温度和干旱胁迫对3种大黄种子萌发和幼苗生长的影响
2022-04-19王雪茹李依民张明英颜永刚
李 慧,王雪茹,李依民*,徐 进,刘 东,高 静,张明英,杜 弢,颜永刚,张 岗*
温度和干旱胁迫对3种大黄种子萌发和幼苗生长的影响
李 慧1, 2,王雪茹1,李依民1, 2*,徐 进3,刘 东4,高 静1, 2,张明英1,杜 弢4,颜永刚1,张 岗1, 2*
1. 陕西中医药大学药学院/陕西省秦岭中草药应用开发工程技术研究中心,陕西 西安 712046 2. 陕西省中医药管理局“秦药”研发重点实验室,陕西 西安 712046 3. 镇巴县百花谷现代农牧开发有限公司,陕西 汉中 723600 4. 甘肃中医药大学药学院/甘肃省中药质量与标准研究重点实验室,甘肃 兰州 730099
研究不同温度和聚乙二醇(PEG)模拟干旱胁迫对大黄种子萌发的影响,探讨大黄种子萌发的最适温度和对干旱胁迫的耐性。用6个温度(10、15、20、25、30、35 ℃)和6个聚乙二醇(PEG-6000,以下简称PEG)浓度(0、5%、10%、15%、20%、25%)模拟温度胁迫与干旱胁迫对大黄种子进行处理,置人工培养箱培育,借助SPSS 24.0单因素方差分析法对其发芽势、发芽率、胚根、胚轴和子叶等指标统计分析。温度与干旱胁迫对3种大黄种子萌发及幼苗生长有显著影响(<0.05);同一温度下,3种大黄的种子萌发指标随PEG浓度的升高而降低,掌叶大黄和唐古特大黄幼苗各指标亦是随PEG浓度的升高而降低,而药用大黄幼苗各指标随着PEG浓度的升高先增加后减小,在低浓度PEG(5%)达到最大,25%PEG重度干旱时严重抑制掌叶大黄和唐古特大黄萌发(<0.05),完全抑制药用大黄萌发;20 ℃或25 ℃下,各PEG浓度处理的3种大黄种子萌发及幼苗生长指标均达到最大值;在低于20 ℃或者高于25 ℃时,各PEG浓度下的指标呈下降趋势,35 ℃高温下种子萌发和幼苗生长指标随着PEG浓度的升高呈直线下降。掌叶大黄和唐古特大黄种子在各处理下所有指标均高于药用大黄,萌发的速度和整齐度较好,种子的活力高,表现出较强的抗旱性,轻度干旱环境有利于药用大黄幼苗生长,为大黄种质资源评价及规范种植研究与应用提供了理论依据。
大黄;干旱;温度;交互胁迫;种子萌发;幼苗生长
大黄来源于蓼科多年生高大草本掌叶大黄L.、唐古特大黄Maxim. ex Balf.和药用大黄Baill的干燥根及根茎,泻下攻积、清热泻火、凉血解毒、逐瘀通经,用于治疗实热积滞便秘、血热吐衄、目赤咽肿、痈肿疔疮等症[1]。现已从大黄中解析有160多种化学成分,其中以蒽醌类为代表活性成分,在抗炎、抗肿瘤、保护心血管、保肝、改善脑损伤等方面作用显著[2-3]。作为常用大宗药材,大黄野生资源远不足于市场供给。为保障临床药源充足,大黄规范化种植已有一定规模,现主产于甘肃、青海、四川、陕西与湖北等省高海拔区域[4]。不同产区环境条件差异对大黄生长发育及次生代谢的影响,可能是高品质大黄种植生产中面临的首要科学问题。其中,以气候和海拔造成的温度与十分差异直接影响大黄种子萌发生理而显得尤为关键。
种子萌发是植物体生长发育的先决条件,种子萌发特性与温度、水分或光照等环境条件关系密切,是决定植物逆境生理适应的关键环节[5]。温度是影响植物种子萌发的主要因素之一[6],其通过调节细胞膜通透性及酶活性影响种子活力及萌发[7]。大量研究发现不同植物种子对温度的响应规律以及最佳萌发温度均有差异,适宜的温度可以提高种子发芽率,加快发芽速度,利于幼苗生长,过高或过低的温度则抑制种子萌发[8-10]。同时,水分也是决定种子萌发的另一关键因素。干旱是最为普遍的水分胁迫,直接影响植物种子萌发和幼苗生长发育[11]。种子长期处于干旱环境中导致发芽率低、成活率低,植物生长发育不良,影响其质量和产量[12]。自然条件下,环境胁迫通常系温度、干旱或盐等多种因素共存的复合胁迫,对植物种子萌发及形态建成等造成极大影响[13-14]。然而,药用植物生长发育与复合胁迫的关系研究尚不系统,是亟待开展的热点领域。
目前,大黄研究主要集中在化学成分、药理作用及临床应用方面[15-16]。课题组围绕大黄生产实际问题,在国内外率先开展了大黄种质资源、质量评价、基因表达调控等研究工作,取得了一定进展[17-19]。大黄作为高海拔适生物种,对生长环境要求高,但是其在特殊生境下生理生态基础研究匮乏,限制了对大黄品质形成的深入研究和科学理解。水分、温度等复合胁迫在药用植物道地药材的优良性状及品质形成过程中起着独特的作用,但是目前受关注较少,研究非常有限[13]。为此,本研究首次系统开展了3种大黄种子萌发实验,设置10、15、20、25、30、35 ℃ 6个温度梯度,在各温度条件下又分别设置6个聚乙二醇(PEG-6000,以下简称PEG)溶液浓度梯度(0、5%、10%、15%、20%、25%)的干旱胁迫,考察3种大黄种子响应温度和干旱交互条件下的萌发特征和幼苗发育情况,预期为大黄规范化种植和高品质药材生产提供理论支撑。
1 材料与方法
1.1 材料
3种大黄种子样品于2020年9月分别采自甘肃和政县、陕西镇巴县和青海湟中县,经陕西中医药大学王继涛高级实验师鉴定为掌叶大黄L.、药用大黄Baill.和唐古特大黄Maxim. ex Balf.的干燥成熟种子。
1.2 培养条件及方法
在光照培养箱(BSG-300,上海博讯)设置温度梯度模拟外界环境温度,以PEG模拟干旱环境。在光照培养箱设置6个恒温(10、15、20、25、30、35 ℃)条件下,分别设置PEG浓度为0(CK)、5%、10%、15%、20%、25% 6个PEG干旱处理,每个处理3组重复,共108组实验。
实验选用健康饱满的3种大黄种子,去离子水浸泡1 d,使用75%乙醇消毒10 s,10%双氧水消毒15 min。将种子置于铺有两层滤纸的150 mm的培养皿内,种子间保持一定的距离,每皿100粒种子,用注射器将相应浓度的PEG溶液注入底部,随后将培养皿分别放入对应条件的光照培养箱内,光照时间12 h/d,光照强度15 000 lx。以胚根突破种皮1 mm为发芽标准,每天记录萌发情况,培养10 d。试验过程为了排除水分蒸发对PEG浓度的影响,每隔3 d更换1次滤纸和相应浓度的PEG溶液。
1.3 指标测定
按照公式计算,各处理的最终发芽率和发芽势。萌发结束后,从各处理的各重复中随机取出10株幼苗,测定胚根长、胚轴长、子叶长、苗长。
最终发芽率=发芽种子数/供试总种子数
发芽势=发芽前5 d发芽数/供试总种子数
1.4 数据处理
采用Microsoft Excel 2019软件和SPSS 24.0统计分析软件对发芽率、发芽势和幼苗根长株高等数据进行处理绘图、差异显著性检验(LSD法,<0.05)和多因素方差分析,并用Design Expert 8.0软件进行绘图。
2 结果与分析
2.1 温度对干旱胁迫下大黄种子萌发的影响
由图1、2可知,3种大黄种子整体发芽率均随着温度的增加呈现先上升后下降的趋势。20、25 ℃时,掌叶大黄在PEG浓度为5%出现最高值95.56%;20 ℃时,药用大黄对照组出现最高值47.33%;25 ℃时,唐古特大黄对照组出现最高值96.67%;20 ℃时掌叶大黄和唐古特大黄种子的最终发芽率相对稳定,各处理组间(除25% PEG浓度)无显著差异(>0.05),表明20 ℃是掌叶大黄和唐古特大黄种子的最适温度。各处理组随着PEG浓度的增加,药用大黄最终发芽率明显降低(<0.05),而掌叶大黄和唐古特大黄种子降低趋势并不明显(>0.05);当PEG浓度为25%时,掌叶大黄和唐古特大黄种子在30 ℃及35 ℃下不发芽,而药用大黄种子在各温度下均不发芽,且在PEG浓度为20%时,只在20 ℃和25 ℃温度下发芽。

ZY-掌叶大黄 YY-药用大黄 TGT-唐古特大黄 柱状图上不同字母表示3种大黄的所有处理在P<0.05水平差异显著,图2、3同

图2 温度与PEG溶液浓度交互作用下3种大黄种子的累积发芽率
如图3所示,3种大黄种子发芽势整体趋势是先增加后减小。在温度10、15和35 ℃下,药用大黄种子发芽势均为0(除15 ℃及35 ℃对照1.33%和0.67%),30 ℃时,PEG高浓度(20%和25%)的发芽势也为0;掌叶大黄种子在低温10 ℃时,除对照外均为0,15 ℃之后发芽势开始升高,且在20~30 ℃,5%和10% PEG浓度组与对照均无差异(>0.05);10、15和35 ℃时,唐古特大黄种子在20%和25%PEG溶液处理下发芽势均为0,且在所有温度下,低浓度PEG(5%、10%)组与对照均无差异(>0.05),10和15 ℃低温时,发芽势显著高于掌叶大黄(<0.05),其他温度差异不明显(>0.05),而在所有温度下,唐古特大黄和掌叶大黄种子的发芽势均高于药用大黄。
多因素方差分析显示(表1),6种不同的温度和6种不同的PEG浓度对3种大黄种子的最终发芽率、发芽势有显著影响。综合上述结果,在干旱胁迫下,唐古特大黄和掌叶大黄优于药用大黄,20 ℃是掌叶大黄和唐古特大黄种子萌发的最适温度,20和25 ℃是药用大黄种子萌发的最适温度。
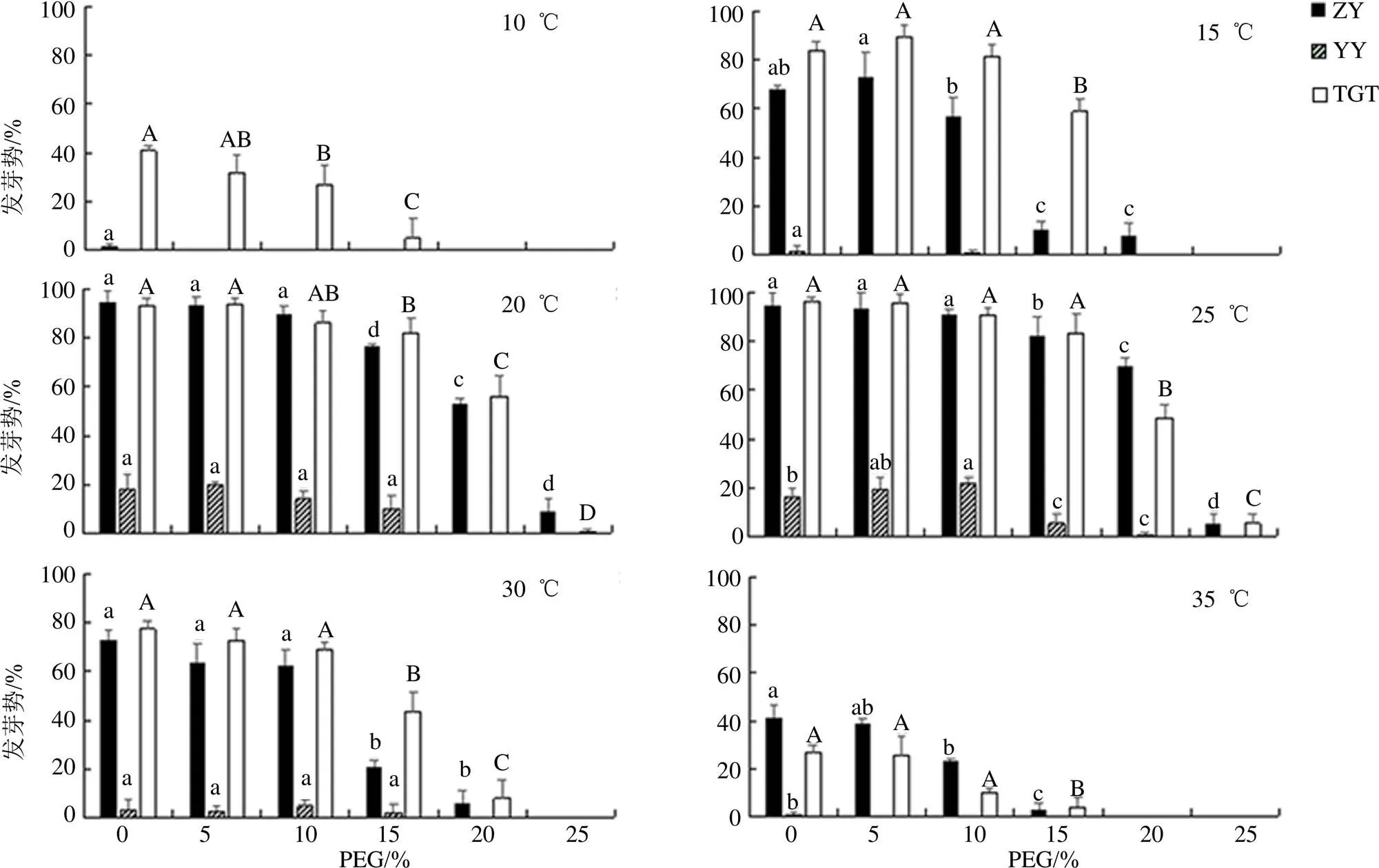
图3 温度与PEG溶液浓度交互作用下3种大黄种子的发芽势
表1 温度与PEG溶液浓度交互作用下3种大黄种子萌发的多因素方差分析(F值)
Table 1 Multivariate an OVA of three rhubarb seeds germination under different treatments (F value)
差异来源F值 发芽势最终发芽率 种子1444.231**1044.651** 温度 551.992** 302.004** 胁迫 512.262** 491.126** 种子+温度 93.788** 22.464** 温度+胁迫 27.920** 14.111** 种子+胁迫 92.825** 45.930** 种子+温度+胁迫 8.683** 4.977**
*<0.05,**<0.01
随着PEG胁迫程度的增加,种子萌发受到的抑制,尤其是PEG浓度25%时。因此,唐古特大黄种子比掌叶大黄种子耐低温耐旱,掌叶大黄比唐古特大黄种子耐高温,药用大黄较掌叶大黄对高温或低温更敏感。
2.2 温度对干旱胁迫下大黄幼苗生长的影响
由表2知,温度由10 ℃升高到35 ℃的过程中,掌叶大黄各处理的胚根长先增后减,25 ℃时达到最大,且随着PEG浓度的升高而缩短;唐古特大黄的胚根长同样是先增后减,同掌叶大黄一致,20 ℃达到最大值,但在PEG浓度为5%之后,25 ℃达到最大值,随后减少;药用大黄的胚根长同样是先增后减,同掌叶大黄一致,25 ℃达到最大值,但当PEG浓度为10%和15%时,20 ℃达到最大值。不考虑未发芽的处理组时,掌叶大黄胚轴长的最大值出现在20 ℃时的对照组,为7.16 mm,最小值出现在15 ℃下PEG浓度为20%时,为1.59 mm;药用大黄胚轴长的最大值出现在25 ℃下PEG浓度为5%,为5.89 mm,最小值出现在35 ℃下PEG浓度为10%,为2.36 mm;唐古特大黄胚轴长的最大值出现在25 ℃下PEG浓度为5%时,为6.52 mm,最小值出现在35 ℃下PEG浓度为10%,为2.14 mm。随着温度变化结合干旱胁迫的影响,胚根和胚轴均发生一定程度的变化(表2、3)。
由表4可知,随着温度的升高,干旱胁迫下掌叶大黄和唐古特大黄幼苗的子叶长呈现先增后减的趋势。掌叶大黄在20 ℃达到最大值;而药用大黄在PEG浓度为5%时,在25 ℃达到最大值,其余同掌叶大黄和唐古特大黄。对照组时掌叶大黄和唐古特大黄幼苗的子叶长度出现最大值,掌叶大黄最大值为7.63 mm,唐古特大黄最大值为8.17 mm;PEG浓度为5%时药用大黄幼苗的子叶长度出现最大值,为7.65 mm。掌叶大黄幼苗苗长在25 ℃的对照组达到最大值;药用大黄苗长在对照组时,25 ℃达到最大值;药用大黄幼苗苗长PEG浓度为5%时,在25 ℃达到最大值,其他浓度时,在20 ℃达到最大值;唐古特大黄幼苗苗长在对照组和PEG浓度为15%时,在20 ℃达到最大值,在其他浓度时,在25 ℃达到最大值(表3、4)。
表2 温度与PEG溶液浓度交互作用下3种大黄幼苗的胚根长和胚轴长()
Table 2 Radicle length and hypocotyl length of three rhubarb seedlings under the interaction of temperature and PEG concentration ()
指标种子温度/℃长度/mm CK5%PEG10% PEG15% PEG20% PEG 胚根ZY109.26±2.42aC10.01±1.63aD——— 1521.45±2.43aB18.27±4.42aB18.66±1.81aB10.27±5.40bC6.78±4.11bC 2041.54±5.49aA41.66±6.70aA29.27±5.65bA25.76±3.94bB16.98±3.43cB 2543.56±9.19aA42.45±3.24aA32.65±7.54bA36.17±9.63abA20.57±3.95cA 3012.19±3.46aC13.53±2.39aC10.18±1.71bC10.07±2.26bC8.12±3.32bC 357.55±1.32aC5.52±1.27bE4.58±0.67bD4.77±0.30bD4.43±0.33bD YY107.98±3.23B———— 159.34±2.41abB12.16±3.99aB7.39±2.44bC6.86±4.04bB— 2020.75±8.45bA27.48±7.15aA26.58±6.13abA16.97±3.05bA— 2521.17±6.17bA29.54±4.63aA22.22±6.24bB8.78±2.91cB— 308.64±2.59aB6.93±1.11abC6.70±1.12bC5.78±1.38cB— 35————— TGT1015.43±3.62aC15.12±2.93aC12.74±2.68abC9.47±1.83bC— 1523.53±4.36abB25.87±6.47aB20.45±4.44bB18.38±5.53bB— 2036.08±6.04aA37.00±3.21aA29.80±4.63bA23.37±4.58cA— 2525.52±7.88bB36.88±7.14aA31.44±5.88aA18.58±6.43cB10.64±6.27dA 3010.57±2.35bD12.78±5.11abC14.27±4.87aC10.64±2.88bC9.16±3.18bA 355.13±0.93abE5.24±1.58aD3.99±0.68bD—— 胚轴ZY102.07±0.51aD1.79±0.42aC——— 154.36±0.65aC4.42±0.75aB4.28±1.47aA2.27±0.80bC1.53±1.22bC 207.16±1.25aA5.92±1.19bA4.79±0.67cA4.56±1.07cA3.19±0.97dA 255.57±1.22aB4.27±1.26bB4.24±0.75bA4.67±0.88bA3.35±0.49cA 304.62±1.23abC5.33±1.05aA4.09±1.02bA3.99±0.65bB3.01±1.32cB 352.89±0.89aD2.65±0.56aC2.24±0.45aB2.67±0.60aC2.36±0.72aB YY102.43±1.23D———— 153.05±0.64abC3.38±0.95aB2.36±0.48bC2.32±0.40bC— 205.56±0.94abA5.73±0.91aA4.85±1.26abA4.77±1.09bA— 255.54±1.08aA5.89±1.36aA4.88±0.91aA3.77±1.25aB— 304.83±1.60aB3.97±0.56bB3.14±0.65bB2.66±3.47cC— 35————— TGT105.01±0.72aB4.32±0.90aC3.10±0.82bC2.42±0.31bC— 155.53±0.80aAB5.16±0.84abB4.70±0.62bB4.19±0.94bB— 205.77±1.54abA6.25±1.48aA5.33±0.91abA4.79±0.88bA— 256.00±1.75aA6.52±0.75aA5.72±0.81aA4.22±0.90bAB3.62±0.91cA 304.92±1.90aB5.04±0.72aB4.57±0.76abB4.05±0.63abB3.68±1.17bA 352.80±0.46abC2.94±0.63aD2.14±0.37bD——
不同字母表示差异显著<0.05,下同
Different letters indicate significant difference˂ 0.05, same as below
2.3 温度对干旱胁迫下大黄种子萌发和幼苗生长影响的互作效应分析
利用Design Expert 8.0软件,以种子类型、外界胁迫、实验温度为考察因素,以最终发芽率、胚根长等测量指标为响应值,来预测不同温度和PEG胁迫下3种大黄种子和幼苗发育的最优处理。由图4可以看出,相同温度下随着胁迫浓度的增加,种子的最终发芽率和幼苗的胚根长、胚轴长、子叶长、苗长减小。最后由软件分析得到最佳处理条件:0.72% PEG、19.30 ℃复合胁迫为掌叶大黄发芽的最优处理组合,0.78%PEG、18.51 ℃为药用大黄发芽的最优处理组合,0.52%PEG、18.09 ℃为唐古特大黄发芽的最优处理组合;适宜3种大黄幼苗发育的处理均为0%PEG时的不同温度下,为掌叶大黄22.32 ℃、药用大黄21.61 ℃、唐古特大黄20.93 ℃。
表3 温度与PEG溶液浓度交互作用下3种大黄幼苗生长的多因素方差分析
Table 3 Multivariate analysis of variance for growth of three rhubarb seedlings under the interaction of temperature and PEG concentration
差异来源胚根长/mm胚轴长/mm子叶长/mm苗长/mm 种子231.448**144.155** 6.779**337.940** 温度646.857**388.082** 10.807**781.500** 胁迫534.149**702.375** 18.991**792.751** 种子+温度 26.048** 29.239** 1.459* 30.334** 温度+胁迫 47.506** 30.423** 1.448* 50.087** 种子+胁迫 19.695** 14.232** 0.733* 24.155** 种子+温度+胁迫 5.739** 8.418** 1.770** 7.472**
*<0.05,**<0.01
表4 温度与PEG溶液浓度交互作用下3种大黄幼苗的子叶长和苗长()
Table 4 Cotyledon length and seedling length of three rhubarb seedlings under the interaction of temperature and PEG concentration ()
指标种子温度/℃长度/mm CK5%PEG10% PEG15% PEG20% PEG 子叶ZY103.92±0.63aD3.88±0.74aC——— 155.94±0.79aC5.69±0.74aB5.02±0.52bB3.22±0.64cC2.93±0.65cB 207.63±1.10aA6.81±1.19bA5.13±0.92bB5.55±0.78bA3.71±0.75cA 256.82±0.96aB5.59±1.15bB5.09±0.95bB5.39±0.99bA3.81±0.88cA 306.38±0.77aBC5.84±0.83aB5.71±0.70aA4.47±1.04bB3.28±2.09bB 353.16±0.57aD3.52±0.90aC3.32±0.55aC3.64±0.51aC3.08±0.32aB YY103.73±4.21D———— 154.34±0.41aC4.38±0.56aB3.24±0.48bB3.09±0.83bD— 206.62±1.07aA6.28±0.87aB5.33±0.96bA4.85±0.87bA— 256.47±1.61bA7.65±0.90aA5.56±1.06cA4.23±0.79dB— 306.00±0.92aB4.42±1.43bB3.73±0.68bB3.96±2.76bC— 35————— TGT105.43±0.46aC4.42±0.52bD3.98±0.54bC2.98±0.52cC— 156.68±1.10aB5.44±0.75bC5.17±0.45bBC4.59±1.07cA— 208.17±1.53aA6.28±1.23bB5.65±1.30bcB5.03±1.15cA— 257.76±0.98aA7.77±0.93aA7.05±0.80bA4.24±0.86cB3.76±0.65bA 305.48±0.64aC5.41±0.58aC4.46±0.82bC3.82±0.71bC3.15±0.77cA 353.74±0.63aD3.56±0.51aE3.18±0.51aD—— 苗ZY1015.82±3.43aD15.57±2.36aC——— 1537.25±7.38aB35.12±3.44aB34.20±6.04aB19.83±7.43bC10.78±5.88cB 2068.97±9.74aA65.94±8.68aA46.74±7.06bA41.00±5.46bB28.46±5.75cA 2574.57±18.67aA67.85±6.98aA52.36±7.14bA52.26±10.45bA31.25±5.40cA 3027.95±4.92aC30.22±3.79aB23.03±3.60bC19.85±2.04bC12.14±2.97cB 3515.94±2.81aD13.23±2.25bC11.56±0.87cD11.96±0.21bcC11.43±1.08cB YY1013.17±5.19C———— 1518.13±3.84aB21.19±5.53aB13.18±2.66bB14.35±4.96bC— 2034.11±6.67abA39.71±9.08aA40.10±8.48aA28.97±4.47bA— 2533.93±8.06bA48.70±4.79aA36.74±6.39bA19.52±4.79cB— 3020.48±4.31aB17.81±3.77abB14.79±1.81bB13.87±3.96bC— 35————— TGT1026.14±2.97aC23.54±3.27abC22.10±3.67bC15.88±3.90cC— 1538.58±3.02aB38.30±6.48aB34.43±4.71aB28.85±4.57bA— 2053.17±7.61aA52.23±2.79aA42.19±6.57bA34.62±4.41cA13.45±4.35dB 2546.21±8.83bA56.35±9.08aA46.35±7.64bA27.25±9.38cA18.94±6.47dA 3025.14±3.88abC27.26±6.38aC24.31±4.07abC21.48±3.18bB17.27±4.86cA 3512.67±1.92aD12.68±1.92aD9.86±1.27bD——

图4 温度与PEG溶液浓度交互作用下对大黄生长影响的响应面
3 讨论
3.1 3种大黄种子萌发及幼苗生长对温度响应差异显著
种子萌发是种子吸水激活代谢反应,随后引起胚膨大和胚根从外周组织中突破的一个过程,其内部进行着一系列酶催化的物质和能量转化,而温度对于维系酶的活性至关重要[20],因此低温或高温均会对种子萌发产生影响。种子内贮藏物可以供给种子萌发及幼苗生长是十分有限的,若遇低温侵袭使出苗延迟,则会因为基础生命代谢的消耗在出苗前耗尽贮藏物[21],高温会影响萌发过程中膜透性和酶的活性,降低代谢活动[22]。只有在适宜的温度下种子的活力才不会受影响,萌发正常出苗整齐。本研究发现3种大黄种子在10、15 ℃的情况下发芽迟缓,药用大黄萌发率较低;在20、25 ℃情况下,种子可以正常萌发及生长,而且可以迅速进入萌发状态;在30 ℃情况下,种子萌发率降低,出苗缓慢;在35 ℃情况下,萌发情况显著降低,药用大黄完全不萌发。唐古特大黄在10 ℃时不论是萌发时间还是萌发率以及幼苗的生长与20、25 ℃时的差异较其他2个大黄最小,因此唐古特大黄对低温的适应性优于其他2种大黄,掌叶大黄在35 ℃时各指标与20、25 ℃时的差异较其他2个大黄最小,因此掌叶大黄对高温的适应性优于其他2种大黄。
3.2 3种大黄种子萌发及幼苗生长对干旱胁迫响应差异显著
水分也是植物生长的先决条件,土壤干旱已经成为影响全球生态系统的重要的环境因子,严重影响植物的生长发育[23]。PEG是一种高分子渗透剂,可以使植物组织和细胞处于类似干旱的水分胁迫中,通常用于植物抗旱性早期鉴定研究,其作用机制是能减缓种子萌发初期水分进出种子的速率,从而控制种子吸水速率和发芽进程[24]。用不同浓度的PEG溶液处理小麦种子测定其萌发指标,5%~10%PEG对种子萌发影响较小,15% PEG延缓种子萌发,当浓度到达30%时,小麦种子几乎不发芽[25];以大花白麻和罗布麻为对象,随着PEG浓度的不断增加,种子含水率减少,种子萌发指标和幼苗生长指标均呈明显的下降趋势,且在25%~30%时受抑制程度最明显[26]。本研究表明低浓度(5%)PEG处理3种大黄,种子发芽率、发芽势以及幼苗胚根、胚轴和子叶与对照无明显差异,甚至对药用大黄幼苗还有轻微促进作用;高浓度(25%)PEG处理时,药用大黄种子萌发完全被抑制,掌叶大黄和唐古特大黄各指标也显著下降;当PEG浓度为15%时,掌叶大黄和唐古特大黄种子萌发与幼苗生长指标均为对照的58%以上。这些结果说明掌叶大黄和唐古特大黄的抗旱性优于药用大黄,这可能系3种大黄生长于不同高海拔山区长期进化的结果。
3.3 温度与干旱复合胁迫对3种大黄种子萌发及幼苗生长有显著影响
自然条件下,植物往往会受到2种或2种以上的非生物胁迫,环境胁迫及在不同胁迫组合会对种子萌发及幼苗生长产生影响[27]。干旱与温度胁迫最常见的两种胁迫因子。本研究发现干旱与温度胁迫交互作用显著影响3种大黄的种子萌发及幼苗生长。各浓度PEG与20、25 ℃交互处理优于各浓度PEG单独处理下3种大黄的种子萌发及幼苗生长,说明适宜的温度会缓减干旱对种子萌发的胁迫。而在10 ℃或35 ℃单独处理下3种大黄种子的萌发及生长情况,其他温度较10 ℃或35 ℃与各浓度PEG交互处理下的好,说明不适宜的胁迫会加速另一种胁迫对种子萌发的抑制。不适宜的温度与干旱胁迫交互作用使种子发生生理失衡,进而降低发芽率、延迟发芽时间,甚至使种子丧失活力[28]。在不同浓度PEG与低温或高温交互处理下,3种大黄种子萌发明显受到抑制,这与前人对黄芪[29]、桔梗[30]、野豌豆[31]等植物种子萌发研究类似,随着温度的下降或者升高,大黄抗旱性也会有一定程度的下降。所以复合胁迫环境中一种胁迫会影响植物对另一种胁迫的抗性。
本研究厘清了3种大黄种子萌发及幼苗生长的最适温度及在干旱胁迫下的生长特征及其差异,为大黄规范化种植生产及品质提高提供了理论依据。然而,3种大黄响应温度和干旱胁迫处理的差异机制尚需深入研究。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典 [S]. 一部. 2020: 23.
[2] Zheng Q X, Wu H F, Guo J,. Review of rhubarbs: Chemistry and pharmacology [J]., 2013, 5(1): 9-32.
[3] Yang F, Xu Y, Xiong A Z,. Evaluation of the protective effect of Rheiet Rhizoma against α-naphthylisothiocyanate induced liver injury based on metabolic profile of bile acids [J]., 2012, 144(3): 599-604.
[4] 陈艳琰, 唐于平, 陈嘉倩, 等. 大黄资源化学研究进展与利用策略 [J]. 中草药, 2018, 49(21): 5170-5178.
[5] 张佳宁, 刘坤. 植物调节萌发时间和萌发地点的机制 [J]. 草业学报, 2014, 23(1): 328-338.
[6] Ding Y L, Shi Y T, Yang S H. Molecular regulation of plant responses to environmental temperatures [J]., 2020, 13(4): 544-564.
[7] 汪建军, 麻安卫, 汪治刚, 等. 不同温度和PEG处理对中华羊茅种子萌发的影响 [J]. 草业学报, 2016, 25(4): 73-80.
[8] Yue S D, Zhou Y, Zhang Y,. Effects of salinity and temperature on seed germination and seedling establishment in the endangered seagrassAsch. & Graebn. in Northern China [J]., 2019, 146: 848-856.
[9] Gu R T, Zhou Y, Song X Y,. Effects of temperature and salinity onsinensis seed germination, seedling establishment, and seedling growth [J]., 2018, 134: 177-185.
[10] 董兰, 徐安凯, 王英哲, 等. 不同处理对长白山野生黄金菊种子萌发的影响 [J]. 分子植物育种, 2019, 17(23): 7879-7885.
[11] 邹京南, 曹亮, 王梦雪, 等. 外源褪黑素对干旱胁迫下大豆结荚期光合及生理的影响 [J]. 生态学杂志, 2019, 38(9): 2709-2718.
[12] Thabet S G, Moursi Y S, Karam M A,. Genetic basis of drought tolerance during seed germination in barley [J]., 2018, 13(11): e0206682.
[13] 蒋待泉, 王红阳, 康传志, 等. 复合胁迫对药用植物次生代谢的影响及机制 [J]. 中国中药杂志, 2020, 45(9): 2009-2016.
[14] Zandalinas S I, Fichman Y, Devireddy A R,. Systemic signaling during abiotic stress combination in plants [J]., 2020, 117(24): 13810-13820.
[15] 王玉, 杨雪, 夏鹏飞, 等. 大黄化学成分、药理作用研究进展及质量标志物的预测分析 [J]. 中草药, 2019, 50(19): 4821-4837.
[16] Zheng J, Fan R, Wu H Q,. Author Correction: Directed self-assembly of herbal small molecules into sustained release hydrogels for treating neural inflammation [J]., 2020, 11(1): 3815.
[17] 黑小斌, 李欢, 李依民, 等. 药用大黄幼苗转录组高通量测序及蒽醌类生物合成基因筛选 [J]. 中国药学杂志, 2019, 54(7): 526-535.
[18] 李欢, 张娜, 李依民, 等. 利用转录组测序挖掘掌叶大黄蒽醌类生物合成相关基因 [J]. 药学学报, 2018, 53(11): 1908-1917.
[19] 黑小斌, 李依民, 李欢, 等. 掌叶大黄种子特性研究及无菌培养体系的构建 [J]. 中草药, 2019, 50(18): 4430-4437.
[20] 程昕昕, 刘言龙, 牛永胜, 等. 普甜玉米种子萌发期糖代谢和水解酶活性动态变化 [J]. 广西植物, 2015, 35(3): 348-353.
[21] Xia Q, Ponnaiah M, Cueff G,. Integrating proteomics and enzymatic profiling to decipher seed metabolism affected by temperature in seed dormancy and germination [J]., 2018, 269: 118-125.
[22] Zhang H, Zhang J Y, Xu Q Y,. Identification of candidate tolerance genes to low-temperature during maize germination by GWAS and RNA-seqapproaches [J]., 2020, 20(1): 333.
[23] 邓东周, 贺丽, 鄢武先, 等. 温度和干旱胁迫对川西北高寒沙区3种优势植物种子萌发的影响 [J]. 种子, 2019, 38(3): 15-18.
[24] 李娜, 罗俊杰, 张仁陟, 等. 持续模拟干旱胁迫对胡麻萌发特性影响及品种抗旱性评价研究 [J]. 核农学报, 2016, 30(2): 379-387.
[25] 魏佳丽, 崔继哲, 赵鹤翔, 等. 盐碱与干旱胁迫对碱菀种子萌发和TvNHX1表达的影响 [J]. 应用生态学报, 2010, 21(6): 1389-1394.
[26] 韩福贵, 徐先英, 马全林, 等. 大花白麻和罗布麻种子萌发对干旱胁迫的响应 [J]. 西北林学院学报, 2021, 36(1): 139-143, 256.
[27] 郭兰萍, 周良云, 康传志, 等. 药用植物适应环境胁迫的策略及道地药材“拟境栽培” [J]. 中国中药杂志, 2020, 45(9): 1969-1974.
[28] 任瑞芬, 杨秀云, 尹大芳, 等. 4种薄荷种子萌发对干旱与低温的响应 [J]. 草业科学, 2015, 32(11): 1815-1822.
[29] 徐蓉蓉, 高静, 任炳浩, 等. 温度对干旱、盐胁迫下两种黄芪属种子萌发和幼苗生长的影响 [J]. 生态学杂志, 2020, 39(9): 2930-2943.
[30] 仉劲, 李国清, 于清伟. 干旱胁迫与温度变化对粉花桔梗种子萌发特性的影响 [J]. 农学学报, 2018, 8(6): 58-61.
[31] 伊风艳, 孙海莲, 晔薷罕, 等. 温度和干旱胁迫对乌拉特肋脉野豌豆种子萌发的影响 [J]. 内蒙古农业大学学报: 自然科学版, 2019, 40(5): 43-49.
Effects of temperature and drought stress on seed germination and seedling growth of three rhubarb species
LI Hui1, 2, WANG Xue-ru1, LI Yi-min1, 2, XU Jin3, LIU Dong4, GAO Jing1, 2, ZHANG Ming-ying1, DU Tao4, YAN Yong-gang1, ZHANG Gang1, 2
1. Shaanxi University of Chinese Medicine of Pharmacy/Shaanxi Qinling Chinese Herbal Medicine Application Development Engineering Technology Research Center, Xi’an 712046, China 2. Key Laboratory for Research of "Qin Medicine" of Shaanxi Administration of Traditional Chinese Medicine, Xi’an 712046, China 3. Baihua Valley Modern Agriculture and Animal Husbandry Development Co., Ltd. of Zhenba County, Hanzhong 723600, China 4. College of Pharmacy, Gansu University of Chinese Medicine/Key Laboratory of Quality and Standard of Traditional Chinese Medicine in Gansu Province, Lanzhou 730099, China
To study the effect of different temperature and PEG-6000 simulated drought stress on seed germination of Rhubarb, exploring the optimum temperature and tolerance of seed germination of rhubarb under drought stress.To use six temperatures (10, 15, 20, 25 and 30, 35oC) and six PEG concentrations (0, 5%, 10%, 15%, 20%, 25%) to simulate the temperature stress and drought stress to deal with rhubarb seeds, artificial culture box, with the help of SPSS 24.0 single factor variance analysis on the germination potential, germination rate, germination index, radical length, hycopotyl length, cotyledon length index statistics and analysis.Drought and temperature had significant effects (<0.05) on the seed germination and seedling growth of these three rhubarbs. Under the same temperature condition, seed germination indexes deceased with the increasement of PEG concentrations for these species. Forand, seedling growth indexes decreased as PEG concentrations increased.seedling growth indexes showed increased then decreased and reached peak at 5% PEG. 25% PEG severely repressedandgermination, and completely inhibitedgermination. At 20oC or 25oC, seed germination and seedling growth indexes of the three rhubarbs all reached maximum under various PEG concentrations. At temperatures below 20oC or over 25oC, seed germination and seedling growth indexes displayed decreasing trend, while showing decreased dramatically at 35oC high temperature.Over all, under all experimental conditions, indexes ofandseeds showed better germination speed and uniformity, higher seed vigor, and stronger drought resistance than those of. Mild drought might be good for seedling growth of. This study provided a theoretical basis for the germplasm assessment, standard cultivation research and applications of rhubarb.
rhubarb; drought; temperature; interactive stress; seed germination; seedling growth
R286.2
A
0253 - 2670(2022)08 - 2480 - 10
10.7501/j.issn.0253-2670.2022.08.025
2021-10-16
国家自然科学基金资助项目(81973430);国家自然科学基金资助项目(82104334);陕西省重点产业创新链项目(2021ZDLSF04-01);陕西中医药大学新进博士科研启动经费(104080001);陕西中医药大学学科创新团队项目(2019-QN01);省部共建特色秦药资源研究开发国家重点实验室(培育)开放课题(QY202101)
李 慧,女,硕士研究生,研究方向为中药资源与分子生药学。E-mail: 1445784065@qq.com
张 岗,男,博士,教授,研究方向为中药资源与分子生药学。E-mail:jay_gumling2003@aliyun.com
李依民,女,博士,副教授,研究方向为中药资源评价与利用。E-mail: 2051058@sntcm.edu.cn
[责任编辑 时圣明]
