鸡异质核糖核蛋白AB单克隆抗体的制备及鉴定
2022-04-23郑鹿平刘金玲柴书军杨艳艳
罗 俊,党 露,郑鹿平,刘金玲,柴书军,赵 东,杨艳艳,滕 蔓
(1. 河南省农业科学院 动物免疫学重点实验室/农业农村部动物免疫学重点实验室/河南省动物免疫学重点实验
室,河南 郑州 450002;2. 河南省农业科学院 中英禽病国际研究中心,河南 郑州 450002;3. 广州医科大学 附属第三医院,广东 广州 510095)
异质核糖核蛋白(hnRNPs)分属于一大类RNA结合蛋白(RBP)家族,目前共鉴定出19个hnRNP 基因,其编码的蛋白质大小介于30~120 ku。hnRNPs蛋白各家族成员间没有共有结构域,多样性的蛋白质结构决定了该家族蛋白质功能的多样性。该家族成员除了参与核酸代谢的多个方面,包括选择性剪接、mRNA 稳定、转录和翻译调节功能以外,还参与DNA 修复和端粒延长、染色质重塑和转录、mRNA 3′端加工、维持mRNA 稳定性以及促进或抑制肿瘤的发生和发展[1‑4]。异质核糖核蛋白AB(Heterogeneous nuclear ribonucleoprotein AB,hnRNPAB)是该蛋白质家族中一个特殊的成员,其直系同源物在N 端存在进化上保守的独特肽结构域[5‑7]。除与其他家族成员在细胞水平上调节基因表达的功能之外,hnRNPAB还在脊椎动物神经系统的发育过程中具有重要作用[8‑11],同时还参与了肿瘤的发生[12‑14]。
鸡hnRNPAB 蛋白包含有8 个转录变体和2 个同型蛋白质,其中p37 型含有353 个氨基酸残基,而p32型含有284个氨基酸残基[15]。前期研究发现,作为细胞癌基因miR-155 的同源物,马立克病毒(Marek’s disease virus,MDV)编码的miR-M4-5p 可特异性靶向识别并下调鸡hnRNPAB 蛋白的表达,从而促进原代鸡胚成纤维细胞(CEF)和永生化细胞系DF-1 的细胞增殖,同时还抑制细胞的凋亡,可能与MDV 的致瘤性有关[13]。目前,市售商品化抗hnRNPAB蛋白的抗体只能识别哺乳动物细胞,无法识别禽源hnRNPAB 蛋白,制约了开展进一步相关研究。鉴于此,通过原核表达系统制备鸡hnRNPAB蛋白,免疫小鼠后通过细胞融合技术筛选制备hnRNPAB 单克隆抗体,为开展hnRNPAB 蛋白相关功能研究奠定基础。
1 材料和方法
1.1 主要试剂和材料
pET-30a(+)载体、SP2/0细胞、HEK293T、BHK-21、Marc-145、MDBK 传代细胞系均为本实验室保存;E.coli克隆菌株DH5α 和表达菌株BL21、限制性内切酶XhoⅠ和EcoRⅠ、ExTaq酶、DNA Ligation Kit 2.0 和DNA Marker 等均购自TaKaRa 公司;异丙基-β-D-硫代半乳糖苷(IPTG)购自Solarbio 公司;Ni-NTA Fast Start Kit 购自Qiagen 公司;PEG-1500 购自德 国 Roche 公 司 ;NE-PERTMNuclear and Cytoplasmis Extraction Reagents 购自Thermo 公司;NcmECL Ultra(NCM)显影液和无血清细胞冻存液购自新赛美公司;FITC-羊抗鼠IgG 和HRP-羊抗鼠IgG 购自美国Abbkine公司;细胞培养基和胎牛血清(FBS)均购自Gibco公司。
1.2 鸡hnRNPAB基因表达载体的构建
用Primer Premier 5.0 软件设计扩增鸡hnRNPAB基因(GenBank 登录号:NM_205328)的特异 性 PCR 引 物 , 上 游 引 物 : 5′-GGAATTCGAAGGCGACCAGATCAACG-3′,下游引物: 5′-CCTCGAGTAAAGGCGAAAGGAGCTGC-3′(下划线标示限制性内切酶EcoRⅠ和XhoⅠ的酶切位点),常规方法提取CEF 总RNA 反转录为cDNA,以此为模板,PCR 扩增鸡hnRNPAB基因的部分片段。切胶回收经1%琼脂糖凝胶电泳分析纯化的PCR 产物,连接pMDTM19-T 载体并转化DH5α 感受态细胞,挑选阳性质粒测序鉴定。用EcoRⅠ和XhoⅠ双酶切测序正确的pMDTM19-T-hnRNPAB质粒,胶回收后用DNA Ligation Kit 2.0 连接酶连接获得pET-30a-hnRNPAB重组表达质粒,转化感受态细胞E.coliBL21,并涂含氨苄青霉素的LB 平板,过夜培养,将PCR 鉴定为阳性的单菌落扩大培养、提取质粒并进行双酶切鉴定。
1.3 鸡hnRNPAB重组蛋白的表达及纯化
用含100 mg/L 氨苄青霉素的LB 培养基活化含pET-30a-hnRNPAB重组表达质粒的菌种,置37 ℃摇床,以250 r/min 振摇培养菌液,至其OD600达到0.6~0.8 时加入IPTG,32 ℃继续诱导培养,收集菌体,离心并重悬后进行超声破碎,通过聚丙烯酰胺凝胶电泳(SDS-PAGE)分析裂解后的上清液和沉淀中蛋白质的表达情况和可溶性。将检测确定诱导表达的菌种按上述方法大量诱导表达,收集菌体按照Qiagen 公司的Ni-NTA Fast Start Kit 试剂盒说明书进行蛋白质纯化,测定纯化的蛋白质含量,通过SDS-PAGE电泳检测纯化结果。
1.4 Balb/c小鼠免疫
将纯化的hnRNPAB 蛋白与佐剂乳化后免疫6周龄Balb/c 雌性小鼠5 只(50 μg/只),背部皮下分点注射,其中弗氏完全佐剂用于首免,弗氏不完全佐剂用于此后的加强免疫,每次免疫间隔3周,共免疫4次,4 免后的第21 天断尾采血。以纯化的hnRNPAB 蛋白包被酶标板(10 mg/L),分别利用间接酶联免疫吸附试验(iELISA)和间接免疫荧光试验(IFA)来检测免疫小鼠血清效价。选择其中效价最高的小鼠,以2 倍免疫剂量且不加佐剂的抗原腹腔注射小鼠进行超免,3~5 d 后无菌取小鼠脾脏用于细胞融合。
1.5 杂交瘤细胞制备及筛选
采用常规方法进行细胞融合,SP2/0 细胞与超免后Balb/c 小鼠脾细胞按1׃10 的比例混合后,加PEG-1500 融 合 剂1 mL,用RPMI-1640/HAT/10%FBS培养液将融合后的细胞分散接种于6块96孔细胞板中,200 μL/孔,置37 ℃、5%CO2培养箱中培养10 d后取杂交瘤细胞上清进行检测。按照iELISA方法,以纯化的hnRNPAB 蛋白包被酶标板(10 mg/L),检测杂交瘤细胞上清,将结果为阳性的杂交瘤细胞转至24 孔细胞板扩大培养3~4 d,选择iELISA 复测强阳性孔进行有限稀释,一般2~3次,最后选出状态良好的单克隆杂交瘤细胞,扩大培养后离心收集细胞,用无血清细胞冻存液重悬后冻存于液氮中。
1.6 单克隆抗体制备
采用体内腹水诱生法生产单克隆抗体。将灭菌液体石蜡腹腔注射经产的Balb/c母鼠进行致敏预处理,0.5 mL/只。收集3×106~5×106个单克隆抗体杂交瘤细胞,无菌PBS洗2遍,最后重悬于0.5 mL无菌PBS 中,腹腔接种已致敏至少14 d 的Balb/c 经产母鼠。观察7~14 d,待小鼠腹部明显膨胀时无菌采集腹水,在5 000 r/min 条件下离心15 min,收集上清,-20 ℃保存备用。
1.7 单克隆抗体的Western blot鉴定
收集1×106个CEF,2 000 r/min 离心5 min,弃去上清,按照NE-PERTMNuclear and Cytoplasmis Extraction Reagents 试剂盒说明书提取CEF 核蛋白,-80 ℃保存备用。将提取的CEF 核蛋白进行SDS-PAGE 电泳,然后将凝胶电泳分离的蛋白质转移到PVDF 膜上,5%脱脂奶粉通过室温封闭PVDF膜2 h,PBST 洗涤3 遍;加待检的抗hnRNPAB 蛋白单克隆抗体,置4 ℃冰箱孵育过夜,PBST 充分振摇洗涤;加1%脱脂奶粉稀释的HRP标记的羊抗鼠IgG(1∶1 000),37 ℃孵育1 h,PBST 充分振摇洗涤。将NcmECL Ultra(NCM)显影液A、B(1∶1)混合后滴在膜上孵育1~3 min,置凝胶成像仪中观察结果。
1.8 单克隆抗体的IFA染色及间接免疫细胞化学试验(ICA)鉴定
分别用IFA 和ICA 对筛选的单克隆抗体与不同来源细胞蛋白质的反应性进行检测。取8~10 日龄SPF 鸡胚,常规方法制备CEF,同时从液氮中取出冻存的HEK293T、BHK-21、Marc-145、MDBK 和PK-15 传代细胞系进行复苏,分别接种到24 孔细胞板中,置37 ℃、5%CO2培养箱中培养24 h 后弃去生长液,加预冷的甲醇/丙酮(1∶1)200 μL/孔,室温固定15 min;弃 去 固 定 液,PBST 洗3 遍,加 入 待 检 的hnRNPAB 单克隆抗体,置37 ℃温箱孵育30 min;PBST 洗3 遍,加入FITC-羊抗鼠IgG 和HRP-羊抗鼠IgG(1∶500),37 ℃温箱孵育30 min;PBST 洗3 遍后加PBST(200 μL/孔),荧光显微镜下观察细胞染色结果。
2 结果与分析
2.1 鸡hnRNPAB蛋白表达载体的构建及鉴定
PCR 扩增鸡hnRNPAB部分基因片段,回收纯化PCR 扩增产物并连接pMDTM19-T 载体后转化DH5α感受态细胞,对PCR 鉴定为阳性的菌液提取质粒并进行测序分析,将符合预期的pMDTM19-ThnRNPAB质 粒 用EcoR Ⅰ和XhoⅠ双 酶 切,回 收hnRNPAB基因片段并连接转化E.coliBL21 感受态细胞,挑取单菌落,扩大培养后提取质粒,分别进行双酶切和PCR鉴定,可见1 317 bp大小的电泳条带,符合预期(图1)。测序分析结果也与预期完全相符,确认成功构建重组表达质粒pET-30ahnRNPAB。
2.2 鸡hnRNPAB蛋白的表达与纯化
用IPTG 诱导培养pET-30a-hnRNPAB转化的E.coliBL21 阳性菌液8 h 后,收集并超声破碎菌体,通过12% SDS-PAGE 电泳分析超声上清液和沉淀中蛋白质的表达情况。结果表明,鸡hnRNPAB 蛋白以可溶性形式表达。上清液中可溶性表达的目的蛋白质用树脂Ni 进行纯化,收集洗脱液经SDSPAGE 电泳分析,结果表明,纯化后可获得纯度较高的hnRNPAB蛋白(图2)。

图2 鸡hnRNPAB蛋白的诱导表达及纯化蛋白的SDSPAGE分析Fig.2 SDS-PAGE analysis of expressed and purified chicken hnRNPAB protein
2.3 抗鸡hnRNPAB蛋白的单克隆抗体制备
用纯化后的hnRNPAB 重组表达蛋白4 次免疫Balb/c 小鼠,最后一次免疫第21 天,iELISA 检测免疫小鼠血清效价。结果显示,5 只小鼠均产生较高效价的抗体,其中2 号小鼠血清iELISA 效价最高,达到了1∶32 000(表1)。IFA 检测结果也显示,抗体荧光染色主要集中在CEF 细胞核内(图3)。故选择2 号小鼠进行超免,无菌摘取超免小鼠脾脏,与SP2/0 细胞(10∶1)混合后进行细胞融合,9 d 后取杂交瘤细胞上清,利用iELISA 试验筛选效价最高的杂交瘤细胞株,进行3 轮亚克隆后获得1 株稳定分泌抗鸡hnRNPAB 蛋白的单克隆抗体杂交瘤细胞株,命名为5H5-H1。收集5H5-H1 杂交瘤细胞接种石蜡致敏的经产母鼠,成功制备了抗鸡hnRNPAB 蛋白的单克隆抗体腹水。

表1 免疫小鼠血清iELISA效价Tab.1 The iELISA titers of serum from immunized mice
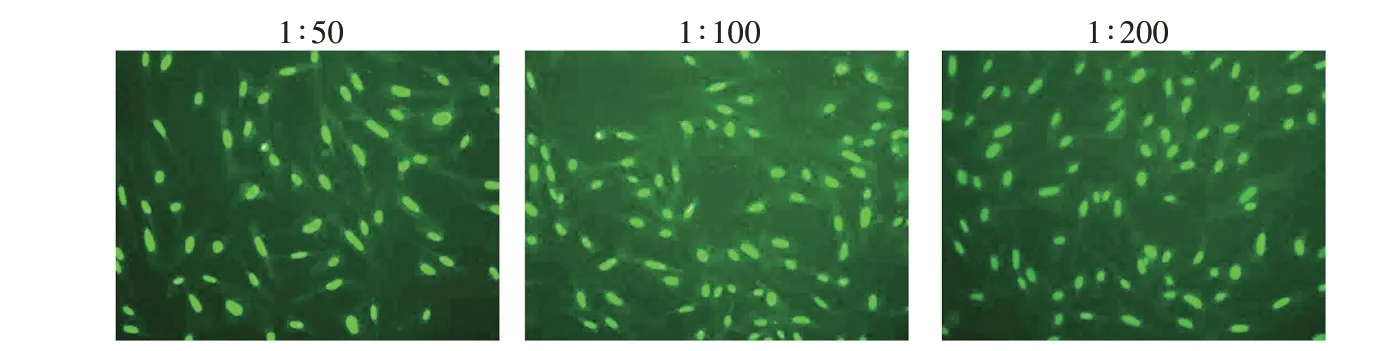
图3 hnRNPAB免疫小鼠血清与CEF反应的IFA染色(100×)Fig.3 IFA staining of CEF cells using serum from hnRNPAB-immunized mice(100×)
2.4 抗鸡hnRNPAB单克隆抗体的Western blot鉴定
提取CEF 核蛋白,经SDS-PAGE 电泳分离后转膜封闭,然后依次孵育5H5-H1 单克隆抗体腹水(1∶4 000)和HRP标记的羊抗鼠IgG(1∶1 000),进行Western blot 鉴定。结果显示,单克隆抗体5H5-H1特异性识别鸡hnRNPAB 蛋白,出现2条特异性反应条带,分别约为37 ku 和32 ku,与预期的鸡hnRNPAB蛋白大小相符(图4)。
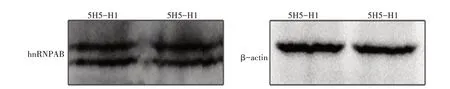
图4 Western blot分析5H5-H1单克隆抗体与鸡hnRNPAB蛋白的反应性Fig.4 Specific reaction of mAb 5H5-H1 to chicken hnRNPAB protein determined by Western blot analysis
2.5 hnRNPAB单克隆抗体与多种细胞蛋白质反应的IFA和ICA染色鉴定
待原代CEF 和传代细胞HEK293T、BHK-21、Marc-145、MDBK、PK-15 长至稀疏单层,固定细胞,分别孵育5H5-H1 单克隆抗体腹水(1∶4 000),然后再分别孵育FITC-羊抗鼠IgG 和HRP-羊抗鼠IgG,鉴定5H5-H1 单克隆抗体与不同细胞的反应性。IFA 和ICA 结果显示,5H5-H1单克隆抗体与CEF 及5 种哺乳动物细胞均发生特异性反应,并且这种特异性染色都是集中在细胞核(图5)。
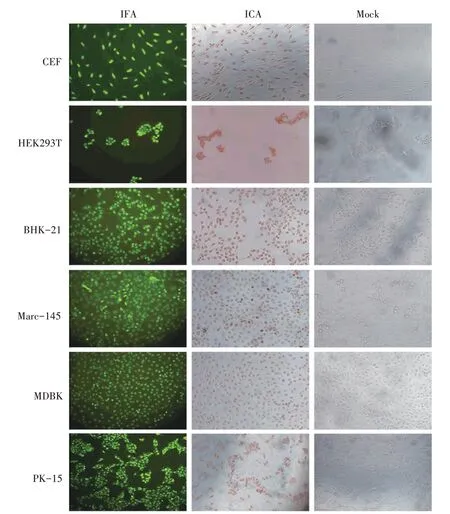
图5 IFA和ICA染色检测5H5-H1单克隆抗体与6种不同细胞的反应性(100×)Fig.5 Reaction of mAb 5H5-H1 to six cell lines determined by IFA and ICA(100×)
3 结论与讨论
哺乳动物的hnRNPAB 蛋白氨基酸同源性很高,基本都在92%以上,而鸡的hnRNPAB 蛋白氨基酸序列与哺乳动物hnRNPAB 蛋白氨基酸序列同源性仅为83.3%~86.2%。但不同物种的hnRNPAB 蛋白仍具有一些共同结构,如RNA 识别基序(RNA recognition motif,RRM)、Gly-Rich 基序和其他辅助区域[2]。笔者所在课题组长期致力于家禽肿瘤病的相关研究,前期研究发现,MDV 编码的miR-M4-5p可靶向下调鸡hnRNPAB基因的表达,进而调控细胞增殖,与MDV 肿瘤发生有关[13]。为开展相关功能研究及信号通路分析,曾先后购买多种进口或国产的商业化抗hnRNPAB 蛋白的抗体。然而,这些抗体与大部分哺乳动物细胞的hnRNPAB 蛋白反应性良好,但均与禽类细胞hnRNPAB 蛋白不发生反应,这很可能是由于禽类与哺乳动物hnRNPAB 蛋白氨基酸同源性不高所致。
为了制备能够与禽源细胞hnRNPAB 蛋白反应的单克隆抗体,本研究首先对鸡hnRNPAB 蛋白与哺乳类hnRNPAB 蛋白氨基酸进行了同源性序列比对,进而对鸡hnRNPAB 蛋白抗原性和疏水性进行分析,去掉鸡hnRNPAB蛋白N末端相对疏水且含有较少潜在抗原的区域,选取鸡hnRNPAB 蛋白与哺乳类hnRNPAB 蛋白比较保守的抗原性区域,进行引物设计,从CEF 中PCR 扩增了鸡hnRNPAB基因,成功构建了pET-30a-hnRNPAB原核表达载体,转化E.coliBL21感受态细胞,经IPTG诱导后获得了可溶性表达的hnRNPAB 蛋白,以纯化后的可溶性重组蛋白多次免疫Balb/c 小鼠后进行细胞融合,多种方法筛选最终获得了能够稳定分泌抗鸡hnRNPAB蛋白的特异性单克隆抗体杂交瘤细胞株5H5-H1,并制备了相应的单克隆抗体腹水。IFA 和Western blot鉴定结果表明,5H5-H1单克隆抗体与鸡CEF细胞核内表达的hnRNPAB 蛋白产生了很强的特异性反应。进一步的IFA 和ICA 分析表明,该单克隆抗体与其他来源于人、鼠、猴、牛和猪的多种哺乳动物细胞系表达的hnRNPAB 蛋白反应特异性均良好。综上,本研究制备的hnRNPAB 单克隆抗体,不仅识别鸡源细胞表达的相应蛋白质,而且可以与多种哺乳动物细胞表达的hnRNPAB 蛋白产生特异性反应,可同时满足禽类和哺乳类hnRNPAB 蛋白相关的功能研究的需要,具有比较重要的科学价值。
