高营养级捕食者在浅水湖泊沿岸带与敞水区能量耦合的维持作用
2022-04-13祝孔豪郭钰伦
徐 瑛 祝孔豪 郭钰伦 王 康 王 韬
(1. 新余市生态环境局, 新余 338000; 2. 中国科学院水生生物研究所, 武汉 430072)
水域生态系统的捕食者, 能够通过营养级联效应调节低营养级物种的组成和丰度, 影响水生生物群落的结构和多样性, 对于维持食物网的能量流动和生态系统的稳定性发挥着重要作用。捕食者除通过捕食影响猎物的丰度和分布外, 还可通过间接途径影响其他非猎物类群和生态系统过程[1,2]。已有越来越多的研究表明, 捕食者不仅可以通过下行效应对生产者有机物质生产过程产生直接作用, 还可调节生态系统环境因素对低营养级生物造成的影响[3,4], 在水域生态系统中具有重要的生态地位和不可替代的功能作用[5]。在近几十年来, 人类活动的影响已造成了许多捕食者的灭绝, 从而造成了生态系统一系列难以预料的变化, 生态学家对于捕食者的研究更加关注[5,6]。
湖泊是涵盖多种生境条件的复杂生态系统。湖泊不同区域生境条件往往不同, 环境因子与饵料生物也存在差异, 进而可能导致捕食者群落特性在同一湖泊中的空间异质性[7]。有研究表明, 捕食者由于活动能力强、活动范围大, 在不同生境间食物网能量流动与物质循环中发挥了重要作用[8]。这种不同生境间的耦合对于食物网动力学至关重要[9]。在湖泊生态系统中, 沿岸带生境围绕固体湖岸基质,一般有大量水生植物及附着藻类群落, 敞水区生境则为悬浮状态, 二者差异较大[10], 但关于二者之间食物网如何紧密联系实现物质与能量的传递与流动的研究尚不多见。不同生境间的耦合是一个重要的生态系统过程, 它通过化学、物理及生物过程的作用促进营养和能量的流动, 进而将不同的生境连接起来[9]。捕食者被认为在生境耦合的生态系统过程中发挥了重要作用[9,11]。
稳定同位素技术是研究生态系统食物网结构和营养关系及其动态变化的重要手段, 基于稳定同位素质量平衡模型, 其可以用于消费者营养溯源,以确定多种营养来源对消费者营养的贡献比重[12,13]。碳稳定同位素比值(δ13C)在捕食者和食物之间的分馏较小(0—1‰), 可用于指示食物来源; 氮稳定同位素比值(δ15N)在捕食者与食物之间存在明显的富集效应 (3‰—4‰), 多用于指示消费者的营养等级[12,14]。稳定同位素技术与胃肠含物分析等技术手段相比,其能反映较长时间尺度内捕食者营养来源特征[15,16]。因此, 可利用碳氮稳定同位素在食物网中的特性来指示湖泊中捕食者的食物来源与营养级, 进而探究捕食者在湖泊不同栖息地物质与能量流动中的作用。
1 材料与方法
1.1 研究区域概况
保安湖(30°12′—30°18′N, 114°39′—114°46′E)位于湖北省大冶市西北部, 地处长江中游江汉平原东部边缘, 原与三山湖连成一片, 现以北练山、南练山为界, 山以东称为三山湖, 以西称为保安湖, 属梁子湖水系, 为长江中游南岸的一个轻度—中度富营养型湖泊。
1.2 样品采集与测定
达氏鲌(Culter dabryi)与红鳍原鲌(Cultrichthys erythropterus)均属于鲤形目(Cypriniformes), 鲤科(Cyprinidae), 鲌亚科(Culterinae), 达氏鲌为鲌属(Culter), 红鳍原鲌为原鲌属(Culterichthys)。鲌类作为湖泊和水库的高营养级消费者, 对水生生物组分具有重要的调控功能。
本研究样品于2019年6月在保安湖采集, 通过在沿岸带和敞水区设置网具, 获取饵料鱼类样品和高营养级捕食者—鲌类(达氏鲌与红鳍原鲌)样品。在样品采集过程中, 研究人员采集到达氏鲌10条(沿岸带与敞水区各5条), 红鳍原鲌采集到7条(沿岸带5条, 敞水区2条); 饵料鱼类种类主要包括湖鲚(Coilia ectenes taihuensis)、黄颡鱼(Pelteobagrus fulvidraco)、圆尾鲴(Distoechodon tumirostris)、似鱎(Toxabramis swinhonis)、䱗(Hemiculter leucisculus)、似刺鳊(Paracanthobrama guichenoti)和中华鳑鲏(Rhodeus sinensis Günther)等, 体长范围分别为湖鲚(193±38) mm, 黄颡鱼(177±37) mm, 圆尾鲴(125±12) mm, 似鱎(100±9) mm, 䱗(109±27) mm, 似刺鳊(202±7) mm, 中华鳑鲏(43±8) mm。鱼类样品放入干净白瓷盘后, 利用干净的镊子及手术剪取适量背部白肌肉, 去除骨刺与鱼皮, 然后放入冻存管密封冷冻保存, 一尾样品取样完成后即用超纯水清洗取样器材, 避免污染。蚌和螺等大型底栖动物样品通过带网夹泥器获取, 冲洗干净后, 利用干净镊子去壳取肌肉组织, 放入冻存管中密封冷冻保存。其余底栖动物样品使用彼德森采泥器采集, 底栖动物进行种类鉴定后, 分类分别放置于蒸馏水中过夜, 让其排空消化道内残留物。为保证足够的样品分析质量, 将同一物种的底栖动物混合在一起进行分析。浮游动物样品使用13#浮游生物网收集,并将收集到的浮游动物样品过滤至灼烧过的GF/C滤膜。将以上采集到的样品放入烘箱中60℃烘干至恒重。浮游动物样品使用手术刀片轻轻从滤膜上刮取下来。所有同位素样品均利用珠磨式组织研磨器(MiniBeadbeater-16)研磨粉碎至粉末状,放入干燥器中干燥保存待测。
利用稳定同位素比质谱仪(CE公司的Carlo-Erba NC 2500元素分析仪与Thermo fisher公司的Delta Plus质谱仪联用)测定保安湖高营养级捕食者(达氏鲌与红鳍原鲌)以及敞水区和沿岸带饵料生物的碳、氮稳定同位素值δ13C和δ15N。分析碳(δ13C)、氮(δ15N)同位素的参照物质分别是VPDB(Pee Dee Belemnite)和空气中N2, 标准物质分别选用国际上通用的IAEA-USGS24和IAEA-USGS26。
1.3 营养富集因子与源矫正
营养富集因子(Trophic enrichment factor, TEF,Δ)由消化与代谢过程中同位素分馏引起, 其定义为Δ=δtissue–δdiet, 其所代表的捕食者组织间与其营养来源间的同位素的差异是构建贝叶斯混合模型的一个重要前提条件[17—19]。TEF的取值受到生物种类、取样组织以及食物来源等多种因素的影响[20],对于TEF的错误选择和应用会造成模型结果的潜在问题[18,21]。捕食者稳定同位素数据落入经TEF矫正后营养来源确定的同位素混合空间中, 对于模型得出精确结果十分重要[22]。为使稳定同位素质量平衡混合模型得出准确的结果,应当合理进行TEF的选择和源矫正[18]。
根据达氏鲌与红鳍原鲌食性, 将其食物来源进行整合, 形成浮游动物(Zooplankton)、底栖动物(Zoobenthos)、敞水区饵料鱼类(Pelagic Prey Fish)和沿岸带饵料鱼类(Littoral Prey Fish)4类营养来源用于模型构建。本研究中达氏鲌与红鳍原鲌为保安湖高营养级鱼类捕食者, 使用Δδ15N=(3.40±0.50)‰和Δδ13C=(0.80±1.20)‰对4类营养来源(敞水区饵料鱼类、沿岸带饵料鱼类、浮游动物和底栖动物)的碳氮稳定同位素数据值进行矫正。为反映数据误差的整体特征, 通过统计平方公差法(Root-Sum-Squares Error, R SSE将TEF的标准误差整合到4类营养来源的稳定同位素误差中[23]。
保安湖高营养级捕食者达氏鲌与红鳍原鲌稳定同位素数据落在4种营养来源确定的同位素混合空间中, 且共线性特征不显著(图 1), 适合对达氏鲌与红鳍原鲌的食物来源组成进行进一步的分析。
1.4 建模数据质量检验
首先, 通过图 1TEF校正后的营养来源稳定同位素混合空间, 可以初步判断达氏鲌与红鳍原鲌稳定性同位素数据是否落入4种营养来源确定的同位素混合空间中。其次, 通过迭代模拟的方法来判别达氏鲌与红鳍原鲌稳定同位素数据落入4种营养来源确定的同位素混合空间中的可能性。迭代模拟即基于TEF校正后的4种营养来源的稳定同位素均值和标准偏差, 迭代生成10000次稳定同位素混合空间, 继而计算达氏鲌与红鳍原鲌稳定同位素数据落入这10000次混合空间中的频次。 达氏鲌与红鳍原鲌稳定同位素数据落在>0.05可能性区域, 可判定数据质量满足建模需要(图 2A—C)。
当捕食者样本的稳定同位素数据过多的存在于由各种营养来源确立的稳定同位素混合空间的质心区域时, 同位素混合空间中营养来源与捕食者的共线性会增加, 导致稳定同位素质量平衡模型求解无法收敛或无法较好的匹配预测值与观测值[24]。因此, 继续通过迭代模拟的方法来判别数据质量。再次基于TEF校正后的4种营养来源的稳定同位素均值和标准偏差, 迭代生成10000次高风险混合同位素空间(以质心为中心的50%不规则多边形面积内)[24,25], 继而计算达氏鲌与红鳍原鲌稳定同位素数据落入高风险混合空间中的次数, 计算落入高风险稳定同位素混合空间的概率, 来检验数据建模的质量; 达氏鲌与红鳍原鲌稳定同位素数据落在<0.95可能性区域, 可判定数据质量满足要求(图 2D—F)[24,25]。
图2A—C显示了保安湖捕食者达氏鲌与红鳍原鲌稳定同位素值的变化将如何影响营养来源混合模型合理求解的概率[25]。达氏鲌与红鳍原鲌的稳定同位素数据值应处于95%概率轮廓内, 本研究中的两种鲌类样品均符合要求。图 2D—F则显示了捕食者达氏鲌与红鳍原鲌同位素值的变化将如何影响营养来源混合模型低估风险的概率[24]。捕食者达氏鲌与红鳍原鲌样品落入风险区的概率总体低于50%, 未出现高于95%概率的样本, 同样满足模型要求。
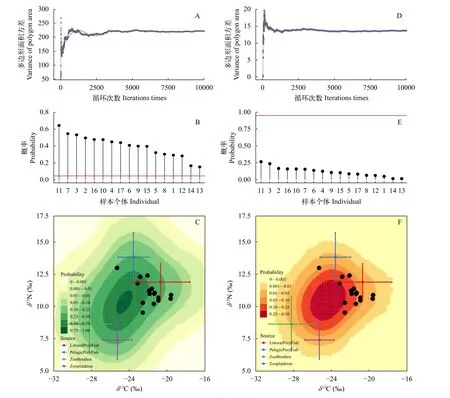
图2 混合多边形迭代布局Fig. 2 Mixing polygon simulation
1.5 模型构建与非度量多维尺度分析(NMDS)
本研究使用R包simmr来拟合所有的同位素贝叶斯混合模型(iter=50000, burn=1000, thin=10,n.chain=4), 使用R包vegan对两种栖息地鲌类食物来源数据进行NMDS分析。
2 结果
2.1 模型结果
模型结果表明(表 1), 沿岸带和敞水区栖息地采集到的达氏鲌与红鳍原鲌样品营养来源组成基本相似, 均以浮游动物占比最高, 以沿岸带饵料鱼类占比其次。
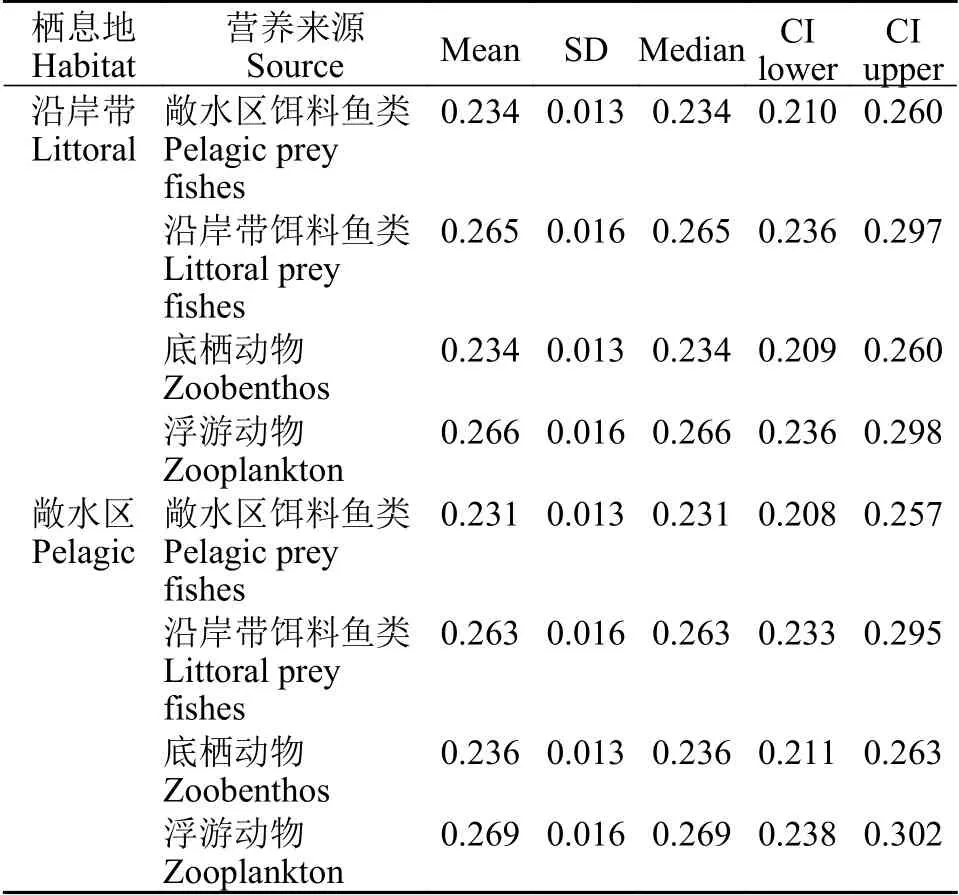
表1 不同营养来源贡献比例Tab. 1 Contribution ratio of different nutrition sources
2.2 模型整体性能评价
在贝叶斯模型中, 可以用偏差信息量准则(Deviance information criterion, DIC), 即等级模型化的赤池信息量准则(Akaike information criterion,AIC)[26], 来比较模型的相对节俭性(Parsimony)[27]。DIC值越小表明贝叶斯模型拟合的越好[28]。本研究中,由于来自沿岸带与敞水区的达氏鲌和红鳍原鲌观测值样本数量并不相同, 因此使用校准的DIC(DICcor,进行比较[29,30]。其中N表示每个模型的样本数,n为比较的模型中样本数最低的数据。本研究中计算DICcor时,n取值为7。模型整体性能评价结果表明(表 2), 沿岸带与敞水区两种生境鲌类营养来源贝叶斯混合模型拟合较好。两种栖息地的模型比较来看, 敞水区较沿岸带DIC值更低, 但DICcor值较沿岸带略高。
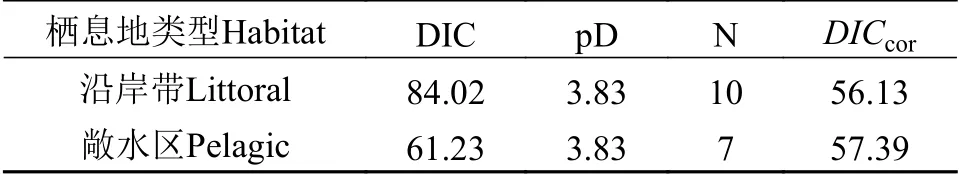
表2 模型整体性能评价Tab. 2 Model performance
2.3 两种栖息地鲌类的非度量多维尺度分析
对两种栖息地鲌类食物来源数据进行NMDS分析(图 3), 来自两种栖息地的鲌类食物来源基本一致, 未出现差异。
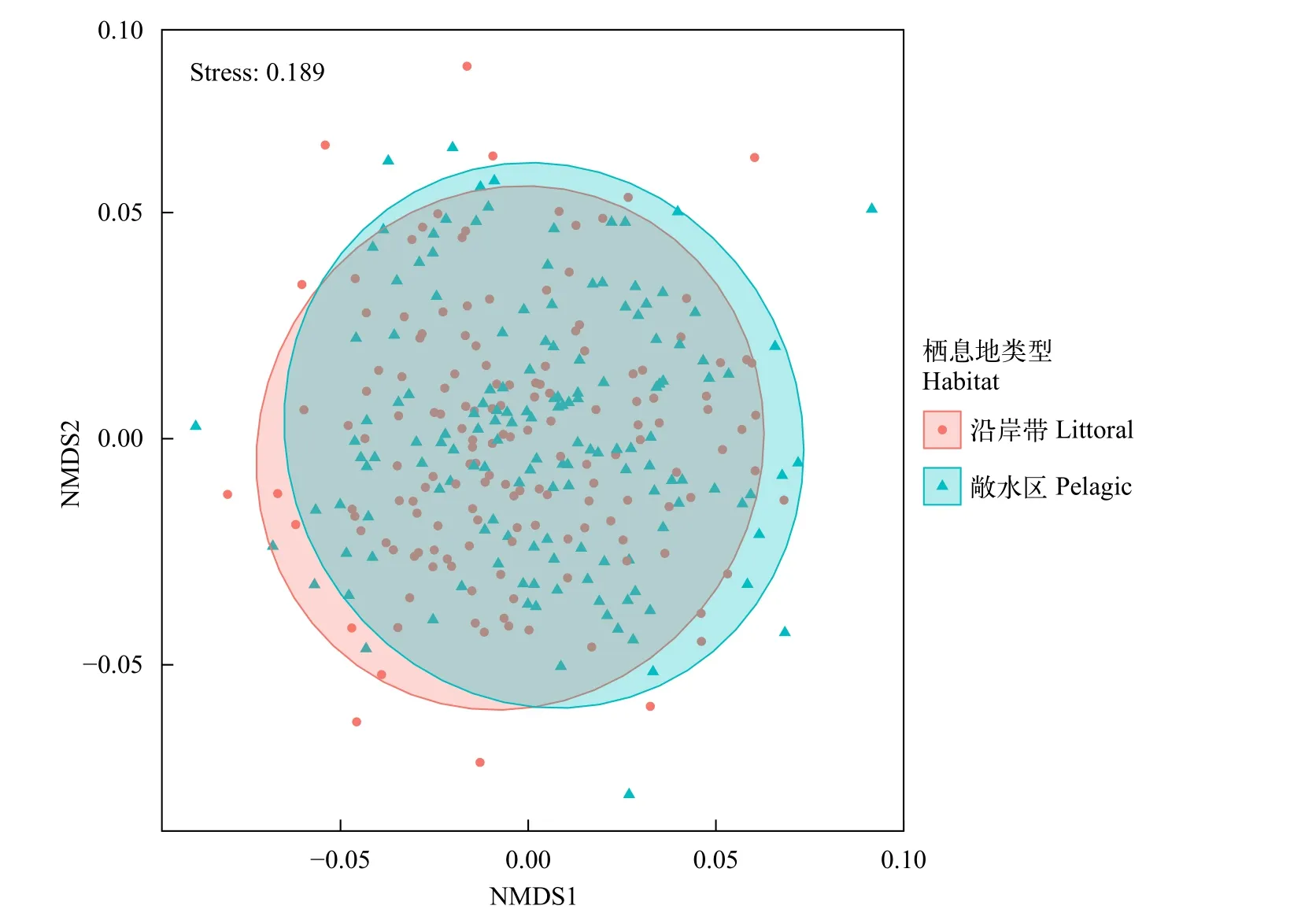
图3 两种栖息地鲌类食物来源NMDS图Fig. 3 Food sources NMDS plot of culters in different habitats
3 讨论
在湖泊中, 鱼类水体空间类型包括敞水区与沿岸带等, 鱼类往往占据沿岸带或者敞水区生境二者之一, 并能形成适应于栖息生境条件下的独特形态。然而, 鱼类由于具有高度的流动特性, 并且由于捕食行为在各种生境间来回游动, 因此往往被认为是水体小生境之间物质与能量的“连接者”与“整合者”[31,32]。敞水区栖息地为湖泊开阔水域, 食物链模式以往主要被认为是浮游植物→浮游动物→浮游食性鱼类→肉食性鱼类[33], 但由于鱼类的运动能力强且具有灵活的捕食策略, 将湖泊不同栖息地紧密联系起来, 其食物组成还应包括沿岸带、底栖等生境中生物与碎屑组分[34]。
某些鱼类群落依靠沿岸带与敞水区两种生境的能量途径, 且两种能量途径对于鱼类群落来说同等重要[35]。保安湖沿岸带与敞水区两种生境中的高营养级捕食者的不同食物来源比重基本相似, 表明保安湖鲌类在两种栖息地中均有摄食行为; 且两种生境中鲌类的食物来源较为广泛, 基本涵盖了沿岸带与敞水区的所有饵料生物, 表明其能够将来源于两种生境中的能量整合到一起。
以往的研究资料表明, 具备灵活运动能力的鱼类或顶级捕食者是水域生态系统中不同生境的“连接者”[36], 能够维持生态系统中食物网的稳定性[11,37]。在以往的研究资料中, 捕食者(指鱼类)对于不同生境所起到的耦合作用的大小通常基于食物组成进行分析, 高营养级捕食者能够从水体各个生境中均衡地获取食物被认为耦合能力更强[38—40]。本研究运用稳定同位素质量平衡混合模型与非度量多维尺度分析, 对保安湖高营养级捕食者营养来源的分析也支持以上观点。保安湖高营养级捕食者—达氏鲌与红鳍原鲌在运动与捕食过程中, 将来自沿岸带与敞水区两种生境中的食物链的营养与能量整合在一起, 实现了不同湖泊生境间的耦合, 对于维持保安湖生态系统的营养循环、食物网功能的完整性与稳定性发挥了重要作用[34]。
