黑龙江省水稻种质稻瘟病抗性评价及抗瘟基因结构分析
2022-04-21张亚玲高清赵羽涵刘瑞付忠举李雪孙宇佳靳学慧
张亚玲,高清,赵羽涵,刘瑞,付忠举,李雪,孙宇佳,靳学慧
黑龙江省水稻种质稻瘟病抗性评价及抗瘟基因结构分析
张亚玲,高清,赵羽涵,刘瑞,付忠举,李雪,孙宇佳,靳学慧
黑龙江八一农垦大学农学院/黑龙江省植物抗性研究中心,黑龙江大庆 163000
【】稻瘟病严重威胁黑龙江省水稻的生产,选育和利用抗瘟品种是最经济、安全和有效的措施。了解黑龙江省水稻主栽品种的稻瘟病抗性,明确稻瘟病抗性基因的有效性,为黑龙江省稻瘟病抗病种质资源的选育和利用提供依据。2018年秋季在黑龙江省水稻主产区采集134株水稻单孢菌株,采用病原物接种鉴定方法对黑龙江省50个水稻主栽品种进行抗性分析;针对已报道的35个稻瘟病抗病基因,利用基因特异性引物对参试品种及阳性对照品种进行抗瘟基因检测,对部分无阳性对照品种抗性基因扩增后测序,与NCBI中公布的参考序列比对,分析稻瘟病抗性基因在相应品种中的存在情况;通过基因聚合类型与品种抗性表现相关性分析,明确与黑龙江省水稻品种抗性表现相关的基因型。根据抗性频率评价,参试的黑龙江省50份水稻品种中,龙粳20表现为抗,龙粳67、龙垦202、龙粳40、龙粳31、龙粳57和龙粳43表现为中抗,其余43个品种均表现感病。通过品种组合抗性分析发现龙粳20与龙粳67等33对品种组合的联合抗性系数(resistance association coefficients,RAC)较高,联合致病性系数(virulence association coefficients,VAC)较低,联合抗病性较好,该类搭配结构具有良好的应用前景。通过特异性引物对品种携带的抗瘟基因鉴定结果显示,抗瘟基因、、和在供试品种中均被检测到,、、、、、、、、、、、和的被检出率为50%—100%,说明这类基因在黑龙江省水稻育种中应用较广泛;、、、、、和的被检出率为10%—50%,仅在2个品种中被检测到,仅在吉粳88中被检测到,而、、、、-、、、和在供试品种中均未被检测到,说明这类基因在黑龙江省水稻品种中分布较少。通过品种基因型分析发现,供试品种携带抗瘟基因12—19个不等,共58种抗瘟基因型,说明供试品种的抗瘟基因组合类型较丰富。单基因及多基因聚合与抗病相关性分析结果显示,、、、和的分布频率与携带该基因且表现抗病的百分率相当;研究发现品种携带抗瘟基因越多,其表现为抗病品种的概率越高,携带+++聚合类型的6份品种均表现为抗病。参试的黑龙江省水稻种质资源抗性偏低,部分品种组合种植有较高应用价值,抗瘟基因在参试品种中分布不等,、、、和在品种抗病方面起主导作用,+++基因聚合类型可以提高水稻抗瘟性。
水稻;稻瘟病;抗性频率;抗瘟性评价;抗瘟基因
0 引言
【研究意义】水稻(L.)是当今世界主要粮食作物之一,稻瘟病是由子囊菌(无性态:)[1]引起的一种常见的水稻真菌病害,对水稻粮食生产造成巨大威胁。培育和种植抗稻瘟病品种是防治稻瘟病最经济有效、绿色环保的途径,但由于稻瘟病菌致病型分化速度快、变异频繁,经常导致某一抗病品种在大面积应用3—5年后便抗性“丧失”,造成严重经济损失[2]。抗瘟品种的合理利用、抗性基因的合理布局以及基因的聚合是减缓抗性“丧失”的主要途径之一[3]。因此,深入研究水稻品种的抗性表现及稻瘟病抗性基因的有效性,对抗瘟选育与品种的合理利用具有重要意义。【前人研究进展】品种抗瘟性评价方法多采用菌株孢子悬浮液喷雾接种法,接种后,人为控制温度、湿度等有利于侵染和发病的条件,能准确反映品种对病菌的真实抗性。田大刚[4]对福建省稻瘟病重发区进行了连续3年的苗圃水稻抗性鉴定试验,从1 092份材料中筛选到高抗、抗病及中抗材料共344份;阎勇等[5]通过人工接种的方法进行水稻抗性鉴定,从52份华南区常用籼稻亲本中筛选到4份广谱抗源及抗瘟性较好的常规稻恢复系和保持系。近年来,黑龙江省水稻抗瘟性鉴定工作取得一系列进展,王桂玲等[6]通过人工接种鉴定274份黑龙江省水稻区试材料,筛选出26份抗性较好的品种;郭丽颖等[7]通过注射接种对36份黑龙江省水稻品种进行抗瘟性鉴定,筛选出稻花香等10个抗性较好的品种,以及稻花香+龙粳31等22个防病效果较好的组合;张亚玲等[8]通过离体接种鉴定黑龙江省20份水稻品种的抗瘟性,筛选出松粳10号等抗性较好的品种,分析了品种组合的利用价值。水稻稻瘟病抗病基因(resistant genes)决定其质量抗性,以不亲和的互作方式存在,迄今已陆续从不同水稻品种中克隆了39个抗瘟基因。近年来,不同品种的抗稻瘟病基因检测研究成果层出不穷,韩笑等[9]、王亚等[10]分别对上海市和河南省主栽品种进行抗瘟基因检测,明确了当地水稻品种中抗瘟基因的存在情况。黑龙江省水稻抗瘟基因检测也不断取得新进展,辛威[11]利用功能性分子标记对225份水稻种质资源进行8个抗瘟基因的检测;周弋力[12]对34份黑龙江省水稻主栽品种进行部分抗瘟基因分子标记检测,以上研究明确部分抗性基因在参试品种中的分布。基因对基因学说及加性效应表示多个抗瘟基因聚合到同一品种中可在一定程度上降低其抗病性下降或丧失的概率。目前,有效基因在一个品种中聚合是快速寻找抗病品种的有效方法,同时也是一种遗传改良方法。相亚超等[13]为了明确黑龙江主栽水稻品种及骨干育种亲本中抗稻瘟病基因的分布情况和利用价值,对102份水稻资源中的7个抗稻瘟病基因进行了分子检测,并对它们的分布情况和抗病效应进行相关性分析,发现、、和对黑龙江水稻稻瘟病抗性贡献较大,当和聚合时,对黑龙江省水稻抗瘟性改良最大。【本研究切入点】黑龙江省是中国重要粳稻产区,其地处高纬地区,温度、光照等自然气候条件有限,水稻种质资源相对匮乏单调,一些抗病种质资源重复利用,使得抗病品种结构简单化,且缺少抗谱广、抗性持久的品种,稻瘟病持续连年发生,严重胁迫水稻生产。选育和合理利用抗病品种是防控稻瘟病最经济有效且环境友好的策略。由于稻瘟病菌生理小种的复杂多样、高度变异等特点,导致优良水稻抗瘟品种的抗瘟性逐渐下降甚至丧失。前人研究大多采用接种病原物的方法进行品种抗瘟性评价,或只对品种抗瘟基因进行鉴定,未将两方面结合来分析品种或抗瘟基因的利用方向。因此,进行水稻品种的抗瘟性评价以及抗瘟基因组成,对稻瘟病防治和抗病育种具有重要意义。【拟解决的关键问题】本研究通过黑龙江省水稻种质资源对水稻主产区稻瘟病菌的抗性评价及其携带的抗瘟基因进行检测分析,旨在了解供试种质的抗瘟性,评估品种及品种组合的应用价值,为黑龙江省筛选优质水稻抗病材料、挖掘新的抗瘟基因资源、合理布局以及稻瘟病防控提供重要理论依据。
1 材料与方法
试验于2019年6月至2021年1月在黑龙江八一农垦大学植物抗性研究中心实验室完成。
1.1 供试水稻品种
用于抗瘟性评价的50份黑龙江省水稻材料包括36个粳型常规稻品种(龙粳21、龙粳62、龙粳20、龙粳26、龙粳29、龙粳31、龙粳40、龙粳43、龙粳46、龙粳50、龙粳51、龙粳52、龙粳57、龙粳58、龙粳59、龙粳63、龙粳65、龙粳66、龙粳47、龙粳67、垦稻30、垦稻34、垦稻31、垦稻41、龙垦201、龙垦202、龙垦227、龙垦257、绥粳18、绥粳23、龙糯3号、北稻6号、龙洋11、稼禾1号、中龙香粳1号和龙庆稻5号),10个黑龙江省水稻品种(龙广优13、稻花香7号、富粳188、合1045、粳花香、哈香稻3号、普粳稻、润香稻、水粳香和鸿源5290)和4个优质种质资源(小粒香、沙沙尼、谷润田香和谷润田香2号)。对照材料为蒙古稻,易感病。
用于抗瘟基因鉴定的供试水稻品种:50份用于抗瘟性评价的供试水稻品种,17份阳性对照材料分别为IRBLt-K59()、IRBLsh-S()、IRBLb-B()、IRBLz5-CA()、IRBL9-W()、IRBLzt-T()、Digu()、IRBL5-M()、IRBLi-F5()、IRBLa-A()、CO39()、IRBLk-Ka()、IRBL1-CL()、IRBLkh-K3()、IRBLkm-Ts()、IRBLkp-K60()和IRBLta-K1(),1份阴性对照材料为丽江新团黑谷(LTH)。以上材料均由黑龙江八一农垦大学植物抗性研究中心提供。
1.2 参试稻瘟病菌株
2018年于黑龙江省4个积温带主要稻作区采集的穗颈瘟标样,经单孢分离获得134株稻瘟病菌株,采用滤纸片保存法保存备用[14]。
1.3 水稻品种抗瘟性评价
采用室内水培法培养水稻幼苗:挑选饱满的种子,用25%的氰烯菌酯3 000倍溶液室温浸种48 h,清水冲洗3次放入培养皿内,用滤纸保湿,于28℃恒温培养箱内催芽48 h,播种于固定在塑料水培盒(内尺寸:225 mm×155 mm×55 mm)内的定植篮(上端内径34 mm,下端内径28 mm,高45 mm)中,每个定植篮播10粒种子,置于水培架上育苗,清水培养3 d后,换霍格兰(Hoagland)营养液[15]培养,每3天更换一次营养液。参考孟峰等[16]方法进行菌株活化及产孢培养和接种。接种后将其移入25℃的保湿棚内进行遮光保湿培养24 h,然后返回室温自然正常培养,接种后7 d,参照靳学慧等[17]方法进行调查,按照叶瘟9级分类标准,0—3级表现记录为抗病(resistance,R),4—9级记录为感病(susceptible,S)记载发病情况。采用阮宏椿等[18]方法对水稻品种抗性进行评价,其评价指标用抗性频率(resistance frequency,RF)来评价水稻品种抗性表现。当RF≥80%为抗病(R),60%≤RF<80%为中度抗病(medium resistance,MR),40%≤RF<60%为中度感病(medium susceptible,MS),RF<40%为感病(S);运用联合抗病性系数(resistance association coefficients,RAC)和联合毒力系数(virulence association coefficients,VAC)分析品种搭配的抗病效果[19],以品种组合既高抗稻瘟病菌又被稻瘟病菌侵染概率小为布局优劣评价标准。相关公式如下:

式中,R1、R2指对品种1和品种2致病性均弱;S1、S2指对品种1和品种2致病性均强。
1.4 参试品种主要抗病基因的鉴定
水稻生长至四叶期,选取新鲜叶片,利用OMEGA植物基因组提取试剂盒(HP Plant DNA Kit)提取水稻基因组DNA。选取已报道的抗稻瘟病基因特异性引物(电子附表1),由生工生物工程(上海)股份有限公司合成。
PCR反应体系为10×Buffer(Mg2+)2.0 µL、dNTP Mixture 1.6 µL、正反向引物(10 µmol·L-1)各0.3 µL、rTaq酶(TaKaRa)0.1 µL、DNA模板(50 ng·µL-1)1 µL,加ddH2O补至20 µL。PCR扩增程序为94℃5 min;94℃45 s,退火45 s(具体温度据特异性引物的GC含量设定),72℃延伸(具体时间据扩增片段大小确定),35个循环;72℃10 min;4℃保存待检测。酶切反应体系:限制性核酸内切酶0.5 µL、PCR产物8 µL和10×Buffer 1.5 µL,37℃水浴2—4 h。
采用1.5%琼脂糖凝胶电泳和8%聚丙烯酰胺凝胶电泳检测。利用BIO-RAD凝胶成像系统拍照,Image Lab系统中的条带检测功能,参照Marker读取扩增条带。每材料试验重复3次。
1.5 部分抗瘟基因PCR产物测序分析
将无阳性对照且扩增出目标条带基因(、、、、、、、、、、、、、和)的扩增产物送生工生物工程(上海)股份有限公司测序及拼接,利用BioEdit软件查看峰图确认结果,从NCBI(https:// www.ncbi.nlm.nih.gov/)及参考文献中获取基因的参考序列,运用DNAMAN软件进行多序列比对。
1.6 数据处理与统计分析
运用Microsoft Excel 2016进行数据统计分析与作图,相关公式如下:

2 结果
2.1 参试水稻品种抗瘟性评价
利用分离的134株稻瘟病菌株对50份水稻品种进行苗期接种鉴定(表1)。表现中度(MR)抗病的品种占参试品种的12%,抗瘟评价为抗病(R)的仅有龙粳20,其抗性频率最高,为82.84%,占供试品种的2%;抗瘟评价为中度抗病(MR)的有龙粳67等6个品种,其他品种均为感病品种,其中,中度感病(MS)的有粳花香等8个品种,占供试品种的16%,感病(S)的有龙广优13等35个品种,占供试品种的70%,说明参试品种大部分为易感病品种。
2.2 参试水稻品种的联合抗瘟性分析
根据50份水稻品种的抗性频率,选出龙粳20等15份抗病性较好的品种,进行联合抗病分析。RAC值越大,2个品种均表现抗病的概率越大,VAC值越大,感病表现的一致性也就越大。从表2可以看出,RAC值最高为0.70(龙粳20与龙粳67),最低为0.13(龙粳57与龙粳62),VAC值最高为0.45(中龙香粳1号与水粳香),最低为0.08(龙粳57与龙粳20、垦稻34与龙粳20)。不同品种组合搭配的抗瘟效果差异较大,龙粳20与龙粳67等33对组合的RAC值较高,同时VAC值较低,占组合总数的31.43%;龙粳40与龙粳31等14对组合的RAC值与VAC值均较高,占组合总数的13.33%;龙粳67与稻花香7号等11对组合的RAC值与VAC值均较低,占组合总数的10.48%;中龙香粳1号与垦稻41等47对组合的RAC值较低,VAC值较高,占组合总数的44.76%。
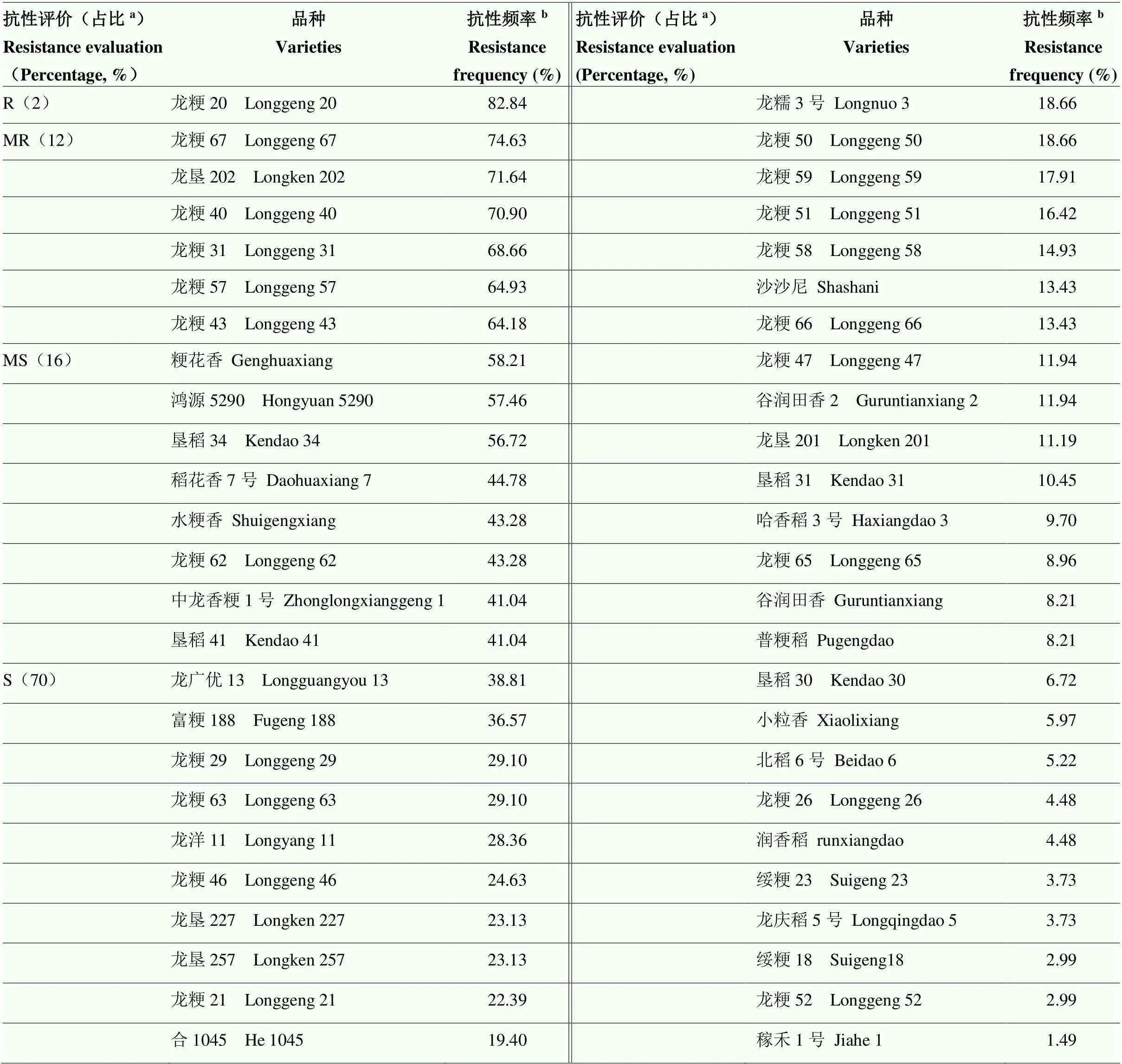
表1 供试水稻品种抗性频率及抗性评价
R:抗病;MR:中抗;MS:中感;S:感病。a:抗性评价占比(%)=(表现为该抗病级别的品种数量/参试品种总数量)×100%;b:抗性频率(%)=(对该品种表现抗病的菌株数量/参试总菌株数量)×100%。下同
R: Resistance; MR: Moderate resistance; MS: Moderate susceptible; S: Susceptible.a: The proportion of resistance evaluation (%)=(The number of varieties of this disease resistance /Total number of test varieties)×100%;b: Resistance frequency (%)=(Number of strains exhibiting disease resistance to the varieties/Total number of strains were tested)×100%. The same as below
2.3 参试水稻品种抗瘟基因的鉴定与分析
2.3.1 抗瘟基因特异性分子标记扩增结果 利用抗瘟基因对应的特异性分子标记对67份参试水稻品种、部分阳性和阴性对照品种的35个稻瘟病抗病基因进行PCR鉴定(图1和图2),以能够扩增出表1中抗病基因(R)对应的片段且不能扩增出等位感病基因(S)片段,表明该品种携带目标抗瘟基因,对于、、利用其引物能扩增出目标片段且能被对应的内切酶酶切,表明携带抗病基因。利用的特异性引物未扩增出任何条带,利用、的特异性引物未扩增出目标抗病条带,利用的特异性引物扩增出的条带不能被cⅡ酶切,表明供试品种中均不携带这4个基因。
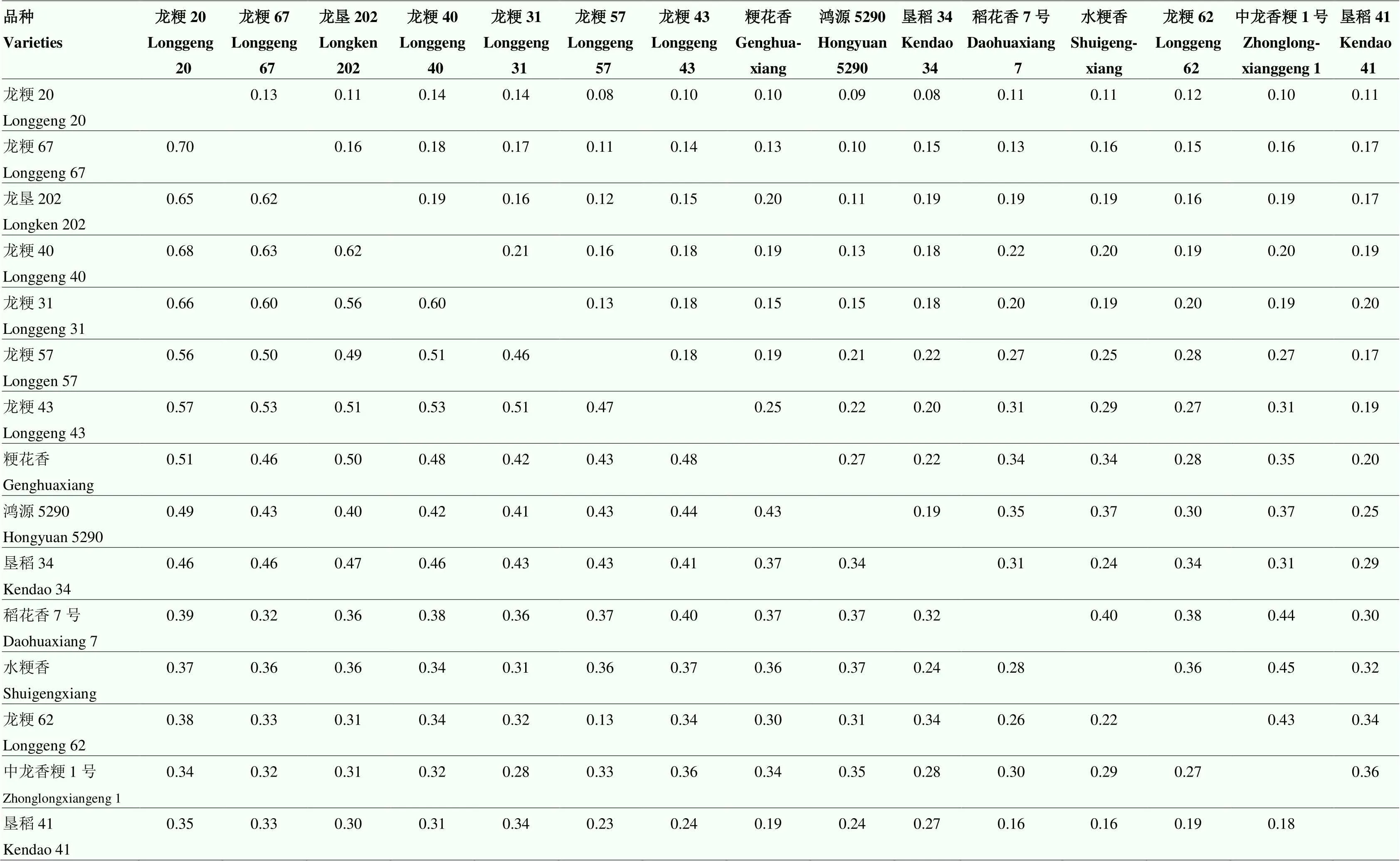
表2 部分水稻品种的联合抗病性系数和联合毒力系数
对角线左下部分为联合抗病性系数(RAC),右上部分为联合毒力系数(VAC)
The lower left part of the diagonal is the resistance association coefficient (RAC), the upper right part is the virulence association coefficient (VAC)

A:Pit;B:Pib;C:Pi2;D:Pi9;E:Piz-t;F:Pi5;G:Pi54;H:Pik;I:Pi1;J:Pikh;K:Pikm;L:Pita;M:Pish;N:Pi33;O:Pia;P:Pi-CO39;Q:Pikp;R:Pid2;S:Pid3;T:Pii。M:DL2000 DNA Marker,+:各基因的阳性对照材料,-:阴性对照材料丽江新团黑谷(LTH)或ddH2O,a:阳性对照材料PCR产物,b:阳性对照材料酶切产物,c:LTH PCR产物,d:LTH酶切产物。1:龙粳21,2:龙粳62,3:龙糯3号,4:垦稻30,5:垦稻34,6:绥粳18,7:绥粳23,8:小粒香,9:北稻6号,10:沙沙尼
2.3.2 部分抗瘟基因序列分析 无对照的抗瘟基因扩增出目标条带,将其PCR产物测序,与参考序列比对显示,的条带测序结果均与感病等位基因一致(221 bp)(图3-A);条带测序结果均与感病品种日本晴的序列(AL772419.4)一致,启动子功能位点碱基为A而非G,代表供试品种均携带其感病等位基因而非抗病基因(图3-B);其余扩增的抗瘟基因、、、、、、、和目标条带测序结果与参考序列一致(图4)。
2.4 抗瘟基因在供试品种中的分布
利用特异性引物鉴定35个抗瘟基因在供试品种中的分布,结果显示,抗瘟基因、、和在供试品种中均被检测到,、、、、、、、、、、、和的检出率为50%—100%,、、、、、和的检出率为10%—50%,仅在2个品种中被检测到,仅在1个品种中被检测到,而、、、、、、、和在供试品种中均未检测到(图5)。供试品种携带抗瘟基因12—19个不等,共58种抗瘟基因组合类型,携带15个抗瘟基因的品种最多,有19个,占28.36%,共17种基因组合类型,其次携带17个抗瘟基因的品种有13个,占19.40%,共12种基因组合类型,携带19个抗瘟基因的品种最少,仅有2个,占2.99%,共2种基因组合类型(表3)。结果表明,供试品种抗瘟基因组合类型比较丰富。
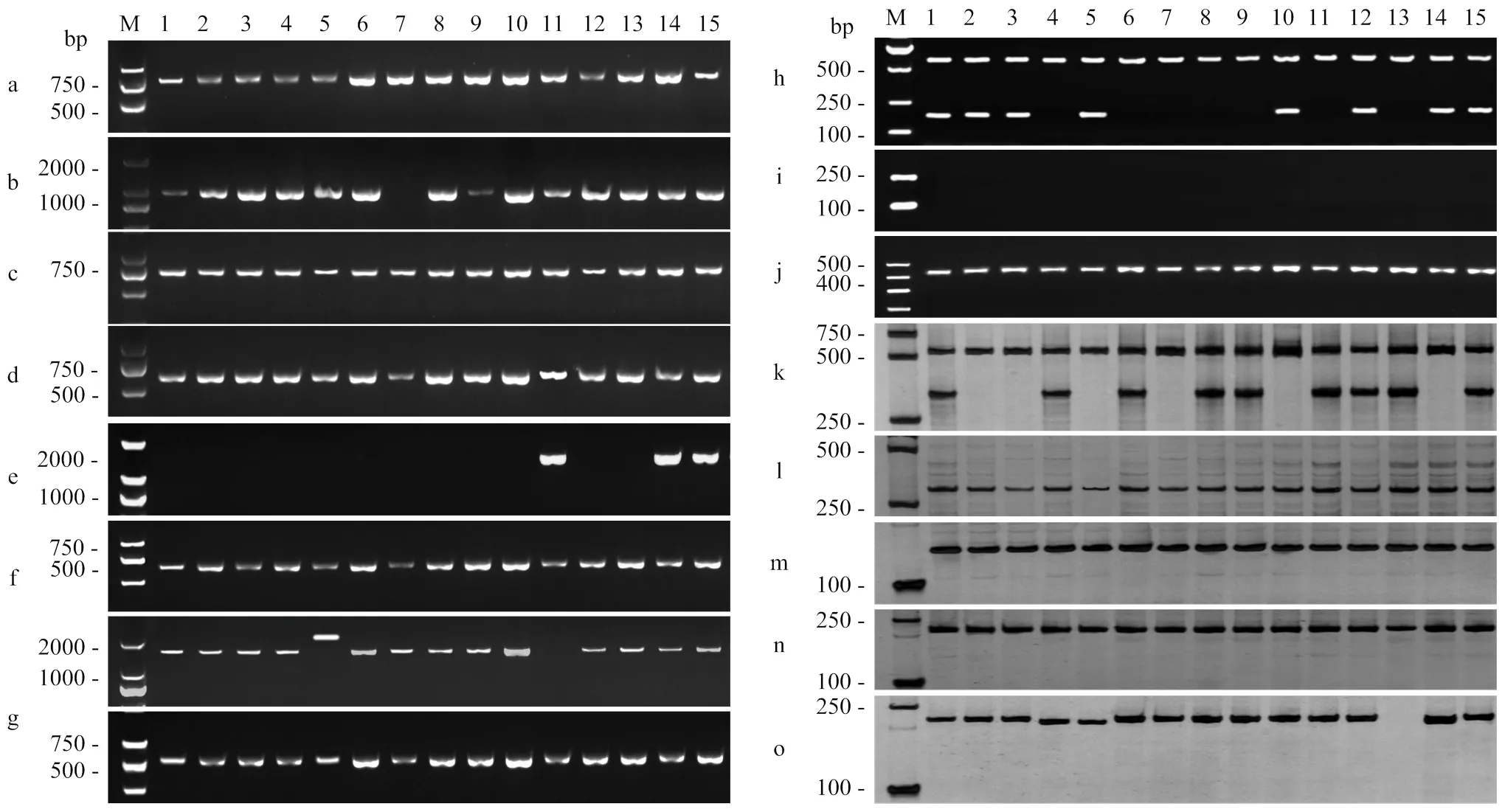
a:Pi37;b:Pi64;c:Bsr-d1;d:Pi63;e:Pi50;f:Pi36;g:Pi56;h:Pike;i:Pb1;j:Pi25;k:Pi35;l:pi21;m:Pigm;n:Pid3-A4;o:Ptr;M:DL500/2000 DNA Marker;1:龙粳21,2:龙粳62,3:龙糯3号,4:垦稻30,5:垦稻34,6:绥粳18,7:绥粳23,8:小粒香,9:北稻6号,10:沙沙尼,11:龙粳20,12:龙粳26,13:龙粳29,14:龙粳31,15:龙粳40
2.5 单基因及多基因聚合与抗病相关性分析
通过对50份水稻品种携带的抗瘟基因与抗病表现进行关联性分析,结果显示,大部分抗瘟基因的分布频率远高于携带该基因且表现抗病的百分率,说明携带这类基因的水稻品种既可能表现为抗病也可能表现为感病,而、、、和的分布频率与携带该基因且表现抗病的百分率相差不大,说明携带该类基因的水稻品种有较高概率表现抗病(图6)。
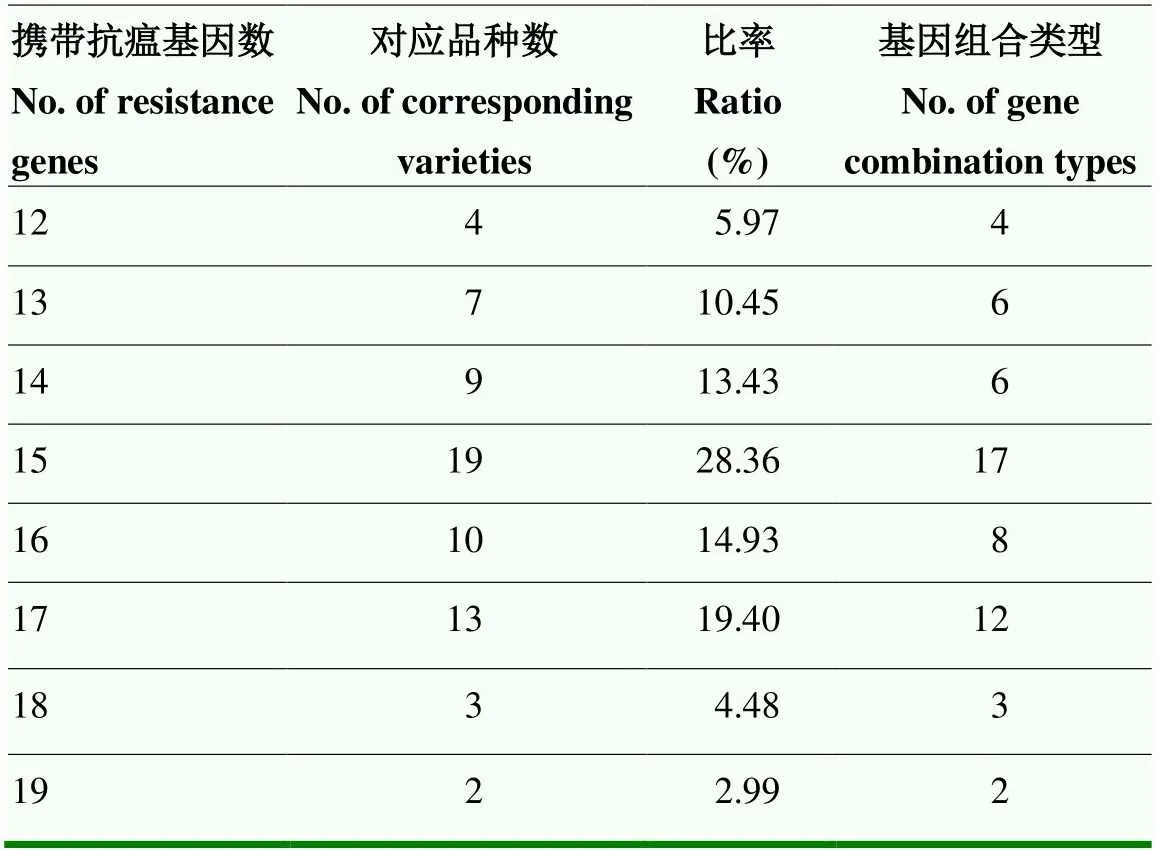
表3 供试品种携带抗瘟基因数目统计

A:Pi37;B:Pi64;C:Pi63;D:Pi50;E:Pi36;F:Pi35;G:pi21;H:Pigm;I:Ptr
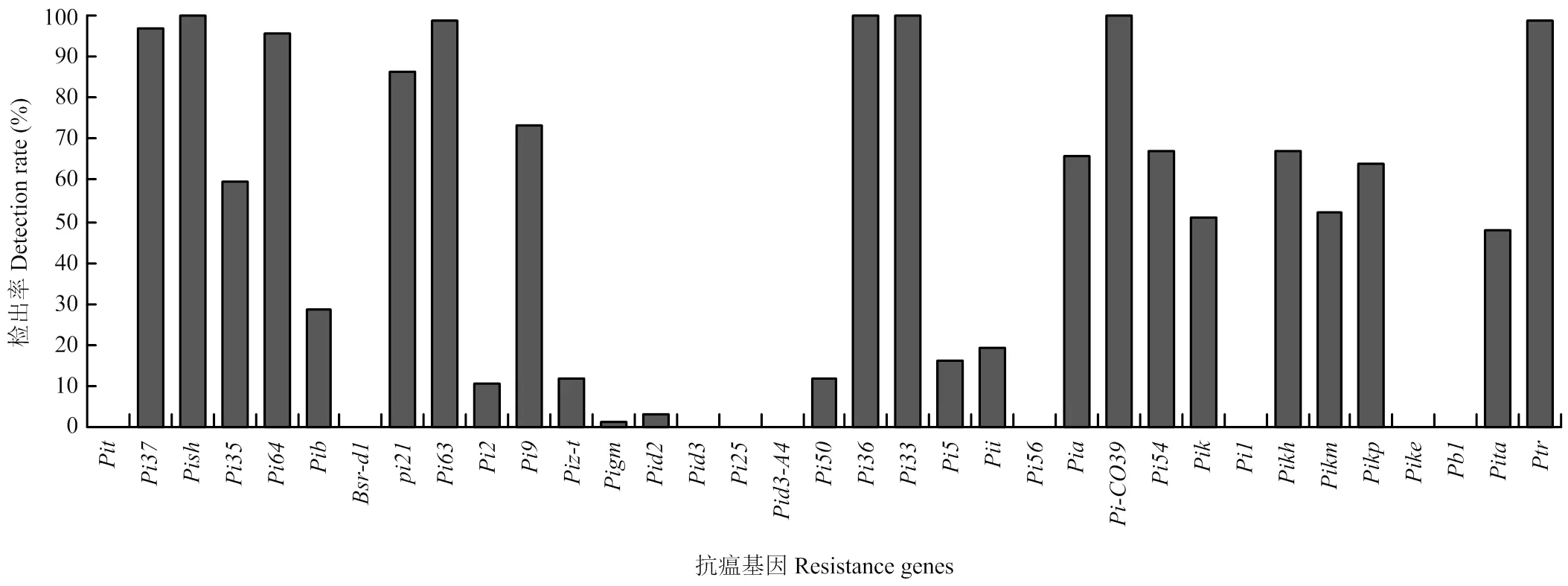
图5 抗瘟基因的检出率
结合50份供试品种的抗性表现,进一步分析不同抗瘟基因聚合对水稻抗病性的贡献(表4),50份供试水稻品种携带抗瘟基因12—19个不等,共有43种抗瘟基因型,基因型的水稻材料最多(3份),分别为绥粳18、龙粳63和润香稻,且研究发现携带相同抗瘟基因型的品种,抗性表现基本一致,表明抗瘟基因型是决定品种抗病程度的关键性因素。携带16个抗瘟基因的5份品种中有1份表现抗病,携带17个抗瘟基因的10份品种中有3份表现抗病,携带18个抗瘟基因的3份品种中有1份表现抗病,携带19个抗瘟基因的2份品种均表现抗病,而携带12—15个抗瘟基因的品种均表现感病,推断品种所携带抗瘟基因数量与其抗病性存在着一定程度的正相关作用,且携带十几个抗瘟基因仍表现感病的品种,可能是由于某些抗病基因之间存在拮抗作用反而降低了水稻抗病性,仍需进一步研究验证。携带+++(代表另外推测的基因或基因组合)聚合类型的6份品种均抗病,推测该基因型与其他抗瘟基因聚合或与未知新抗病基因互作可能对抗病性起正向调节作用,对水稻抗病性有一定的贡献,可为进一步挖掘广谱抗病新基因提供参考。
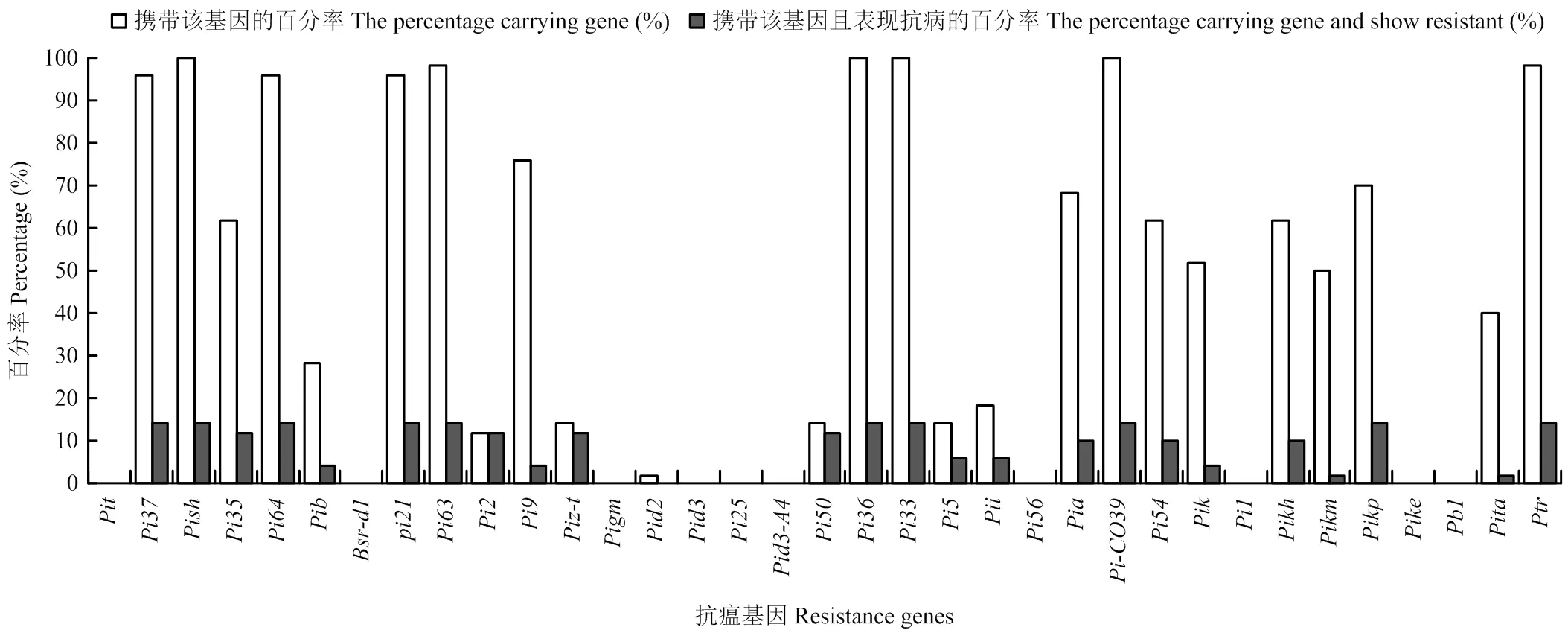
图6 抗瘟基因对抗病表型贡献情况
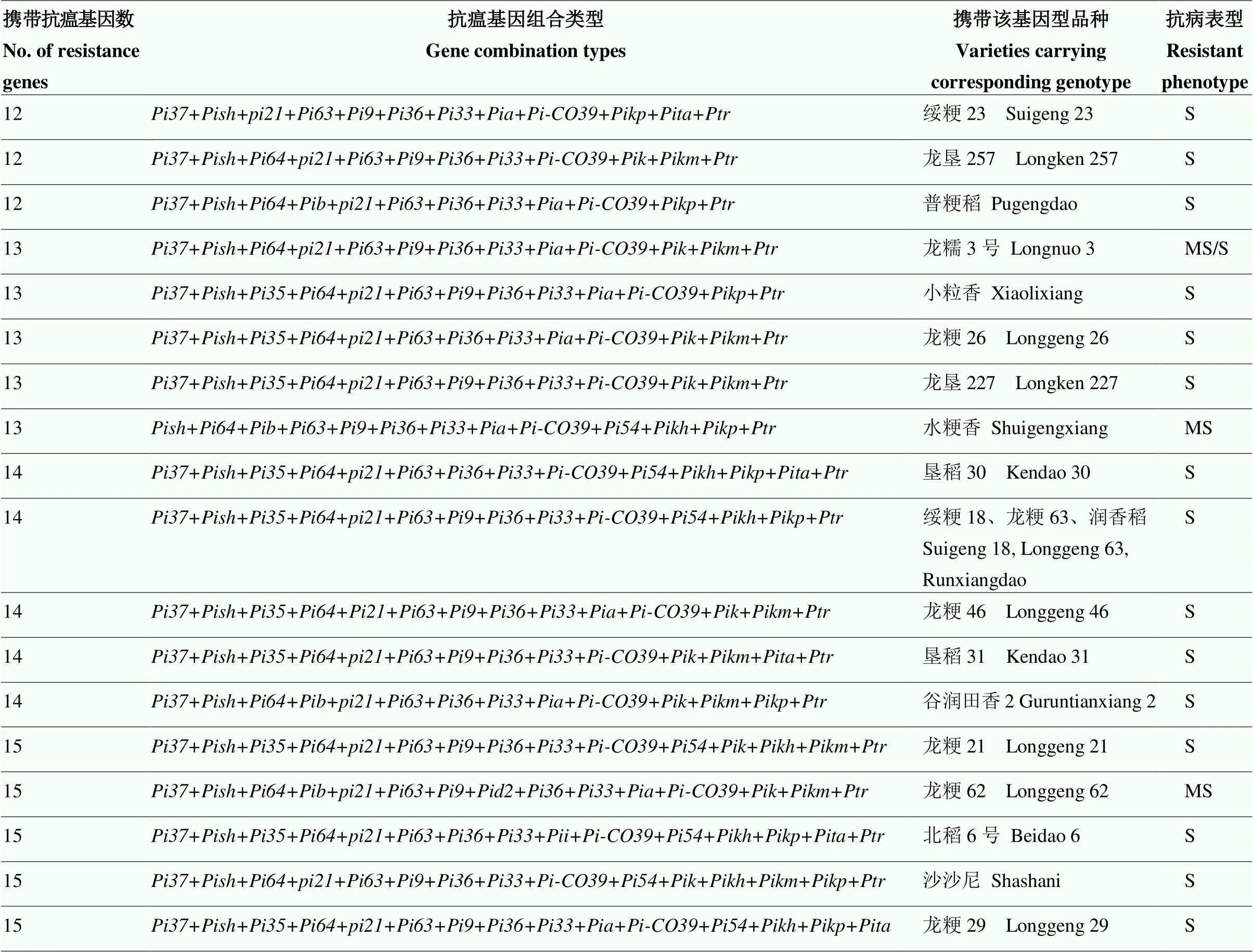
表4 不同抗瘟基因型的抗病表现
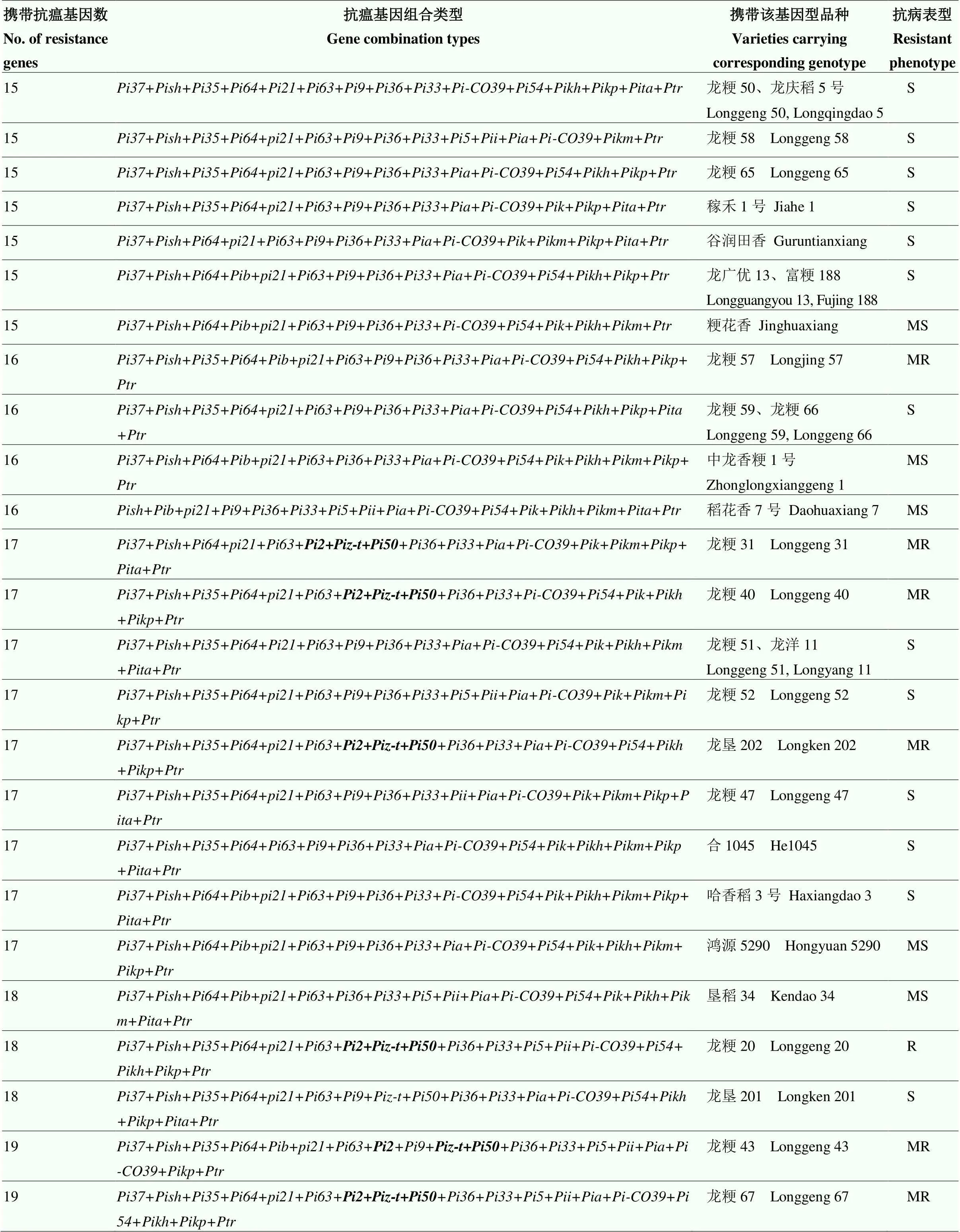
续表4 Continued table 4
粗体“++”表示携带此3个基因组合品种表现抗病反应
The bold of “++indicates that varieties carry three genes show resistance
3 讨论
3.1 黑龙江省水稻品种抗瘟性评价与应用价值
水稻品种抗瘟性鉴定对于监测品种持久抗性及筛选优异抗病材料具有重要意义。本研究中供试品种的抗瘟性评价结果表明,龙粳20抗性频率最高,抗性评价为R,其次是龙粳67、龙垦202、龙粳40、龙粳31、龙粳57、龙粳43等6个品种抗瘟性较好,同时也是黑龙江省水稻种植重要品种,如龙粳31具有早熟、高产、耐寒、优质、抗病、抗倒等诸多优点[20],在黑龙江省种植面积在90万hm2以上。针对黑龙江省水稻品种抗稻瘟病菌的接种鉴定,王桂玲等[6]研究发现龙粳20和龙粳31等品种具有较高抗瘟性;周弋力[12]研究表明龙粳31、龙粳40、龙粳43等32个品种具有良好抗性,稼禾1号抗性表现较差,以上结果与本研究结果相似,因此,在黑龙江省实施单品种布局,可优先考虑推广种植龙粳20、龙粳40、龙粳31、龙粳43等抗谱较高的品种,而稼禾1号等抗性较差的品种已有潜在的感病风险,气候允许条件下很有可能造成稻瘟病大流行。以上研究表明黑龙江水稻种质资源绝大部分仍保有良好抗性,而本研究显示大部分供试品种的抗瘟性偏低,推测出现此差异的原因:一是研究对象存在较大不同;二是寄主垂直抗病性具有小种专化性,不同菌株接种产生的抗性表现存在差异;三是随着使用年限的增长及田间菌群环境的变化,部分品种的抗病性会有所下降。
有研究证明水稻遗传多样性的品种混合栽培能减轻稻瘟病发生[21],辛威[11]研究表明龙粳31与其他9个水稻品种的组合对稻瘟病均表现高抗;郭丽颖等[7]研究表明稻花香与龙粳21联合等22个组合具有很好的防病效果。本研究分析抗性频率较高的15个品种两两组合的联合抗瘟性,剖析品种搭配种植的应用价值,结果表现为4种情况:一是RAC值较高,VAC值较低,表明2个品种的抗病遗传基础互补性高,感病遗传基础同质性低,对于病原菌的侵染可以起到错位作用,从而避免稻瘟病的大发生,这种搭配结构在农业生产上具有较好的应用前景,如龙粳20与龙粳67联合;二是RAC值与VAC值均较高,表明2个品种的抗感遗传亲缘关系较近,在某一年内搭配种植可能会表现良好抗性,但若病菌致病型发生改变就可能引起病害大发生,从农业种植稳定化方面考虑,该类种植结构不适合长时间大面积推广,如龙粳40与龙粳31联合;三是RAC值与VAC值均较低,菌株使搭配双方同时抗病或者同时感病的概率较小,表明2个品种的遗传信息异质性较大,亲缘关系较远,这种种植结构不能体现联合搭配的必要性,在多品种布局条件下应用价值不大,如龙粳67与稻花香7号联合;四是RAC值较低,VAC值较高,说明2个品种的感病遗传背景相似度高,感病程度高,这种类型的种植结构有较大感病风险,如中龙香粳1号与垦稻41联合。
本研究明确了黑龙江省部分水稻品种的抗瘟性及两两搭配的联合抗病性,但随田间环境的变化及稻瘟病菌的不断变异,应继续对选育的水稻品种进行持续的抗性监测,并注重选用远缘的抗病材料和抗病基因资源,培育广谱抗病性品种,丰富黑龙江省水稻种质资源。
3.2 抗瘟基因在黑龙江省水稻种质资源中的分布及抗病相关性
抗瘟基因的挖掘与利用是选育抗病品种的关键性因素。本研究中供试品种的抗瘟基因鉴定结果表明,抗瘟基因、、和在供试品种中均被检测到,、、、、、、、、、、、和检出率为50%—100%,说明这类基因在黑龙江省水稻育种中应用较广泛。、、、、、和检出率为10%—50%,仅在2个品种中被检出,仅在吉粳88中被检出,而、、、、、、、和在供试品种中均未检测到,说明这类基因在黑龙江省水稻品种中分布较少,具有提高产量和持久抗病的新调控机制[2],应用前景广泛,吉粳88值得深入研究其抗病机制。周弋力[12]研究发现34份黑龙江省水稻主栽品种中抗瘟基因、、、、和的分布频率分别为70.6%、55.9%、52.9%、23.5%、23.5%和2.9%;辛威[11]研究显示225份黑龙江省水稻品种资源中抗瘟基因、、、、和的分布频率分别为77.33%、68.44%、63.11%、57.33%、14.67%和9.33%,以上结果与本研究结果相似。供试品种携带抗瘟基因12—19个不等,共58种抗瘟基因型,表明供试品种抗瘟基因组合类型比较丰富。
对单基因及多基因聚合与水稻抗病相关性分析发现,大部分抗瘟基因的分布频率远高于携带该基因且表现抗病的百分率,说明携带这类基因的水稻品种既可能表现为抗病也可能表现为感病,、、、和的分布频率与携带该基因且表现抗病的百分率相差不大,说明携带该类基因的水稻品种有较高概率表现抗病,这与相亚超等[13]研究发现、对黑龙江省水稻抗稻瘟病贡献较大有相类似的结论。研究发现携带相同抗瘟基因型的品种,抗性表现基本一致,表明抗瘟基因型是决定品种抗病程度的关键因素。分析品种抗瘟基因数量与抗病性关系发现,两者之间存在一定程度的正相关作用,与辛威[11]研究发现供试材料的抗病性随抗病基因数目增多呈上升趋势,与汪文娟等[22]研究发现品种中携带抗瘟基因数量越多,其表现抗病的概率就越高结论一致。本研究发现有品种携带十几个抗瘟基因仍表现感病,可能是由于某些抗病基因之间存在拮抗作用反而降低了水稻抗病性,仍需进一步研究验证,这与李思博[23]表明抗病基因可能存在某种关联,或是协同或是拮抗推测一致。携带+++聚合类型的6份品种均抗病,推测该基因型与其他抗瘟基因聚合或与未知新抗病基因互作可能对抗病性起正向调节作用,对水稻抗病性有一定的贡献,这与徐未未等[24]研究表明某些抗性基因之间存在互作,可协同抵抗单个基因不能抵抗的小种,增强抗病性结论一致,可见多基因聚合不是简单的累积效应,而是存在更复杂的互作效果。因此,在今后的抗病育种工作中仍需增加水稻鉴定材料,更全面地掌握抗瘟基因的分布情况,明确不同抗瘟基因聚合类型的抗病效应,进一步通过分子标记、转基因等手段改良品种抗性,培育持久广谱抗病后代。
4 结论
参试的黑龙江省水稻种质资源龙粳20、龙粳67、龙垦202、龙粳40、龙粳31、龙粳57、龙粳43的抗性频率较高,抗病性较好,而稼禾1号等抗性频率相对较低的品种已存在潜在的感病风险。龙粳20与龙粳67等联合抗病性系数较高、联合毒力系数较低的搭配结构具有良好应用前景。抗瘟基因、、、、、、、、、、、、、、、、、、、、、、、、和以不同频率分布在供试品种中,而供试品种均不携带、、、、、、、和。携带、、、和的水稻品种有较高概率表现抗病,品种携带抗瘟基因数量越多其表现为抗病品种的概率越高,+++聚合类型对水稻抗病性有一定的贡献。参试品种抗瘟性表现不同,选择相对抗病品种合理种植或品种联合使用会有良好应用前景,参试品种中抗性基因分布不等,但能通过分析抗瘟基因的有效性来指导品种的合理利用与抗瘟育种方向,以控制病害大流行。
[1] Couch B C, Kohn L M. A multilocus gene genealogy concordant with host preference indicates segregation of a new species,, from.Mycologia, 2002, 94(4): 683-693.
[2] DENG Y W, ZHAI K R, XIE Z,YANG D Y, ZHU X D, LIU J Z, WANG X, QIN P, YANG Y Z, ZHANG G M, LI Q, ZHANG J F, WU S Q,Joëlle M, MAO B, WNAG E T, XIE H A, DIDIER T, HE Z H. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science, 2017, 355(6328): 962-965.
[3] ZHANG Y L, ZHU Q L, YAO Y X, ZHAO Z H, CORRELL J C, WANG L, PAN Q H. The race structure of the rice blast pathogen across southern and northeastern China.Rice, 2017, 10(1): 46.
[4] 田大刚.介导稻瘟病抗性的蛋白质组学分析及抗病基因座的应用[D]. 福建: 福建农林大学, 2017.
TIAN D G. Proteomic analysis ofmediated rice blast resistance and application of the resistant locus[D]. Fujian: Fujian Agriculture and Forestry University, 2017. (in Chinese)
[5] 阎勇, 马增凤, 秦钢, 陈远孟, 秦媛媛, 颜群, 刘驰, 张月雄, 黄大辉. 华南常用籼稻亲本稻瘟病抗性评价及抗性基因鉴定. 南方农业学报, 2017, 48(4): 587-593.
YAN Y, MA Z F, QIN G, CHEN Y M, QIN Y Y, YAN Q, LIU C, ZHANG Y X, HUANG D H. Evaluation of blast resistance and identification of resistance genes in main indica rice patent materials in South China. Journal of Southern Agriculture, 2017, 48(4): 587-593. (in Chinese)
[6] 王桂玲, 宋成艳, 刘乃生, 周雪松. 黑龙江省水稻区(生)试品种(系)对稻瘟病抗性评价与推广. 黑龙江农业科学, 2012(12): 1-4.
WANG G L, SONG C Y, LIU N S, ZHOU X S. Evaluation and extension of rice varieties(lines) in regional(performance)test resistant to blast in Heilongjiang province. Heilongjiang Agricultural Sciences, 2012(12): 1-4. (in Chinese)
[7] 郭丽颖, 赵宏伟, 王敬国, 刘化龙, 孙健, 宋微, 姜思达, 兴旺, 邹德堂. 黑龙江省稻瘟病菌生理小种鉴定和主栽水稻品种抗病性及遗传多样性分析. 核农学报, 2015, 29(8): 1444-1454.
GUO L Y, ZHAO H W, WANG J G, LIU H L, SUN J, SONG W, JIANG S D, XING W, ZOU D T. Identification of physiological race of rice blast fungus and disease resistance and genetic diversity a analysis on major cultivars in Heilongjiang province. Journal of Nuclear Agricultural Sciences, 2015, 29(8): 1444-1454. (in Chinese)
[8] 张亚玲, 赵宏森, 曹有鑫, 付天恒, 杨树, 靳学慧. 抗性频率、联合致病性系数和联合抗病性系数在水稻品种抗瘟性评价中的应用. 江苏农业科学, 2016, 44(7): 158-161.
ZHANG Y L, ZHAO H S, CAO Y X, FU T H, YANG S, JIN X H. Application of resistance frequency, joint pathogenic coefficient and joint disease resistance coefficient in the evaluation of blast resistance of rice varieties. Jiangsu Agricultural Science, 2016, 44(7): 158-161. (in Chinese)
[9] 韩笑, 贺叶晨星, 张梅, 雷子伊, 张涛, 李建粤. 利用分子标记分析22种水稻10个抗稻瘟病基因的基因型. 上海师范大学学报(自然科学版), 2017, 46(5): 654-661.
HAN X, HE Y C X, ZHANG M, LEI Z Y, ZHANG T, LI J Y. Genotyping of 10 blast resistance genes in 22 rice varieties by molecular marker. Journal of Shanghai Normal University (Natural Sciences), 2017, 46(5): 654-661. (in Chinese)
[10] 王亚, 陈献功, 尹海庆, 王越涛, 杨瑞芳, 臧之光, 王生轩. 河南主要水稻种质资源中抗稻瘟病基因的分子检测. 分子植物育种, 2018, 16(10): 3203-3212.
WANG Y, CHEN X G, YIN H Q, WANG Y T, YANG R F, ZANG Z G, WANG S X. Molecular detection of rice blast resistance gene in the main rice germ plasms in henan province. Molecular Plant Breeding, 2018, 16(10): 3203-3212. (in Chinese)
[11] 辛威. 寒地粳稻种质资源稻瘟病抗性鉴定及基因定位[D]. 哈尔滨: 东北农业大学, 2017.
XIN W, Identification and genetic mapping of rice blast resistance in japonica rice germplasm resources in cold region[D]. Harbin: Northeast Agricultural University, 2017. (in Chinese)
[12] 周弋力. 黑龙江省水稻主栽品种抗瘟性评价及稻瘟病菌无毒基因分析[D]. 大庆: 黑龙江八一农垦大学, 2019.
ZHOU Y L. Resistance evaluation of main rice varieties in Heilongjiang province and a virulence genotyping of[D]. Daqing: Heilongjiang Bayi Agricultural University, 2019. (in Chinese)
[13] 相亚超, 王丽丽, 徐凡, 马殿荣. 抗稻瘟病基因在黑龙江水稻资源中的分布. 分子植物育种, 2018, 16(23): 7705-7717.
XIANG Y C, WANG L L, XU F, MA D R. Study on the distribution of rice blast resistant genes in rice resources of Heilongjiang province. Molecular Plant Breeding, 2018, 16(23): 7705-7717. (in Chinese)
[14] 蒋金芬, 韩红萍, 梁友方. 滤纸片法低温冷冻保存菌种的实验室应用. 中国公共卫生, 2006, 22(3): 310.
Jiang J F, Han H P, Liang Y F. Laboratory application of filter paper method for cryopreservation. Chinese Journal of public health, 2006, 22(3): 310. (in Chinese)
[15] 连兆煌. 无土栽培原理与技术. 北京: 农业出版社, 1994.
Lian Z H.Principles and techniques of soilless cultivation. Beijing: Agriculture Press, 1994. (in Chinese)
[16] 孟峰, 张亚玲, 靳学慧, 张晓玉, 姜军. 黑龙江省稻瘟病菌无毒基因、和的检测与分析. 中国农业科学, 2019, 52(23): 4262-4273.
MENG F, ZHANG Y L, JIN X H, ZHANG X Y, JIANG J. Detection and analysis ofavirulence genes,andin Heilongjiang province. Scientia Agricultura Sinica, 2019, 52(23): 4262-4273. (in Chinese)
[17] 靳学慧, 马汇泉. 农业植物病理学. 赤峰: 内蒙古科学技术出版社, 1999.
Jin X H, MA H Q. Agricultural plant pathology.Chifeng: Inner Mongolia Science and Technology Press, 1999. (in Chinese)
[18] 阮宏椿, 杨秀娟, 陈双龙, 董瑞霞, 陈福如, 王伟新, 杜宜新. 福建省水稻新品种对稻瘟病菌的抗性鉴定与评价. 福建农业学报, 2006(4): 304-307.
RUAN H C, YANG X J, CHEN S L, DONG R X, CHEN F R, WANG W X, DU Y X, Identification and evaluation of rice new varieties resistant toin Fujian province. Fujian Journal of Agricultural Sciences, 2006(4): 304-307. (in Chinese)
[19] 张亚玲, 王宝玉, 台莲梅, 郑雯, 邓本良, 靳学慧. 黑龙江省稻瘟病菌生理小种对水稻品种致病性分析. 中国植保导刊, 2014, 34(2): 22-26.
ZHANG Y L, WANG B Y, TAI L M, ZHENG W, DENG B L, JING X H. Analysis on the pathogenic of rice blast fungus physiological race to rice varieties in Heilongjiang province. China Plant Protection, 2014, 34(2): 22-26. (in Chinese)
[20] 李洪亮, 柴永山, 孙玉友, 高春艳, 魏才强, 解忠, 张巍巍, 刘丹, 程杜娟,侯国强, 徐德海, 赵云彤. 寒地超级稻龙粳31祖先亲本追溯及遗传基础解析. 植物遗传资源学报, 2016, 17(3): 433-437.
LI H L, CHAI Y S, SUN Y Y, GAO C Y, WEI C Q, XIE Z, ZHANG W W, LIU D, CHENG D J, HOU G Q, XU D H, ZHAO Y T. Ancestors tracking and analysis on genetic basis of super rice Longjing31 in cold region. Journal of Plant Genetic Resources, 2016, 17(3): 433-437.
[21] YADAV M K, ARAVINDAN S, NGANGKHAM U, RAGHU S, PRABHUKARTHIKEYAN S R, KEERTHANA U, MANDI B C, ADAK T, MUNDA S, DESHMUKH R, PRAMESH D, SAMANTARAY S, RATH P C. Blast resistance in Indian rice landraces: Genetic dissection by gene specific markers.PloS one, 2019, 14(1): e0211061.
[22] 汪文娟, 周继勇, 汪聪颖, 苏菁, 封金奇, 陈炳, 冯爱卿, 杨健源, 陈深, 朱小源. 八个抗稻瘟病基因在华南籼型杂交水稻中的分布. 中国水稻科学, 2017, 31(3): 299-306.
WANG W J, ZHOU J Y, WANG C Y, SU J, FENG J Q, CHEN B, FENG A Q, YANG J Y, CHEN S, ZHU X Y. Distribution of eight rice blast resistance genes in indica hybrid rice in China. Chinese Journal of Rice Science, 2017, 31(3): 299-306. (in Chinese)
[23] 李思博. 辽宁省稻瘟菌无毒基因及水稻抗瘟基因鉴定[D]. 沈阳: 沈阳农业大学, 2018.
LI S B. Identification of avirulent genes and resistance genes ofin Liaoning province[D]. Shenyang: Shenyang Agricultural University, 2018. (in Chinese)
[24] 徐未未, 王兴, 黄永相, 蒋世河, 李伟, 郭建夫. 水稻抗稻瘟病基因的分子标记与标记辅助育种研究进展. 江苏农业学报, 2013, 29(4): 898-906.
XU W W, WANG X, HUANG Y X, JIANG S H, LI W, GUO J F. Advances in molecular markers of blast resistance genes and marker-assisted breeding in rice. Jiangsu Agricultural Sciences, 2013, 29(4): 898-906. (in Chinese)
Evaluation of Rice blast resistance and genetic structure analysis of rice germplasm in Heilongjiang Province
ZHANG YaLing, GAO Qing, ZHAO Yuhan, LIU Rui, FU Zhongju, LI Xue, SUN Yujia, JIN XueHui
College of Agronomy, Heilongjiang Bayi Agricultural University/Heilongjiang Plant Resistance Research Center, Daqing 163000, Heilongjiang
【】Rice blast seriously threatens rice production in Heilongjiang Province and the breeding and utilization of anti-blast varieties are the most economical, safe and effective measures to control it. This study explored the resistance of rice varieties in Heilongjiang Province; clarified the resistance effect of disease resistance genes, and provided a basis for the selection and utilization of germplasm resources of rice blast in Heilongjiang Province. 【】In the autumn of 2018, 134 rice strains were collected from the main rice area in Heilongjiang Province, and the resistance of 50 dominant rice varieties was analyzed. For the reported 35 rice blast resistance genes, a comparison was made between the varieties and positive control varieties. Sequencing results of some positive control varieties were compared with the reference sequence published in the National Centre for Biotechnology Information to analyze the existence of rice blast resistance genes in the corresponding varieties. Through the correlation analysis of gene aggregation type and breed resistance performance, the genotypes related to the resistance performance of rice varieties in Heilongjiang Province were clarified. 【】Among 50 rice varieties in Heilongjiang Province, Longgeng 20 had the best resistance (R). Longgeng 67, Longken 202, Longgeng 40, Longgeng 31, Longgeng 57 and Longgeng 43 had moderate disease resistance; whereas, 43 varieties such as Jiahe 1 had relatively low resistance. Through the analysis of variety combination resistance, we found that 33 pairs of Longgeng 20 + Longgeng 67 had a high RAC(resistance association coefficient) value, low VAC (virulence association coefficient) value and good combined disease resistance. The matching structure showed potential for application. The identification of resistance genes carried by specific primers showed that,,andwere detected, and,,,,,,,,,,,andwere between 50%-100% of occurences frequency, indicating that such genes were widely used in rice breeding in Heilongjiang Province. The detection rate of,,,,,0 andwas between 10% and 50%;was detected in in two varieties, andwas detected in Jigeng 88. However,,,,,,,,andwere not detected in the cultivars, indicating that such genes are less distributed in rice varieties in Heilongjiang Province. The variety genotype analysis found that the varieties carried 12-19 resistant genes, with a total of 58 genotypes, indicating that the test varieties had rich blast resistance gene combination types. By analysis, the single gene and gene polymerization with disease resistance showed that the distribution frequency of,andcorresponded to frequency of resistance. We found that the more varieties carrying resistance genes, the higher the frequency resistance, and six varieties that carrying+++αpolymerization types showed resistance. 【】Our results have demonstrated the following: the resistance of rice germplasm resources in Heilongjiang province is low; combination planting of different varieties could be usefully applied; the distribution of blast resistance genes is different in the participating varieties;,,,andplay a leading role in disease resistance, and+++gene polymerization type could contribute to improving rice blast resistance.
rice; rice blast; resistance frequency; blast resistance evaluation; blast resistance gene

10.3864/j.issn.0578-1752.2022.04.001
2021-08-11;
2021-11-15
国家自然科学基金(U20A2025)、黑龙江省农垦总局科技攻关项目(HKKYZD190205)、黑龙江八一农垦大学科研启动项目(XDB201802,XDB201605)
张亚玲,E-mail:byndzyl@163.com。通信作者靳学慧,E-mail:jxh2686@163.com
(责任编辑 李莉)
