凝胶过滤,尽管劳动强度大,但可以获得高质量的外泌体。Somiya等[55]将样品进行酸化处理,然后再进行超速离心,也获得了单一的、无细胞毒性的外泌体。不同分离方法所得到的乳源外泌体的特征有所不同(表1)。且由于乳汁不同于其他体液,其成分复杂,含有大量小分子的乳蛋白和乳脂,有些小分子物质的大小与外泌体相似,这导致分离获得的乳源外泌体浓度和纯度较低。到目前为止,还没有公认的可以获得高纯度、浓度和生物活性的分离方法,因此对于乳源外泌体的分离技术还有待进一步优化研究。

表1 不同分离方法获得的乳源外泌体特性
乳源外泌体的鉴定需要从两方面进行:首先是物理特性的鉴定,利用电子显微镜、动态光衍射(dynamic light scattering, DLS)和纳米粒子跟踪分析(nanoparticle tracking analysis, NTA)技术,确定乳源外泌体的形态和大小,确定其基本的囊泡状结构,这被称为外泌体检测的金标准[58-59]。在电子显微镜下,几乎所有表征都显示乳源外泌体呈杯状结构,大小在30~150 nm之间。DLS是利用分子的布朗运动捕捉光散射,然后计算外泌体大小的方法,其对于单一尺寸的分子检测较为准确。由于外泌体大小不一,导致其检测结果准确率降低[49]。NTA是一种直接可视化的光学粒子追踪技术,它可以同时显示外泌体的浓度和大小。其最小检测单位是50 nm的外泌体[60-61]。其次再利用蛋白质免疫印迹技术(Western blot)和酶联免疫法通过外泌体表面特异性标志蛋白,如:四跨膜蛋白(CD9,CD63和CD81)、热休克蛋白HSP70、Alixs等的免疫结合进行鉴定核实[62]。同时也可以对外泌体中的特异性内含物进行鉴定[63]。此外,还有研究人员通过对外泌体膜上表面标志物进行标记,然后利用流式细胞术(flow cytometry, FCM)检测阳性表达,以此对外泌体进行定性和定量[64]。
4 乳源外泌体的应用
4.1 潜在的乳房炎诊断标志物
在一些疾病中,乳源外泌体会随着疾病的发生发展而发生变化,这为外泌体作为疾病诊断标志物奠定了基础[65]。在奶牛乳房炎的研究中,研究人员通过培养牛乳腺上皮细胞,然后利用热灭活大肠埃希菌刺激,结果发现,乳腺上皮细胞中有1 478条基因发生差异表达。GO分类富集分析发现,其中有21条基因定位于外泌体,这在细胞层面上证明乳腺炎病原感染可引起宿主细胞所分泌的外泌体成分发生改变[66]。Reinhardt等[32]通过采集健康牛和患乳腺炎牛的乳汁分析乳源外泌体也证实了这一点。在组学分析上,有研究者通过对健康牛和患乳腺炎牛的乳源外泌体进行了全面RNA的分析,检测到492个 已知的和980个未知的miRNAs,其中健康牛和患乳腺炎牛的乳源外泌体差异表达的miRNA有18个,显著差异表达的有2个,分别是miR-223和miR-142-5p,这被视为乳腺炎诊断的潜在候选者[67-68]。张修奇[69]利用差速离心法分离了健康牛和乳房炎牛的乳源外泌体,再次证实了miR-223和miR-142-5p在乳房炎奶牛乳源外泌体中显著表达,然后又补充了miR-146a和miR-29a为牛乳房炎潜在候选者。
4.2 乳源外泌体参与疾病治疗
4.2.1 乳源外泌体与药物 外泌体存在于各种体液中(血液、尿液、乳汁、组织液等),乳汁中外泌体具有量多、易提取、更易使人接受等优点,是配合药物生产的最佳原料。其次,其天然的脂质双层结构可以保护内含物免受破坏、传递酶类和遗传物质、介导细胞间信号传导[70]、递呈抗原和免疫应答[71]。在医学载药领域,目前主要应用的载体是脂质体和聚合物载体,外泌体和这些载体相比,其具有更强的稳定性、生物相容性、靶向性和可改造性,这使得药物通过外泌体能够更加稳定和准确地作用于靶位[72]。外泌体属于纳米级的脂质体,药物借助外泌体后,可以更好地被吸收。另外,外泌体无免疫原性,可以透过血脑屏障,这使得其可以大大提高药物的血脑屏障透过率。据报道,目前外泌体可以用于递送:siRNA、miRNA、蛋白质、小分子药物和纳米颗粒等,用于治疗各种疾病[73]。有研究人员对牛乳源外泌体分离纯化后与紫杉醇共孵育,获得了粒径108 nm的乳源外泌体-紫杉醇复合物,这使药物的生物利用度显著提高,同时降低了用药剂量和毒性[74]。还有研究通过将包裹siRNAs的外泌体复合物用于研究胰腺癌小鼠模型,发现其可以特异性地靶向抑制癌细胞[75]。Kim等[76]通过将姜黄素、RNA结合蛋白(RBP)和外泌体膜蛋白结合制成工程化外泌体,然后通过气管滴入急性肺损伤模型,发现结合外泌体的姜黄素药物效果显著好于单独使用姜黄素。外泌体作为药物载体可以极大程度地协助药物运输,使其更好地发挥作用,是药物运输最佳的载体[77]。但是外泌体载药也存在一些不足,其载药效率较低,需低温保存,并且临床产量不足,这将是未来乳源外泌体载药面临的最大挑战。
4.2.2 乳源外泌体与疾病 乳源外泌体还参与一些疾病的发生发展。在感染性疾病中,外泌体的大小与某些病毒非常相近,其次,其脂膜上包含有参与细胞间信号传导的膜蛋白,内含遗传物质,甚至可以包裹整个病毒[78]。Näslund等[79]研究人乳源外泌体和血浆外泌体对I型人类免疫缺陷病毒(Human Immunodeficiency Virus Type 1, HIV-1)感染的作用时,发现人乳源外泌体可以通过与HIV-1竞争与单核细胞来源的树突状细胞上的受体DC-SIGN结合而抵抗HIV-1垂直传播。Yun等[80]通过使用糖皮质激素诱发了小鼠骨质疏松,然后将分离纯化后的牛乳源外泌体给小鼠口服,发现与未经外泌体治疗的糖皮质激素诱导的骨质疏松组相比,外泌体喂养小鼠组的骨矿物质密度显著提高。在炎症性疾病的研究中,研究人员使用胶原蛋白诱导制造了自发性关节炎的小鼠模型,然后给该小鼠模型口服牛乳源外泌体,发现牛乳外泌体处理使单核细胞趋化蛋白-1(monocyte chemotactic protein 1,MCP-1)和IL-6的血清水平以及脾细胞产生的水平下降[63]。产生这种现象的机制可能与牛乳源外泌体的内含物miR-148a[81]、miR-21[82]以及TGF-β[83]有关。Gao等[84]通过LPS致炎大鼠结肠上皮细胞(IEC-6),然后添加牛乳源外泌体和牦牛乳源外泌体,发现牦牛乳和牛乳外泌体通过激活 PI3K/AKT/C3 信号通路,促进 IEC-6 存活,并且显示CD46是减轻IEC-6细胞炎症的调节剂。另外,牛乳源外泌体的内含物(miRNA)可能参与疾病的发生发展,如前列腺癌、乳腺癌[85]、结肠癌、肝细胞癌[86]、鼻咽癌、B淋巴细胞瘤、肥胖以及II型糖尿病[87](表2)。综上所述,乳源外泌体主要是通过其内含物蛋白质和miRNAs在一些疾病中发挥作用,但是其内含物种类较多,是否是单一内含物发挥作用,还有待进一步研究。
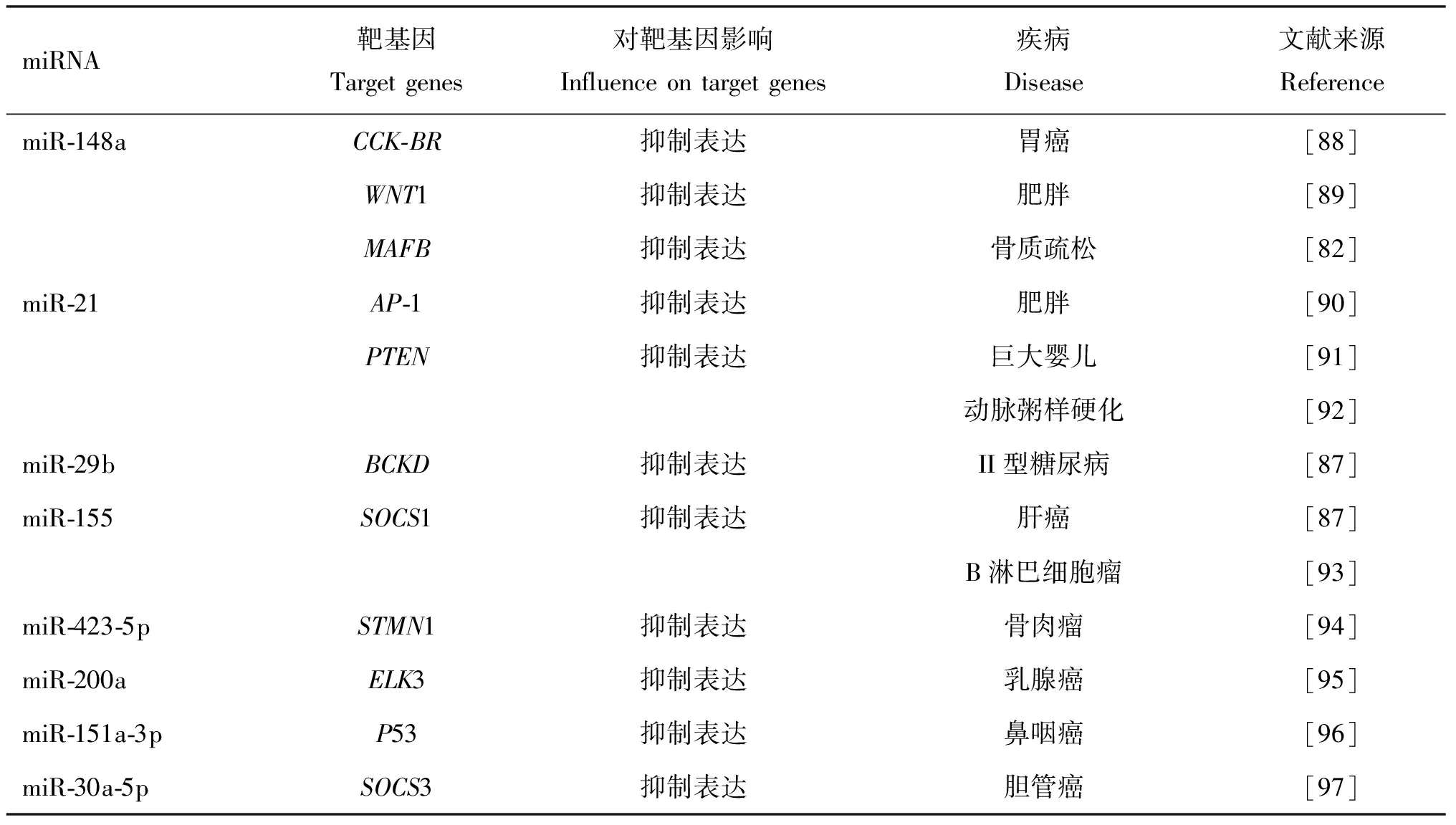
表2 乳源外泌体中miRNA相关的疾病
4.3 疾病预防
初乳中含有免疫球蛋白,可直接被新生动物吸收,提供被动免疫获得抗体,抵抗病原感染。而外泌体作为近年来发现的内含蛋白质和遗传物质的细胞外囊泡,也可以通过其内含物为幼畜的疾病预防提供帮助。乳源外泌体内含与免疫相关的miRNAs,现已发现在人、猪、牛、山羊和熊猫的乳汁中有相同保守的miRNAs如:let-7a、let-7b、let-7f和miR-148a,可能参与调节机体的免疫[65]。研究发现,人乳源外泌体可以抑制白介素-3和干扰素的产生,从而使调节性T细胞的数量增加,调节新生儿的免疫系统发育[98],可预防新生儿坏死性小肠结肠炎(necrotizing enterocolitis,NEC)、支气管肺发育不良(broncho-pulmonary dysplasia,BPD)等相关疾病的发生,同时在生命早期,其还具有预防过敏的潜力[99],但作用机制还不太清楚。Baier等[100]通过交叉试验设计,对26~49岁的3名男性和2名女性给予牛奶饮食,发现牛乳源外泌体中的miRNA可以补偿人内源性miRNA。
4.4 乳源外泌体与肠道发育
在肠道发育方面,乳源外泌体也具有至关重要的作用[101-102]。乳源外泌体可以耐受蛋白酶和核糖核酸酶消化,能够顺利通过消化道并被吸收[103]。有研究显示,通过给小鼠口服饲喂猪乳源外泌体,小鼠十二指肠和空肠的绒毛高度、隐窝深度和绒毛高度与隐窝深度之比(villus height/crypt depth,V/C)显著增加[104]。另有研究通过低蛋白饮食诱导小鼠形成营养不良,然后口服牛乳外泌体,发现牛乳外泌体组的小鼠肠上皮通透性、屏障功能和肠细胞增殖较对照组显著改善[105]。在人体肠道发育方面,牛乳源外泌体被证实可以通过抑制p53(tumor suppressor gene p53,p53)蛋白表达,增加 IEC-6 细胞的存活,以此调节肠道的屏障功能[106-107]。人乳源外泌体通过其内含物circRNAs与相关miRNAs结合,诱导人结肠上皮细胞(FHC)增殖和迁移[108]。Zeng等[109]通过提取猪乳源外泌体,然后灌服小鼠和仔猪,发现乳源外泌体通过其包含物circ-XPO4(circRNA)抑制肠上皮细胞中miR-221-5p的表达来促进分泌性免疫球蛋白A(secretory immunoglobulin A,SIgA)的表达。
乳源外泌体还可以影响肠道微生物,以此调节肠道健康[58]。有研究通过给小鼠饲喂正常牛乳和破坏外泌体结构的牛乳,对比两组小鼠肠道微生物的生长情况,发现正常牛乳显著提高了小鼠肠道微生物的丰度[110]。综上所述,乳源外泌体可以促进机体肠道发育,改善肠道菌群结构,发挥类似于益生元的功能,这为乳源外泌体作为一种新型益生元奠定了一定的基础。
5 小 结
目前,乳源外泌体的研究主要集中在乳房炎等疾病的诊断检测和作为益生元或药物载体应用方面。但是,因规模化提取纯化工艺等制约,乳源外泌体的临床应用仍然面临着很多挑战,仍有待进一步深入研究。
