柳江鱼类群落结构及多样性研究
2022-04-13朱书礼陈蔚涛李新辉李跃飞
朱书礼 陈蔚涛 李新辉 李 捷 李跃飞
(中国水产科学研究院珠江水产研究所, 国家渔业资源环境广州观测实验站, 农业农村部珠江中下游渔业资源环境科学观测实验站, 广东省水产免疫技术重点实验室, 广州 510380)
柳江是珠江水系西江干流的第二大支流, 发源于贵州省独山县, 在广西壮族自治区象州县注入西江(红水河段), 地理位置107°27′—110°34′E, 23°41′—26°30′N, 河长755 km, 流域面积58270 km2[1,2]。柳江上游称都柳江, 入广西三江县后称融江, 流至柳城县后称柳江。柳江地形独特, 生境复杂, 蕴含着丰富多样的物种[3]。据不完全统计, 柳江共记录鱼类141种[2]。柳江为区域经济发展提供了重要支撑,但随着经济快速发展, 河道开发和水体环境污染加剧[4,5], 导致河流生境破碎化、径流分布时空格局改变, 鱼类自然栖息环境受到的干扰加剧, 河流生态系统受到严重威胁; 加之酷渔滥捕、过度捕捞,柳江鱼类资源及多样性呈现衰退趋势[6]。
在淡水生态系统中, 鱼类是生态系统功能最敏感和最可靠的指标之一[7,8], 维持鱼类的多样性是渔业可持续发展的基础。了解鱼类群落结构和多样性特征, 研究其对水域环境变化的响应机制, 对渔业资源修复和淡水生态系统保护具有重要意义,也是当前保护科学研究的热点[9—11]。目前, 柳江鱼类群落结构及其多样性研究报道较少, 制约了柳江渔业资源的管理和保护。本研究基于2018年鱼类及环境调查数据, 分析了柳江鱼类多样性现状, 探讨鱼类群落与环境因子的关系, 以期为柳江渔业资源修复和鱼类多样性保护提供参考依据。
1 材料与方法
1.1 研究地点
龙江是柳江最大的支流, 发源于贵州省三都县,至广西柳城县入柳江, 干流长367 km; 洛清江发源于广西临桂县, 在鹿寨县汇入柳江, 干流长275 km[1]。本研究在柳江干流的都柳江、融江、柳江江段及支流的洛清江、龙江共设置了9个调查站位, 分别为榕江、从江、融安、柳城、象州、永福、鹿寨、河池和宜州。榕江、从江、融安、柳城和象州位于干流, 河池和宜州位于龙江支流, 永福和鹿寨位于洛清江支流(图 1)。
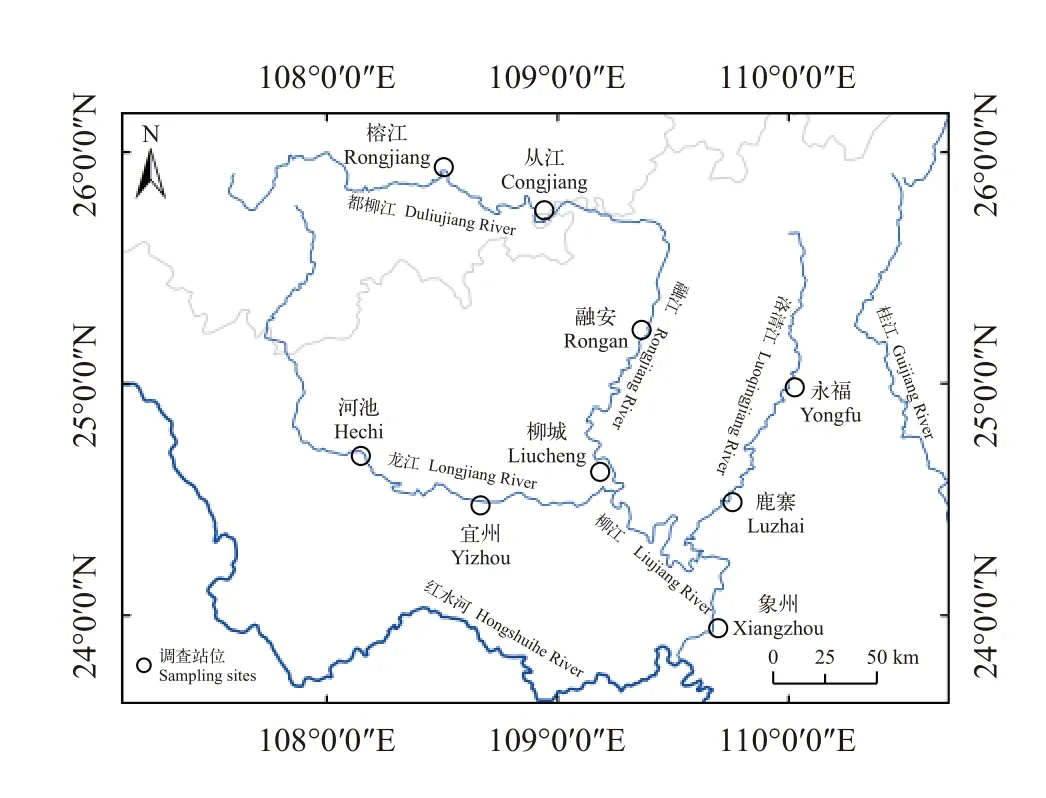
图1 柳江调查站位示意图Fig. 1 Map of sampling sites in the Liujiang River
1.2 采样方法
在各采样点渔船码头对渔民所有渔获物进行调查, 掌握鱼类种类组成, 调查的渔具主要为钩钓网、流刺网、定置刺网和虾笼。为了避免单一渔具捕获种类的局限性, 每种渔具类型渔船随机购买1—2船渔获物, 对采集到的样本进行分类, 并测量体长、体重, 进而分析鱼类群落结构状况。调查时间为2018年7月和12月, 分别代表夏季和冬季。鱼类种类鉴定参考《珠江鱼类志》[12]、《广西淡水鱼类志》[2]、《中国动物志》[13,14], 鱼类名录中分类和学名参考《中国内陆鱼类物种与分布》[15]中的鱼类名录信息。在采样站点使用便携式水质分析仪(YSI)、测距仪和流速仪进行环境调查, 测量指标包括水温(WT)、溶解氧(DO)、pH、溶解性总固体(TDS)、电导率(COD)、氧化还原电位(ORP)、盐度(SAL)、河宽(Width)、流速(Flow)和海拔(ALT)。
1.3 数据分析
鱼类物种多样性分析采用Shannon-Wiener多样性指数(H')、Margalef种类丰富度指数(Dm)、Pielou均匀度指数(J')、Simpson指数(Ds)[16—18], 计算公式如下:
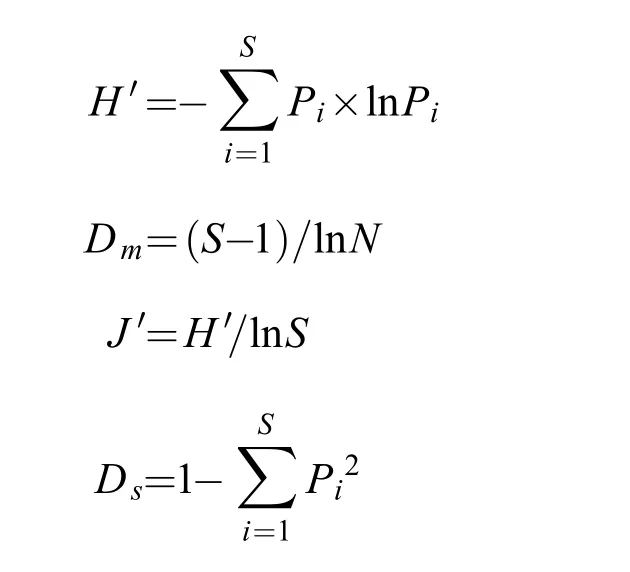
式中,S为鱼类种类数,N为总尾数,Pi为第i种鱼类所占的比例。
生物多样性指数分级评价参考国家环境保护标准(HJ442-2008), 根据H'指数范围将生物多样性水平划分为极贫乏(H'=0)、贫乏(H'<1)、一般(1≤H'<2)、较丰富(2≤H'<3)和丰富(H'≥3)5个级别[19]。
用Pinkas相对重要性指数(IRI)研究鱼类群落优势种, 计算公式如下[20]:
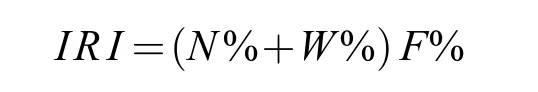
式中,N%为某一种鱼类的尾数占总尾数的百分比,W%为某一种鱼类的生物量占总生物量的百分比,F%为某一种鱼类出现的站位数占总站位数的百分比。本文选取IRI≥1000的鱼类为优势种。
丰度/生物量比较曲线(Abundance-Biomass Comparison Curve, 简称ABC曲线)最早是利用底栖生物群落结构的特征来监测海洋环境污染状况[21],近年来常用于鱼类群落结构受外界干扰的评价研究[16,22]。ABC曲线的统计量用W表示, 计算公式为:
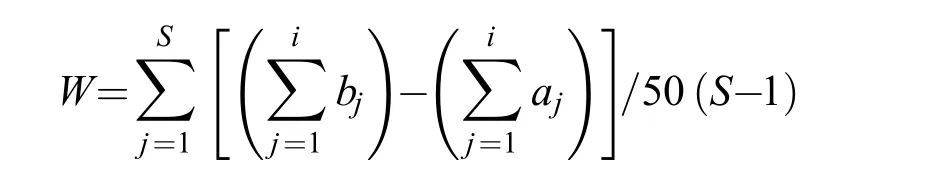
式中,S为出现的物种数,为物种的累计生物量,为物种的累计丰度。
在未受干扰的群落中,W为正值, 生物量优势度曲线在数量优势度曲线之上; 当群落受到中度干扰时,W值处于0附近, 两条曲线出现相交; 如果数量优势度曲线在生物量优势度曲线之上,W为负值,表明群落处于严重干扰状态。ABC曲线采用PRIMER 6.0软件完成。
鱼类群落时空分布特征利用非度量多维尺度分析(NMDS)进行分析[23], 物种数据进行lg(x+1)转化, 然后基于Bray-Curtis相似性度量。鱼类多样性与环境因子的关系采用典范对应分析(CCA)进行分析, 确定主要的环境影响因子[23]。为减小稀有物种的影响, CCA分析时将出现率小于5%的鱼类排除在外[24]。NMDS和CCA分析利用R统计软件完成。
2 结果
2.1 鱼类组成
柳江调查统计鱼类1711 kg, 36474尾, 隶属5目20科80属, 共115种。鲤形目80种, 鲇形目16种, 鲈形目14种, 合鳃鱼目3种, 鳗鲡目2种。鲤科68种, 鳅科9种, 鲿科8种, 鮨科5种, 平鳍鳅科3, 塘鳢科、鳗鲡科、鳢科、胡子鲇科、刺鳅科、斗鱼科和鲇科各2种, 虾虎鱼科、长臀科、钝头科、丽鱼科、鮰科、科、合鳃鱼科和沙塘鳢科各1种。在空间上, 各站位分别采集到鱼类物种数分别为: 榕江40种、从江44种、融安58种、柳城58种、象州46种、永福61种、鹿寨64种、河池56种和宜州54种。
在此次采集的鱼类中, 花鳗鲡Anguilla marmorata和斑鳠Hemibagrus guttatus为国家Ⅱ级保护野生动物[25]; 《中国濒危动物红皮书》[26]中将花鳗鲡和大眼卷口鱼Ptychidio macrops列为濒危, 长臀Cranoglanis bouderius列为易危; 《中国物种红色名录》[27]中将大眼卷口鱼和花鳗鲡列为濒危, 长臀和波纹鳜Siniperca undulata列为易危; IUCN红色名录(IUCN Red List)[28]中将卷口鱼Ptychidio jordani列为极危种, 日本鳗鲡Anguilla japonica列为濒危种, 南方拟䱗Pseudohemiculter dispar和长臀列为易危种, 南方白甲鱼Onychostoma gerlachi、鲢Hypophthalmichthys molitrix、鲮Cirrhinus molitorella、三角鲤Cyprinus multitaeniatus和波纹鳜列为近危种。
此次柳江调查新增记录鱼类19种: 红鳍原鲌Cultrichthys erythropterus、寡鳞飘鱼Pseudolaubuca engraulis、海南似鱎Toxabramis houdemeri、海南鲌Culter recurviceps、线细鳊Metzia lineata、胡Huigobio chenhsienensis、南方鳅鮀Gobiobotia meridionalis、虹彩光唇鱼、麦瑞加拉鲮Cirrhinus mrigala、拟平鳅Liniparhomaloptera disparis disparis、革胡子鲇Clarias gariepinus、斑点叉尾鮰Ictalurus punctatus、中间黄颡鱼Pelteobagrus intermedius、长吻Leiocassis longirostris、乌苏里拟鲿Pseudobagrus ussuriensis、鳜Siniperca chuatsi、尼罗罗非鱼Oreochromis niloticus、云斑尖塘鳢Oxyeleotris marmoratus、圆尾斗鱼Macropodus chinensis。其中, 麦瑞加拉鲮、革胡子鲇、斑点叉尾鮰、尼罗罗非鱼和云斑尖塘鳢为外来物种。
按照鱼类栖息环境和洄游方式, 柳江鱼类分为3种生态类型: 江海洄游性鱼类、江湖洄游性鱼类和定居性鱼类; 定居性鱼类占绝对优势, 占比85.22%。按照鱼类食性类型分为4种: 杂食性、肉食性、植食性和滤食性; 杂食性占比59.13%, 肉食性占比33.91%, 植食性占比5.22%。
2.2 鱼类多样性及优势种
Shannon-Wiener多样性指数、Simpson指数和Pielou均匀度指数均用鱼类个体数量(N)计算(图 2)。Shannon-Wiener指数为2.13—3.12, Simpson指数为0.81—0.93, Pielou均匀度指数为0.58—0.80。Margalef种类丰富度指数为4.65—7.18。根据Shannon-Wiener指数可知, 各河段鱼类多样性总体处于较丰富的级别, 鱼类丰富度较高, 个体分布较均匀。

图2 各站点鱼类多样性指数Fig. 2 Fish biodiversity index of each station
根据IRI值的大小排序,IRI≥1000的鱼类为优势种类。结果显示, 柳江鱼类优势种时空分布存在差异(表 1)。榕江夏季8种, 冬季6种, 相同种类为银Squalidus argentatus和大眼华鳊Sinibrama macrops。从江夏季7种, 冬季5种, 相同种类为鲤Cyprinus carpio和大眼华鳊。融安夏季和冬季各6种,相同种类为蛇Saurogobio dabryi和南方拟䱗。柳城夏季5种, 冬季8种, 相同种类为银、南方拟䱗和鳙。象州夏季和冬季各6种, 相同种类为粗唇Pseudobagrus crassilabris和斑鳠Hemibagrus guttatus。永福夏季5种, 冬季8种, 相同种类为鲤。鹿寨夏季6种, 冬季8种, 相同种类为银。河池夏季9种, 冬季4种, 相同种类为银和银鲴Xenocypris macrolepis。宜州夏季和冬季各6种, 相同种类为银。各调查站点共有的优势种类为银和鲤。

表1 柳江鱼类优势种的时空分布Tab. 1 Spatial and temporal distribution of dominant fish species in Liujiang River
2.3 丰度/生物量比较曲线
丰度/生物量比较曲线显示, 宜州、榕江、鹿寨和柳城的W值均小于0, 丰度优势度曲线总体在生物量优势度曲线上方, 且丰度优势度曲线的起点要高于生物量优势度曲线的起点, 表明这4个站点鱼类群落受到严重干扰。象州和融安的W值大于0,且生物量优势度曲线与丰度优势度曲线相交, 表明这2个站点鱼类群落受到中度干扰。河池、从江和永福的W值大于0, 生物量优势度曲线位于丰度优势度曲线之上, 生物量优势度曲线的起点高于丰度优势度曲线的起点, 表明这3个站点鱼类群落结构相对稳定(图 3)。
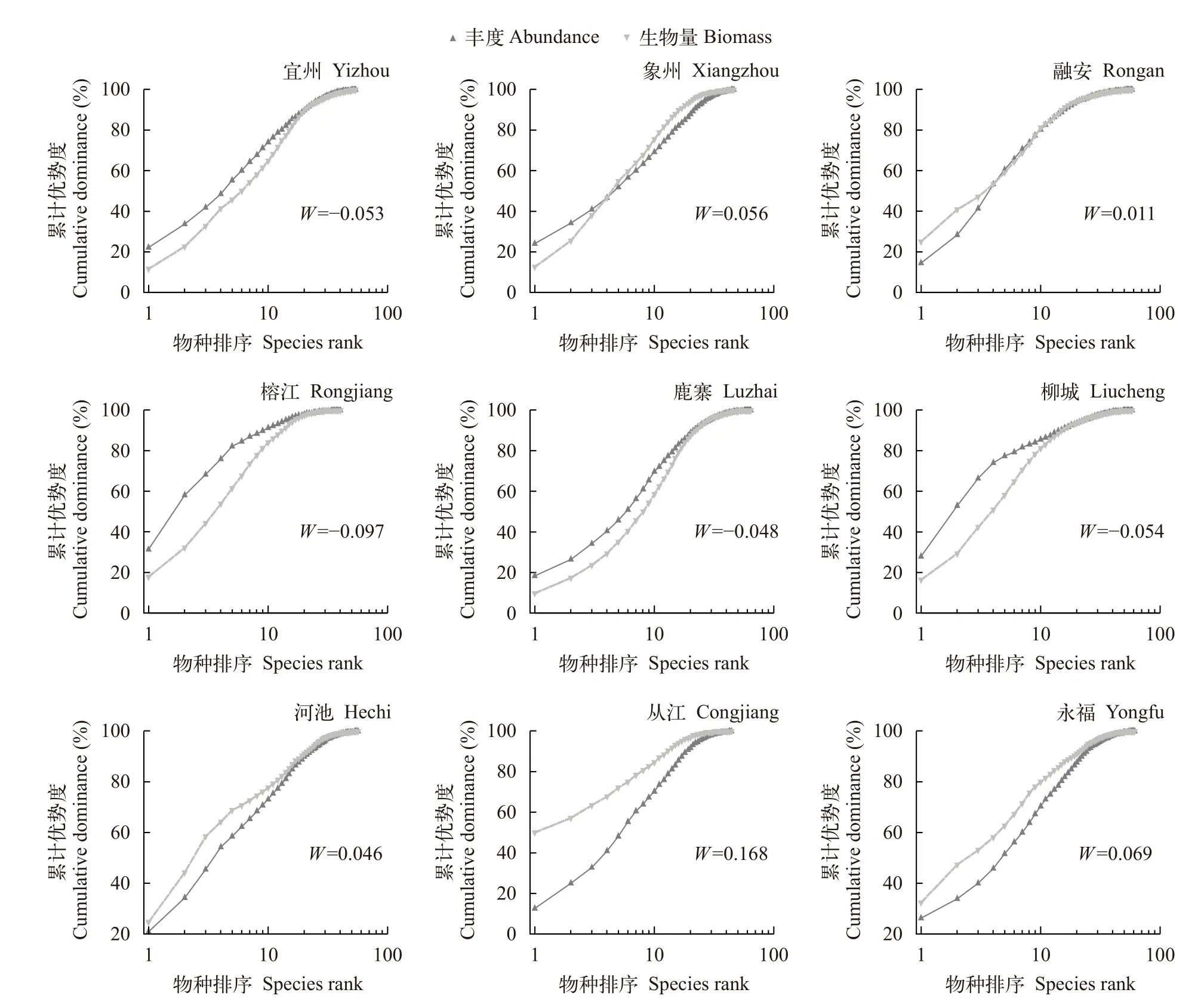
图3 各站点鱼类群落的丰度/生物量曲线Fig. 3 Abundance/biomass curve of fish communities at each station
2.4 鱼类群落时空变化
采用Bray-Curtis相似性的非度量多维尺度排序方法分析, 结果表明柳江鱼类群落结构表现为空间自相关, 地理空间相近的站点聚为一类。位于柳江上游的从江和榕江为一类, 以宽鳍鱲、大鳍鳠、大眼华鳊和胡等为主; 支流洛清江的永福和鹿寨为一类, 以圆吻鲴、海南拟䱗、棒花鱼、带半刺光唇鱼和麦穗鱼等为主; 位于柳江中下游的融安、柳城、象州和龙江支流的河池和宜州为一类, 以银鲴、南方拟䱗、䱗、鳙、鲢、鲮和尼罗罗非鱼等为主。各采样点鱼类群落组成的季节性组成变化不明显(图 4)。
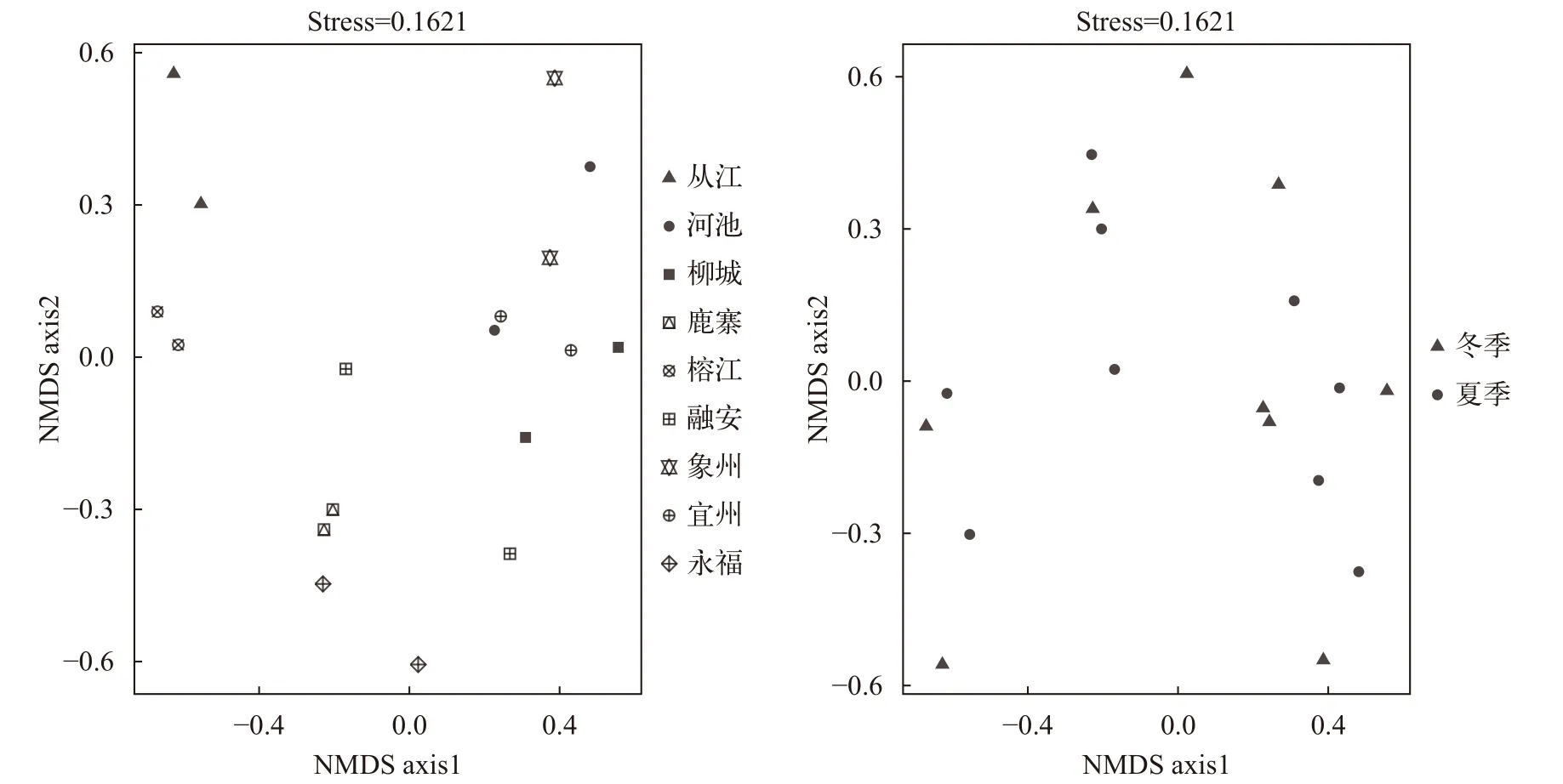
图4 鱼类群落时空变化的NMDS排序Fig. 4 NMDS ordination of spatiotemporal variation of fish community
2.5 鱼类分布与环境因子的关系
本研究监测了10个环境因子: 水温(WT)、溶解氧(DO)、pH、溶解性总固体(TDS)、电导率(COD)、氧化还原电位(ORP)、盐度(SAL)、河宽(Width)、流速(Flow)和海拔(ALT)。鱼类群落结构差异与环境因子关系的CCA分析结果显示, 10个环境因子对鱼类分布的总解释量为67.74%, 其中CCA轴1和CCA轴2对鱼类分布的解释量分别为16.80%和10.64%, 环境因子Width和ALT主要贡献于轴1, WT、DO、pH、COD和ORP主要贡献于轴2(图 5)。随机置换检验结果表明, Width、ALT和pH是柳江鱼类群落结构差异的最主要影响因子(P<0.05; 表 2)。其中, 鲮、鲢、赤眼鳟等种类偏好相对较宽的河段, 黄尾鲴Xenocypris davidi、北江光唇鱼Acrossocheilus beijiangensis等种类偏好海拔相对较低的河段, 乌苏里拟鲿Pelteobagrus ussuriensis、麦穗鱼、越南鱊Acheilognathus tonkinensis、中华花鳅Cobitis sinensis、带半刺光唇鱼等偏好pH相对较低的河段。
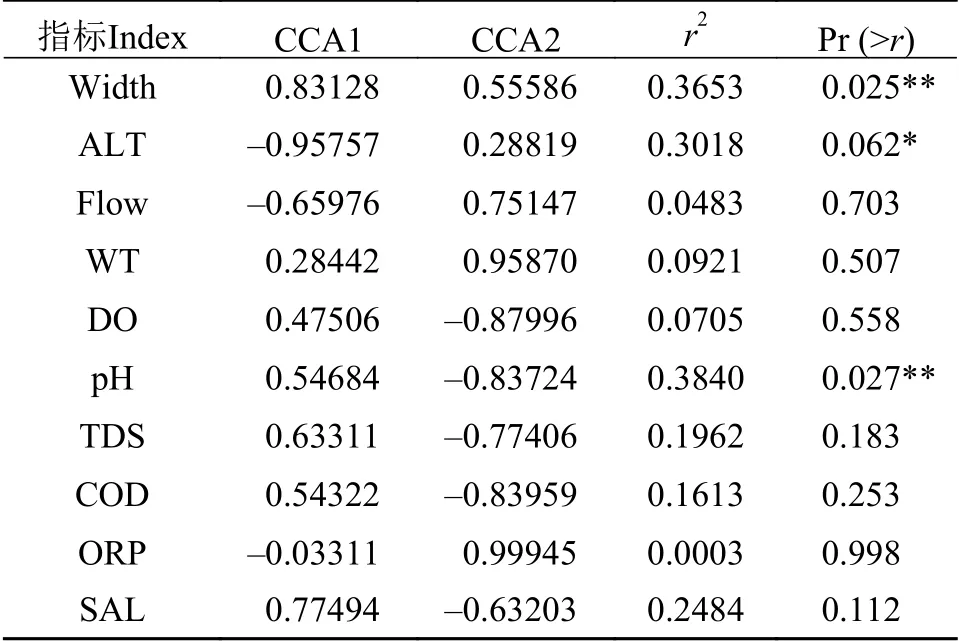
表2 排序轴与环境因子的相关系数Tab. 2 Correlation coefficient between the sorting axis and environmental factors
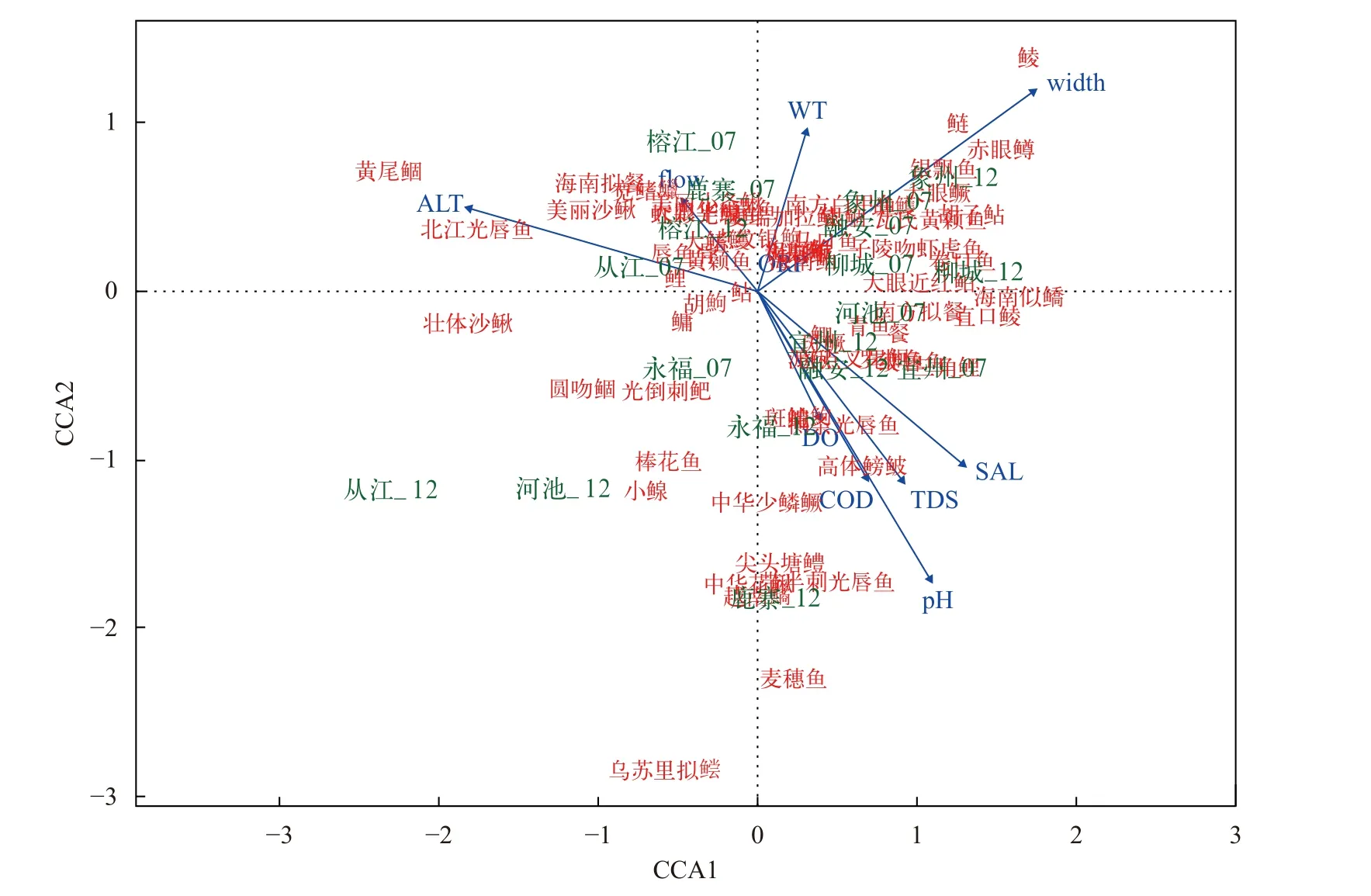
图5 鱼类分布与环境因子关系的CCA分析排序双序图Fig. 5 Double sequence diagram of CCA analysis on the relationship between fish distribution and environmental factors
3 讨论
3.1 鱼类生物多样性及其变化
与历史资料相比, 柳江鱼类种类组成发生了较大变化。《广西淡水鱼类志》[2]记载鱼类141种, 朱瑜等[3]报道鱼类137种, 本次调查采集鱼类115种, 有20多种鱼类未采集到, 新增记录鱼类19种。历史资料记载的赤魟Dasyatis akajei、中华鲟Acipenser sinensis、鳤Ochetobibus elongatus等[2]种类此次调查均未采集到。历史资料记载象州石龙三江口曾是中华鲟的主要产卵场, 现已多年未见其踪迹; 鳤曾经是西江重要经济鱼类, 现今其资源量衰退严重,种群已极度濒危[29]。采集濒危物种花鳗鲡1尾和大眼卷口鱼16尾, 易危物种长臀2尾, 表明这些种类在柳江仍处于濒危状态。历史上柳江常见经济鱼类有50多种, 主要有青鱼、草鱼、赤眼鳟、大眼近红鲌、翘嘴鲌Culter alburnus、鳊Parabramis pekinensis、细鳞鲴Xenocypris microlepis、圆吻鲴、
鳙、鲢、花䱻Hemibarbus maculatus、蛇、光倒刺鲃Spinibarbus hollandi、倒刺鲃Spinibarbus denticulatus等[3]。然而, 当前渔业资源状况形势严峻, 重要经济鱼类种类减少, 资源衰退严重。此次调查常见的种类为银、鲤、大眼华鳊、胡、银鲴、南方拟䱗, 大部分为定居性、杂食性的小型鱼类。本次调查新增鱼类记录中, 海南似鱎、红鳍原鲌和胡等在珠江流域有广泛的分布, 《广西淡水鱼类志》[2]记载柳江有拟鲿属鱼类分布, 本次采集的乌苏里拟鲿依据《中国动物志》[14]鉴定分类,乌苏里拟鲿从珠江至黑龙江水系均有分布[15]。相较于柳江历史调查资料, 此次新增近二十种鱼类,说明柳江已有的鱼类研究相对较少, 调查范围和强度较小, 未能充分体现柳江鱼类的多样性。外来水生生物入侵也对柳江鱼类多样性造成威胁, 本次调查采集的麦瑞加拉鲮、革胡子鲇、斑点叉尾鮰、尼罗罗非鱼和云斑尖塘鳢属于外来物种, 并且尼罗罗非鱼在柳城、鹿寨和宜州已成为优势种。生物入侵是全球变化的一部分, 外来入侵生物通过与本地生物竞争生态位, 对本地生物群落和生态系统产生严重影响, 甚至导致本地种群的灭绝[30,31], 柳江鱼类资源的保护和管理也要关注生物入侵的威胁。根据Magurran[32]提出的多样性指数一般范围(1.5—3.5), 柳江鱼类Shannon-Wiener多样性指数为2.13—3.12, 鱼类多样性处于一般至较丰富水平。与珠江流域其他江段相比[33—35], 柳江鱼类多样性水平相对较高。然而, 柳江鱼类群落结构及其多样性也受到不同程度的干扰。其中, 宜州、榕江、鹿寨和柳城江段鱼类群落受到严重干扰; 象州和融安江段处于中度干扰状态。柳江大型经济鱼类占比减少, 鱼类优势种向小型化转变, 并显示低龄化趋势,整体渔业资源水平呈下降趋势。水坝建设、水质污染和过度捕捞是造成柳江渔业资源衰退的主要原因。柳江干流规划了11级电站, 龙江已建7个水
电站, 洛清江已建5座水库[1]。水电梯级开发对河流造成阻隔作用, 改变水文情势, 原有连续的河流生态系统被片段化为水库-河流生态系统, 库区和坝下水文情势差异巨大。对洄游性鱼类产生阻隔, 鱼类生殖和索饵等生活史过程受到影响, 同时也影响鱼类的种群间交流。此外, 珍稀濒危特有鱼类适应柳江原有特殊生境, 水坝建设对原有特殊生境破坏大, 加剧了其濒危程度。陈锋等[6]研究表明, 珠江梯级开发对鱼类造成的主要影响表现为: 洄游性鱼类大幅下降, 缓流和静水鱼类大幅增加, 产黏沉卵鱼类显著下降, 外来物种增加, 珍稀特有鱼类濒危程度加剧等。柳江总体水质较好, 但局部地区仍然受到工业污染的威胁, 对渔业资源产生严重影响。有些江段捞石、采砂活动频繁, 破坏了鱼类栖息地,对鱼类直接造成损害。过度捕捞严重威胁淡水渔业资源和鱼类多样性[36], 目前柳江仍存在电鱼、毒鱼和炸鱼等非法捕捞行为, 对鱼类种群造成巨大危害。
3.2 鱼类群落空间变化及其与环境因子的关系
鱼类群落的时空分布受环境因子影响, 河流中不同区域鱼类群落组成因栖息地类型不同而表现出差异[37—40]。柳江鱼类群落组成表现为空间自相关, 大体划分为“从江-榕江”“永福-鹿寨”和“河池-宜州-柳城-象州-融安”3个类群。在地理位置上, 从江和榕江位于柳江干流上游都柳江段, 柳城、融安和象州位于柳江干流下游融江和柳江段, 河池和宜州位于支流龙江, 鹿寨和永福位于支流洛清江。柳江各江段鱼类群落组成空间变化与地理位置基本一致, 而支流龙江与干流融江段鱼类组成较为相似。鱼类群落分布受多种因子的影响, 已有的研究表明距源距离、河流宽度、pH、水深、水温、溶解氧、浑浊度、河床类型和坡度等是影响鱼类群落分布的主要因子[37,41,42]。本研究发现河流宽度、海拔和pH是柳江鱼类群落分布差异的主要影响因子。作为影响鱼类分布的主要因子, 河流宽度和深度的比值用于河流鱼类生境评价, 通常比值越大物种的丰富度越大, 意味着鱼类栖息地更优良; 相反,较小的比值表明鱼类栖息地较差[43]。Rodrigues-Filho等[44]研究认为, 河流鱼类多样性受pH和河流宽度影响, 在某种程度上, 具有较高酸度值和较宽河道的河流生物多样性水平较低。Dangles等[45]的研究中也观察到水生系统的自然酸性导致生物多样性丰富度的降低。本研究结果与此相似, pH较高的永福(8.23)、鹿寨(8.99)、河池(8.79)和宜州(8.5)站点鱼类丰富度相对较高, pH较低的榕江(7.79)和从江(7.69)站点鱼类丰富度较低。pH是影响鱼类物种丰富度的主要原因之一, 柳江属于典型的喀斯特地貌地区, 岩性较为复杂, 以碳酸岩石为主; 水化学组成主要由岩石风化作用所控制, 以碳酸盐岩贡献为主, 不同的下伏岩层分布决定了相应江段控制作用的强弱, 从而决定了流域水化学组成的空间差异性[1,46]。本研究中鱼类物种丰富度随海拔增加而呈现下降趋势, 符合Rapoport’s法则[47]; 其他流域鱼类物种研究也表明, 淡水鱼类的多样性随着海拔升高而逐渐降低[48,49]。
3.3 保护管理措施建议
珠江是我国重要的水源地和水生生物宝库, 为珍稀濒危物种和重要水生经济物种提供了良好生存条件和繁衍空间[50], 柳江作为珠江的重要支流,研究其鱼类多样性对流域渔业资源修复和水生态系统保护具有重要作用。由于水坝建设、采砂、过度捕捞等因素影响, 柳江鱼类资源呈现衰退趋势,主要体现在经济鱼类占比减少、鱼类小型化和低龄化。因此, 建议在以下几个方面加强鱼类多样性保护: (1)水产种质资源保护区是鱼类资源保护的一种有效形式, 在渔业保护中发挥重要作用[51]。目前柳江已有“柳江长臀桂华鲮赤魟国家级水产种质资源保护区”, 建议进一步根据柳江独特的生境特征, 以珍稀濒危特有鱼类为保护对象, 在关键生境区域划定保护区生态红线, 遏制鱼类生境的丧失和退化。(2)渔业监管部门应严格执行渔业法和禁渔期制度的规定, 加大执法力度, 严厉打击偷捕、电捕行为, 杜绝非法捕捞。(3)针对柳江重要经济鱼类, 例如四大家鱼, 赤眼鳟、翘嘴鲌、光倒刺鲃、倒刺鲃等开展增殖放流, 并进行增殖放流效果评估。(4)开展鱼类洄游通道与产卵场修复研究, 推进河流连通、水利调控和产卵场生态修复。在已有梯级水电站加建过鱼通道, 或者补建集运鱼系统促进鱼类种群交流。根据鱼类繁殖需要, 进行水文生态调度。(5)对于珍稀濒危特有鱼类, 例如花鳗鲡、大眼卷口鱼、长臀、南方白甲鱼、三角鲤等建立鱼类保育中心, 同时开展流域珍稀特有鱼类的科研、监测、救护和繁育工作。

附表 1 柳江鱼类调查名录Attached table 1 Fish species in the Liujiang River
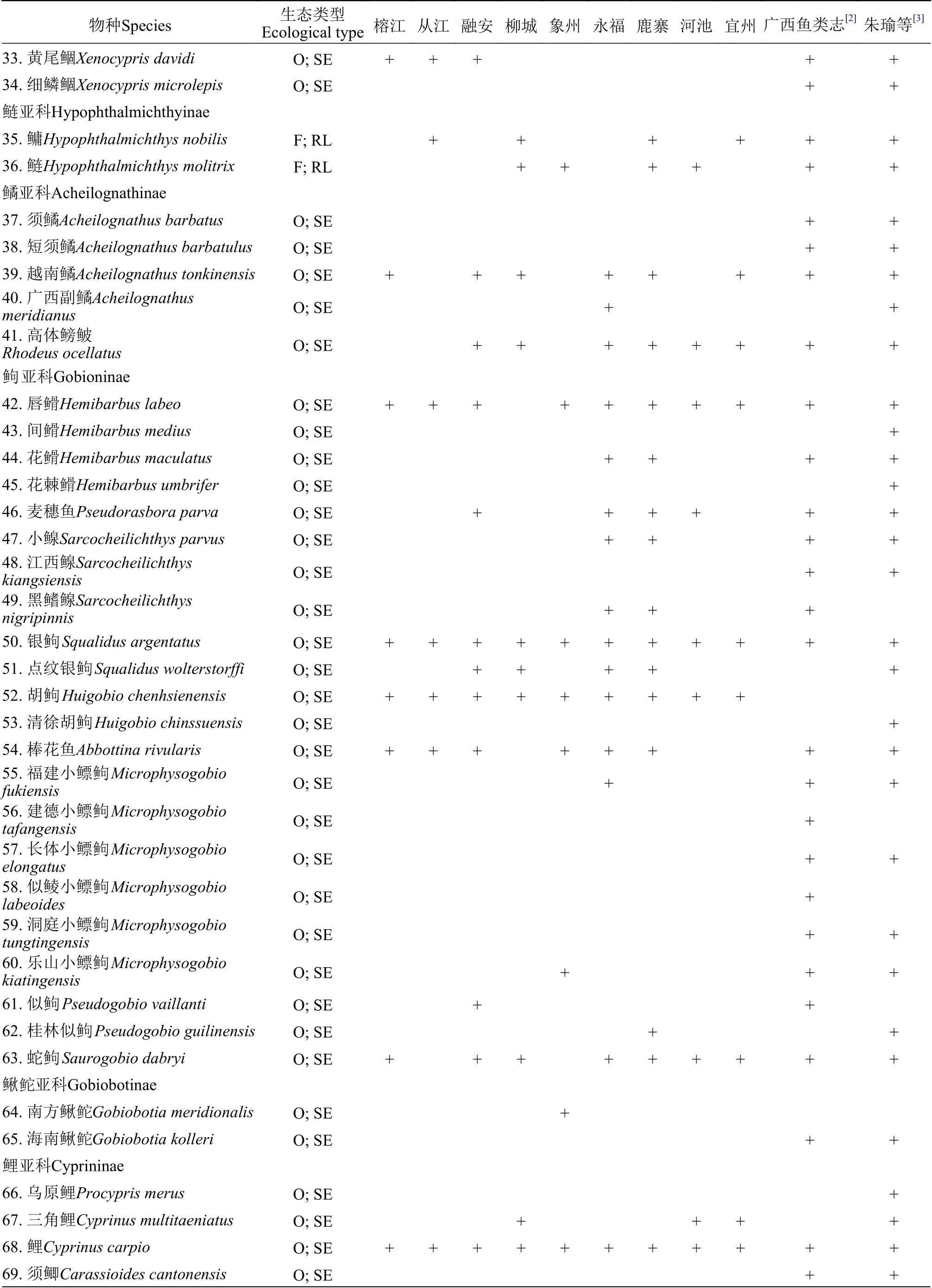
续表1
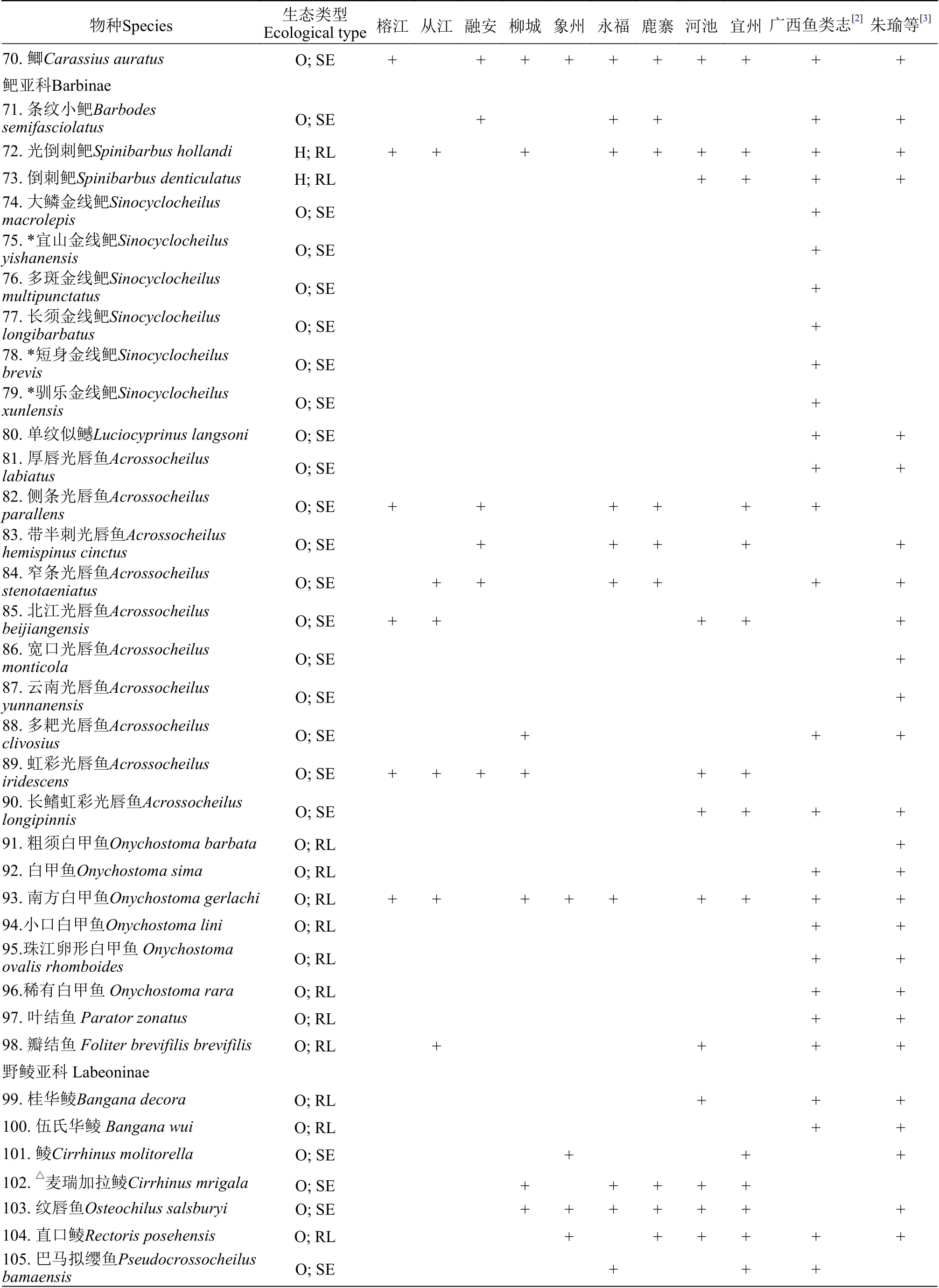
续表1

续表1
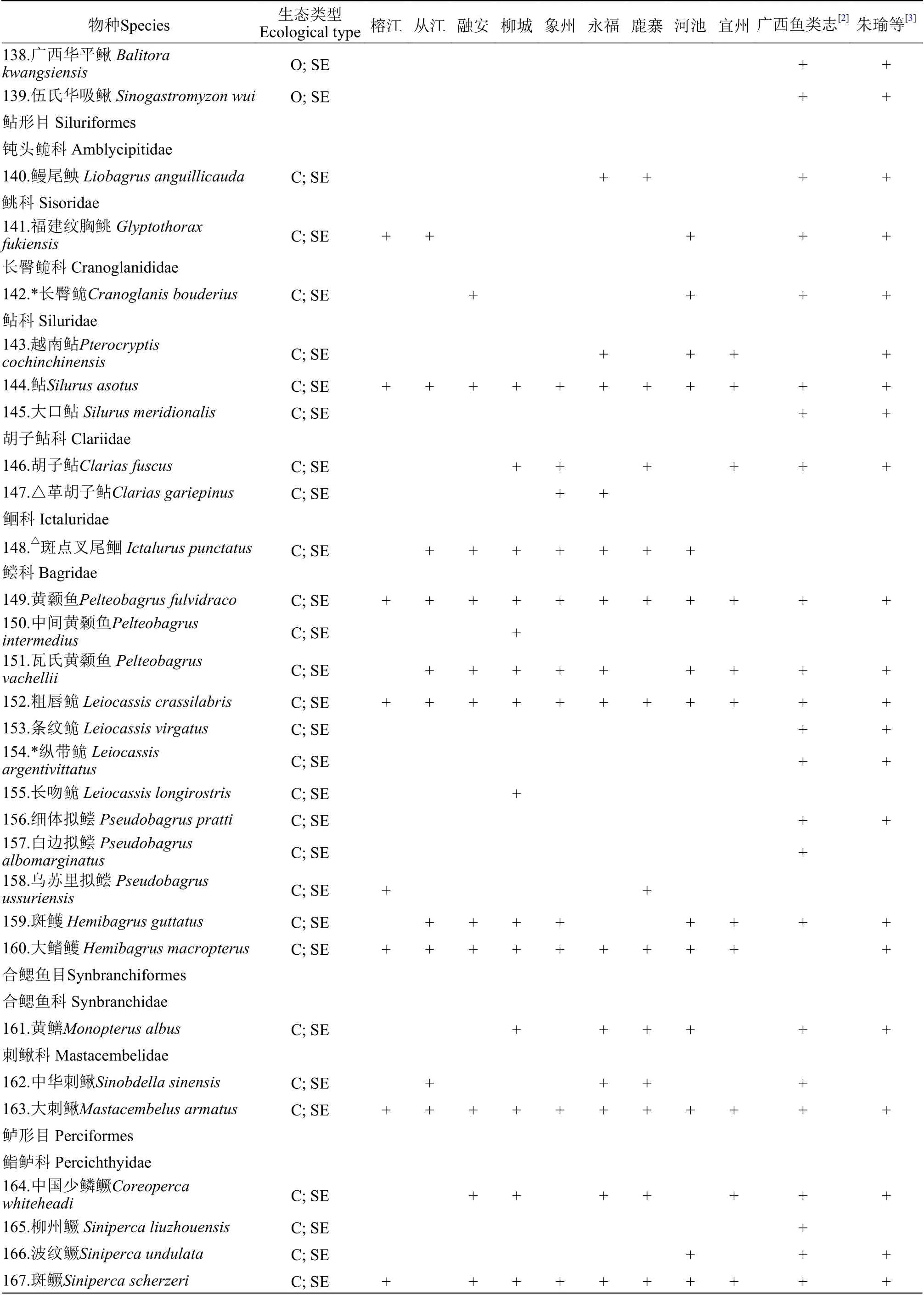
续表1
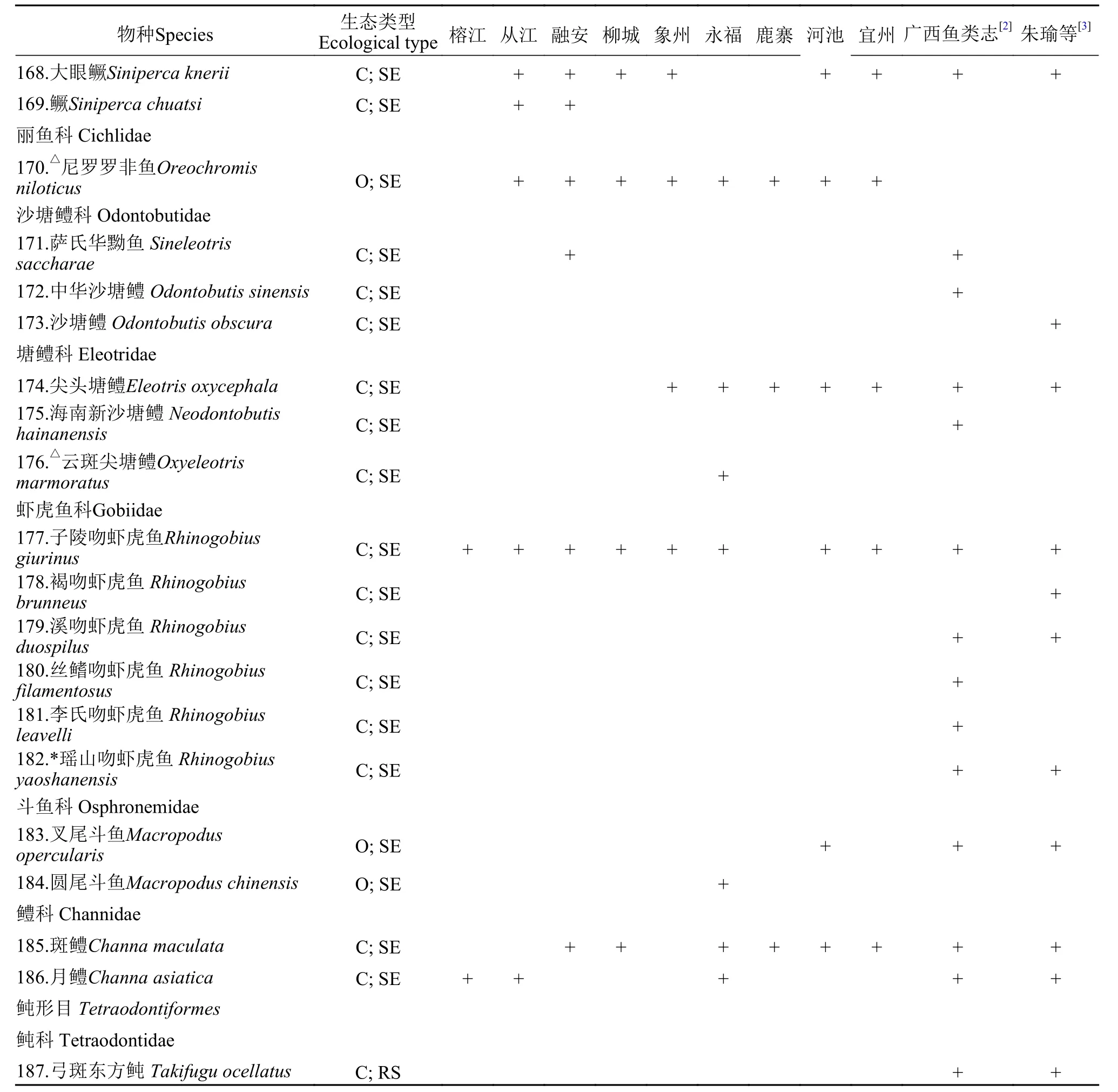
续表1
