岩溶石漠化不同修复林分的生物量和生产力及其动态变化*
2022-04-12周晓果孙冬婧温远光明安刚贾宏炎朱宏光赵岩岩黄勇杰
周晓果,孙冬婧,温远光,**,王 磊,明安刚,贾宏炎,朱宏光,赵岩岩,黄勇杰,梁 佳
(1.广西科学院生态环境研究所,广西南宁 530007;2.广西大学林学院,广西森林生态与保育重点实验室,广西南宁 530004;3.中国林业科学研究院热带林业实验中心,广西凭祥 532600)
生物量和生产力是森林生态系统结构和功能中最基本的数量特征,既是人类衣、食、住、行、药等资源的物质基础,也是地球生物圈生命支持系统的核心[1,2]。人类不合理的利用以及全球气候变化,使地球上大面积的森林植被遭到破坏,导致陆地植物生物量和地球总生物量减半[3],危及人类自身的生存和发展[2,3]。如何防止地球植被的持续退化、恢复和重建已退化的森林植被、优化人类赖以生存的植被资源,成为人类必须解决的重大科学难题和紧迫任务[1]。
有研究表明,生物量和生产力是判定和评价生态系统修复、稳定性与可持续性的基础[4,5],林分生物量和生产力的提高有助于提高土壤有机质、促进养分循环、增加生物多样性、维持生态系统结构与功能稳定性、加快退化生态系统修复[6-8],成为当今全球森林生态系统和气候变化研究的核心内容之一[9,10]。当前有关森林生物量和生产力的研究论文卷帙浩繁,由于岩溶(喀斯特)森林生物量的取样十分困难,人们对岩溶森林生物量的研究较少。目前仅有的少数研究是利用收获法和标准木法对贵州茂兰喀斯特森林群落地上生物量[11,12],桂西喀斯特地区黄荆、红背山麻杆、灰毛浆果楝灌丛生物量及生产力[13]和广西石山林(树种组)生物量[14]进行测定,利用前人建立的生物量方程对喀斯特植被不同演替阶段的生物量和净初级生产力进行估算[15-17];最近有学者对贵州喀斯特地区马尾松天然林、马尾松人工林和湿地松人工林生态系统生物量进行研究[18]。然而,以往关于岩溶地区森林生物量的研究更多的是天然次生林、灌丛植被以及人工纯林,对于纯林和混交林的比较,特别是长期固定样地的生物量研究极少,对岩溶森林生产力的研究更少。由于缺乏对人工修复林分的长期监测,无法对修复林分的生长和适应性做出科学评价,导致盲目推广应用,造成重大损失[19]。
本研究基于2011年建立的顶果木Acrocarpusfraxinifolius纯林(PAFP)、降香黄檀Dalbergiaodorifera纯林(PDOP)和顶果木×降香黄檀混交林(MADP)长期固定监测样地,对2013年、2015年、2019年和2021年的4次林分生物量进行调查,探讨岩溶石漠化不同修复林分生物量和生产力随时间的动态变化特征,为进一步探索3种人工修复林分生物量和生产力长期动态变化规律提供基础数据和资料,为石漠化综合治理及林分质量精准提升提供科学依据。
1 材料与方法
1.1 研究区域的自然环境概况
研究区域位于广西南宁市马山县(23°24′-24°2′N,107°41′-108°29′E),属南亚热带季风气候区,气候温暖湿润,雨热同季,干湿季分明。年平均气温21.3℃,年平均降水量1 667.1 mm。境内岩溶广布,土壤以岩溶石灰土为主,土层浅薄,岩基裸露度大。原生性的地带性森林植被荡然无存,生态环境脆弱。
1.2 样方设置与生物量调查
基于2011年在广西南宁市马山县民族村,按近自然经营建立顶果木纯林(PAFP)、降香黄檀纯林(PDOP)和顶果木×降香黄檀混交林(MADP)试验林分,2013年建立长期固定监测样地,每种林分样地面积为20 m×20 m,设3个重复,林分环境概况见表1。分别于2013年、2015年、2019年和2021年进行样地群落生物量调查,并分别于2015年、2019年和2021年进行标准木生物量测定。累计采伐顶果木、降香黄檀标准木各9株。采用收获法测定标准木的器官生物量,将器官分为干、枝、叶、根4个组分,枝和根进一步细分为一级枝、二级枝、三级枝和细根(<0.5 cm)、中根(0.5-1.0 cm)、粗根(>1.0 cm)、根蔸,分别称取各器官生物量鲜重,同时分别采集各器官样品约500 g。将所有样品带回实验室,在85℃恒温下烘干至恒重,进而计算各样品的含水率和干重。
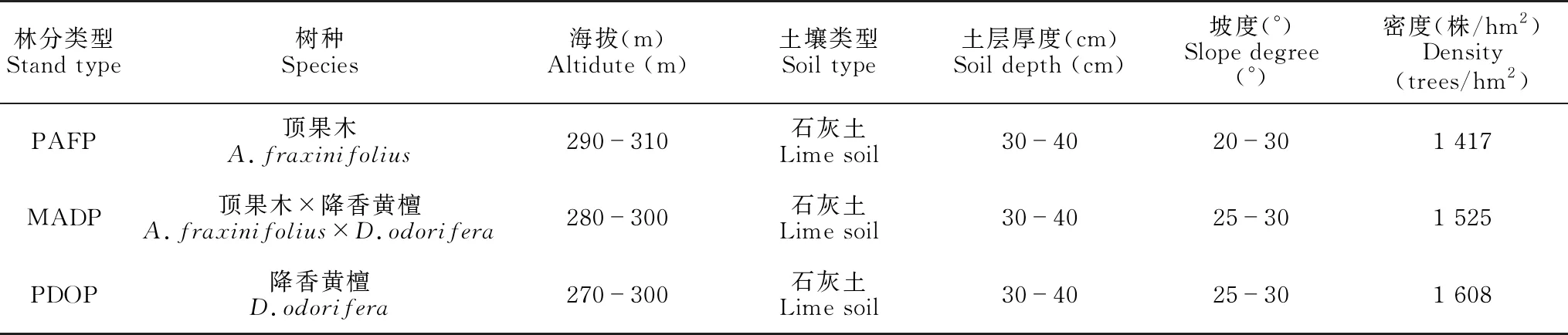
表1 林分环境概况
1.3 生物量和生产力的计算
根据标准木胸径和树高以及器官生物量之间的相对生长关系,建立不同树种、不同器官的生物量估测回归方程(表2),各回归方程的相关性均达极显著相关(P<0.001)。根据表2中的生物量方程结合不同年份各调查样地林木胸径和树高计算林木各器官的生物量,将样地林木各器官生物累加得到林分生物量。关于森林第一性净生产力指标有平均净生产力(总平均或定期平均)和年间净生产力(连年生长量)两种,更多的是采用生物量除以年龄,即总平均生长量作为生产力指标[5]。本文采用定期平均净生产力作为生产力指标。
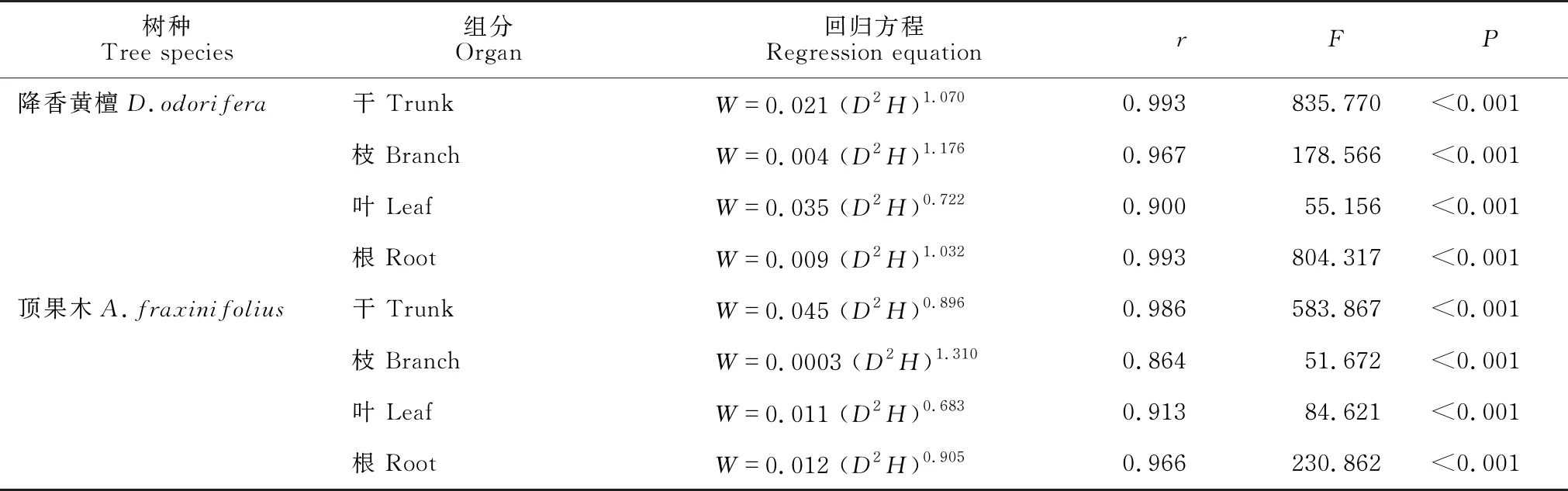
表2 不同树种不同器官生物量估算回归模型
1.4 数据统计分析
采用单因素方差分析(One-way ANOVA)检验不同林分、不同树种、不同器官生物量和生产力的差异显著性,显著性水平设置为P<0.05,采用最小显著性差异法(Least Significant Difference,LSD)进行多重比较,以上分析在SPSS 24.0软件上运行。
2 结果与分析
2.1 不同林分不同树种单株生物量的动态变化
由图1可知,3种林分两树种单株和各器官生物量均随着林分年龄的增加而显著增加。PAFP林分在2013年、2015年、2019年和2021年单株生物量分别为10.41 kg、14.45 kg、20.24 kg、25.83 kg;MADP林分中,顶果木单株生物量分别为8.92 kg、16.03 kg、21.92 kg、27.87 kg,降香黄檀单株生物量分别为3.02 kg、8.13 kg、26.47 kg、51.49 kg;PDOP林分单株生物量分别为2.94 kg、7.64 kg、36.07 kg、56.39 kg。方差分析表明,PAFP林分单株和各器官生物量各年份间存在显著差异(P<0.05);PDOP林分除树干、树枝和树根在2013年和2015年间差异不显著(P>0.05)外,其余各年份间差异显著(P<0.05);MADP林分中顶果木平均单株和器官生物量变化与PAFP林分相同,而降香黄檀则与PDOP相同;混交林中顶果木和降香黄檀的单株生物量和树叶、树根生物量存在显著差异(P<0.05),其中2013年和2015年为顶果木极显著高于降香黄檀(P<0.001),而2019年和2021年为降香黄檀极显著高于顶果木(P<0.001);除2019年的树干和2013年的树枝外,其余各年份间均存在极显著差异(P<0.001)。
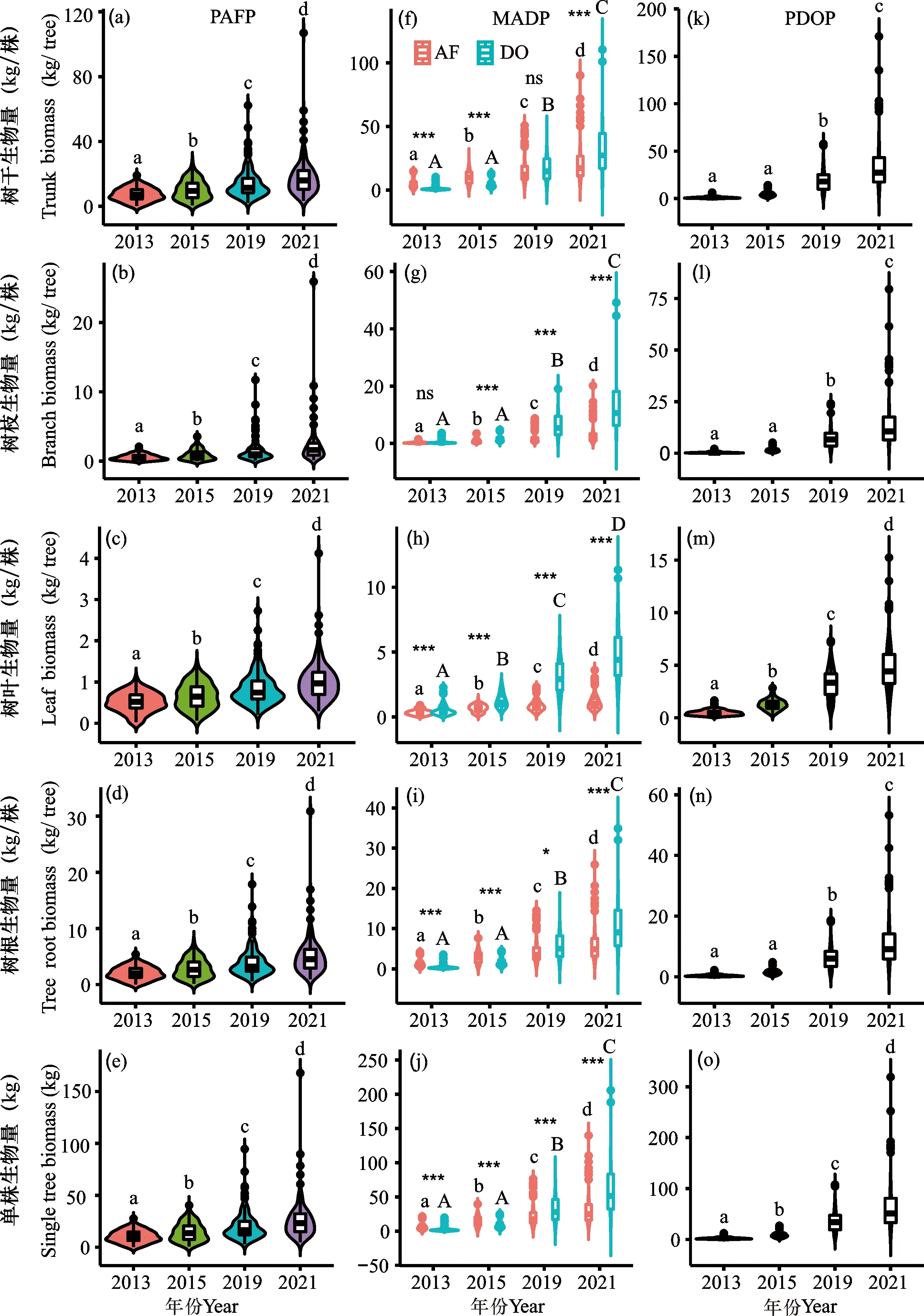
AF:顶果木;DO:降香黄檀;(a)-(e)、(k)-(o)不同小写字母表示同一林分不同年份间差异显著(P<0.05);(f)-(j)不同小写字母表示混交林中不同年份间顶果木胸径差异显著(P<0.05);不同大写字母表示混交林中不同年份间降香黄檀胸径差异显著(P<0.05);***表示同一年份顶果木与降香黄檀胸径差异极显著(P<0.001);*表示同一年份顶果木与降香黄檀胸径差异显著(P<0.05);ns表示同一年份顶果木与降香黄檀胸径差异不显著(P>0.05)
2.2 不同林分生物量的动态变化
由表3可知,随着林分年龄的增长,除2013年和2015年树枝生物量差异不显著外,3种林分各器官生物量及总生物量均显著增加(P<0.05)。方差分析表明,3种林分的总生物量各年份间均存在显著差异(P<0.05)。在林分生长初期(2013年),PAFP林分总生物量显著高于MADP和PDOP(P<0.05);2015年为PAFP和MADP显著高于PDOP (P<0.05);而后期阶段(2019年和2021年)却为PDOP显著高于MADP和PAFP (P<0.05),MADP显著高于PAFP (P<0.05)。
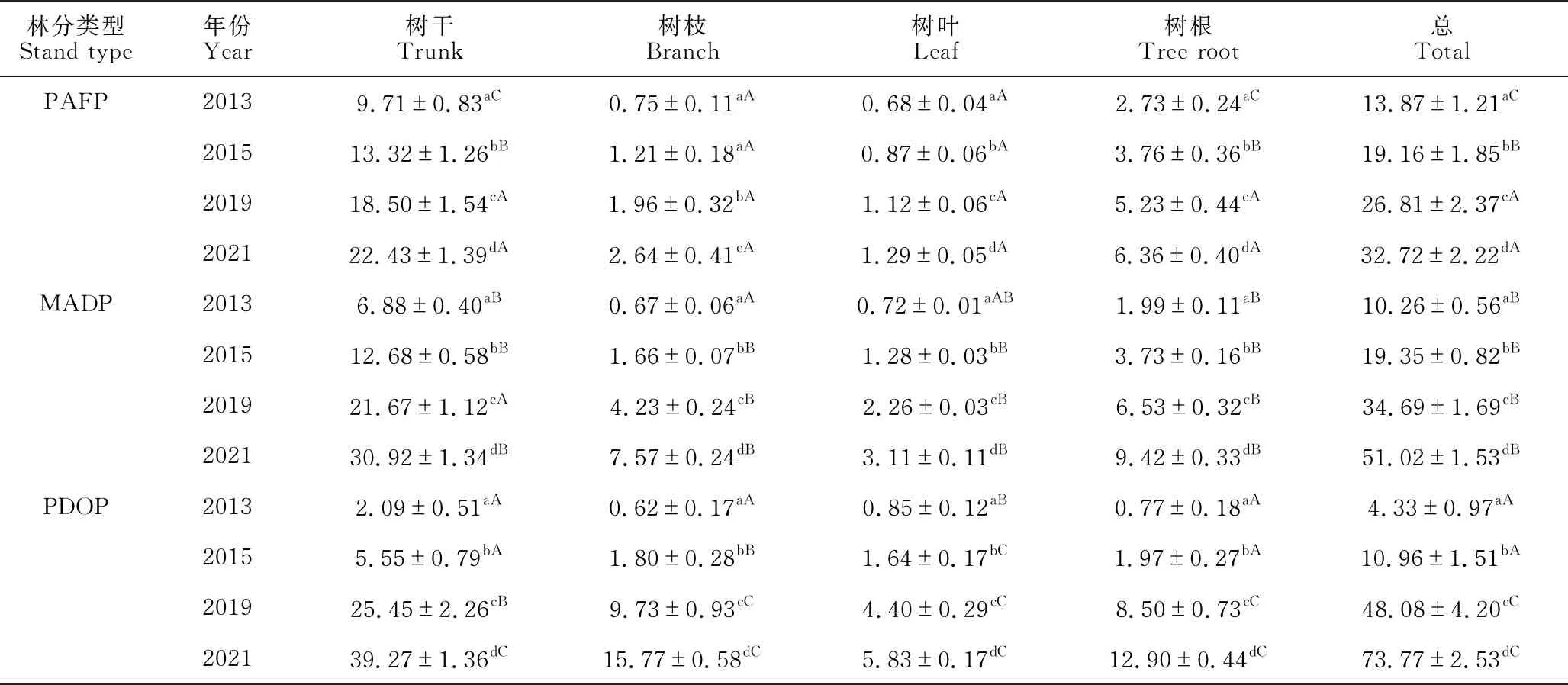
表3 不同林分中不同器官生物量的动态变化(t/hm2)
2.3 不同林分不同树种各器官生物量分配比例的动态变化
由表4可知,随着林分年龄的增长,3种林分两树种各器官生物量分配比例存在明显的动态变化。无论是纯林还是混交林,顶果木树干生物量分配比例随年龄的增加呈递减的趋势,而降香黄檀则表现为递增的趋势。两树种树枝生物量分配比例随年龄的增加呈递增的趋势,而两树种树叶生物量分配比例的变化规律相似,均为随着年龄的增加而显著降低(P<0.05)。不同年份两树种根系生物量分配比例随时间的推移变化都很小,变化在1%以内,但变化较复杂。顶果木纯林2013年、2015年和2019年根系生物量分配比例显著高于2021年(P<0.05),而2013年、2015年和2019年间差异不显著(P>0.05)。而混交林中,顶果木根系生物量2013年和2015年显著高于2019年(P<0.05);降香黄檀为2013年显著低于2015年、2019年和2021年(P<0.05),而2015年和2019年显著高于2021年(P<0.05)。
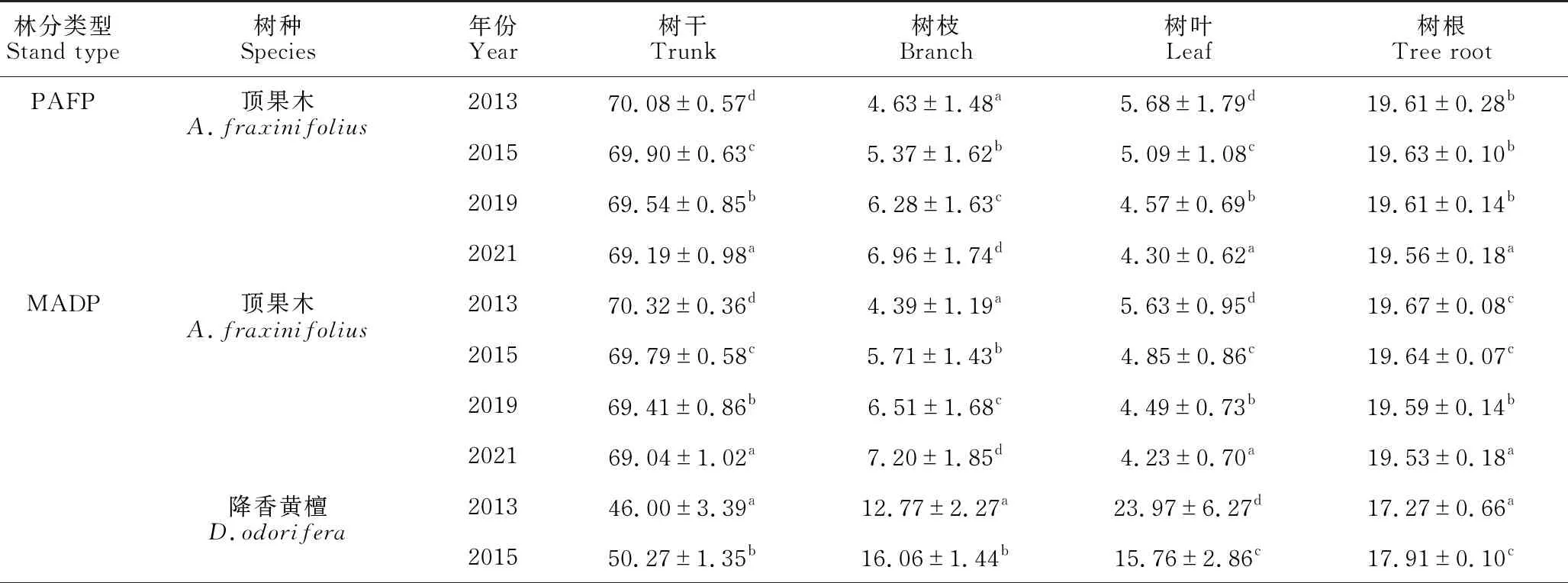
表4 不同林分不同树种各器官生物量分配比例的动态变化(%)
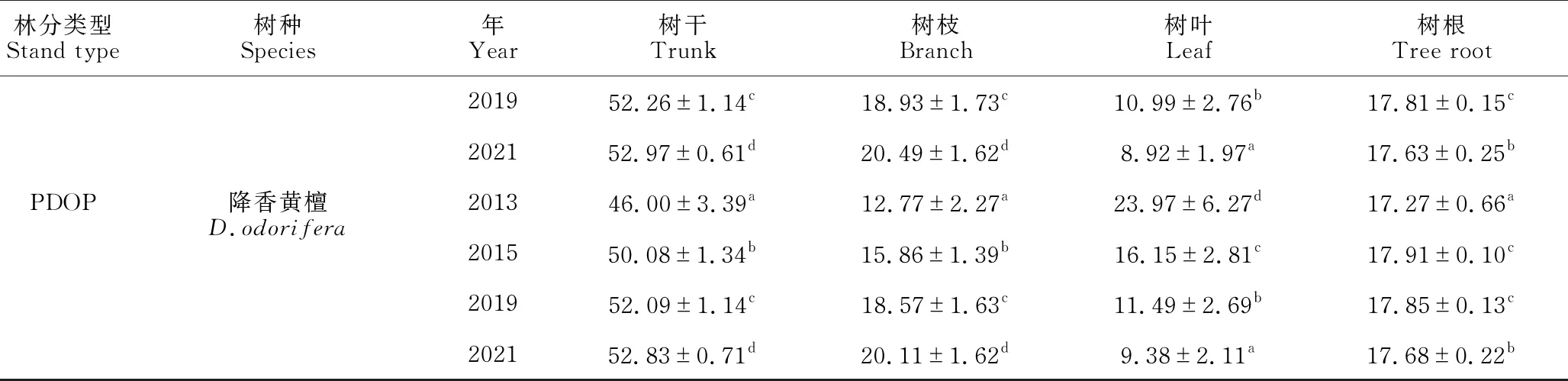
续表
2.4 不同林分各器官生物量分配比例的动态变化
由表5可知,PAFP和MADP林分树干生物量分配比例随年龄的增加呈显著递减的趋势(P<0.05),而PDOP却表现为显著递增的趋势(P<0.05)。3种林分树枝生物量比例均随着林分年龄的增长而显著增加(P<0.05),但不同林分各年份间差异不同,PAFP为2019年和2021年显著高于2013年和2015年(P<0.05),MADP和PDOP林分树枝生物量为各年份间差异显著(P<0.05)。3种林分树叶生物量比例表现为显著递减趋势(P<0.05),PAFP和PDOP为2013年和2015年显著高于2019年和2021年(P<0.05),2013年显著高于2015年(P<0.05);而MADP则表现为2013年显著高于2021年(P<0.05),其余各年份间均差异不显著(P>0.05)。3种林分树根生物量分配比例总体上随着林分年龄的增加而递减,PAFP为2013年和2015年显著高于2021年(P<0.05),MADP为各年份间均差异显著(P<0.05),PDOP为2015年显著高于2013年、2019年和2021年(P<0.05),2013年和2019年显著高于2021年(P<0.05),但2013年与2019年间差异不显著(P>0.05)。
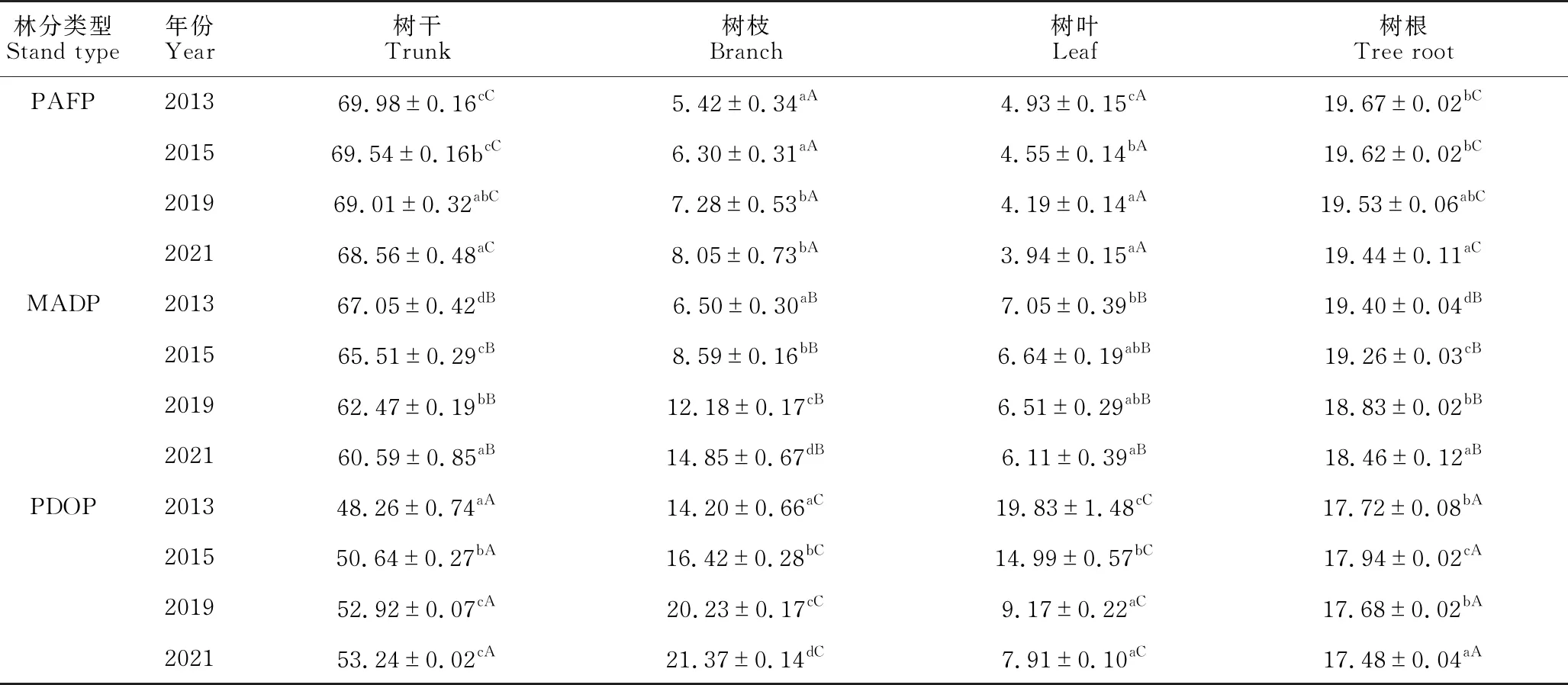
表5 不同林分不同器官生物量分配比例(%)
2.5 不同林分平均净生产力的动态变化
由图2可知,在2013-2015年、2015-2019年和2019-2021年,PAFP定期年平均净生产力分别为2.64 t/hm2、1.92 t/hm2、2.95 t/hm2;MADP分别为4.55 t/hm2、3.83 t/hm2、8.17 t/hm2;PDOP相应分别为3.32 t/hm2、9.28 t/hm2、12.84 t/hm2。PAFP、MADP和PDOP 3种林分在2013-2021年的定期年平均净生产力分别为2.36 t/hm2、5.09 t/hm2、8.68 t/hm2。方差分析表明,除2013-2015年3种林分的定期年平均净生产力无显著差异外(P>0.05),2015-2019年、2019-2021年和2013-2021年均表现为PDOP显著高于MADP和PAFP,MADP显著高于PAFP (P<0.05)。3种林分树干和树根定期年平均净生产力的差异性变化规律与林分定期年平均净生产力相同;树枝2013-2015年表现为MADP和PDOP显著高于PAFP (P<0.05),前两者无显著差异(P>0.05);2015-2019年、2019-2021年和2013-2021年均表现为PDOP显著高于MADP和PAFP(P<0.05),MADP显著高于PAFP (P<0.05);而树叶4个定期年平均净生产力指标均表现为PDOP显著高于MADP和PAFP (P<0.05),MADP显著高于PAFP (P<0.05)。
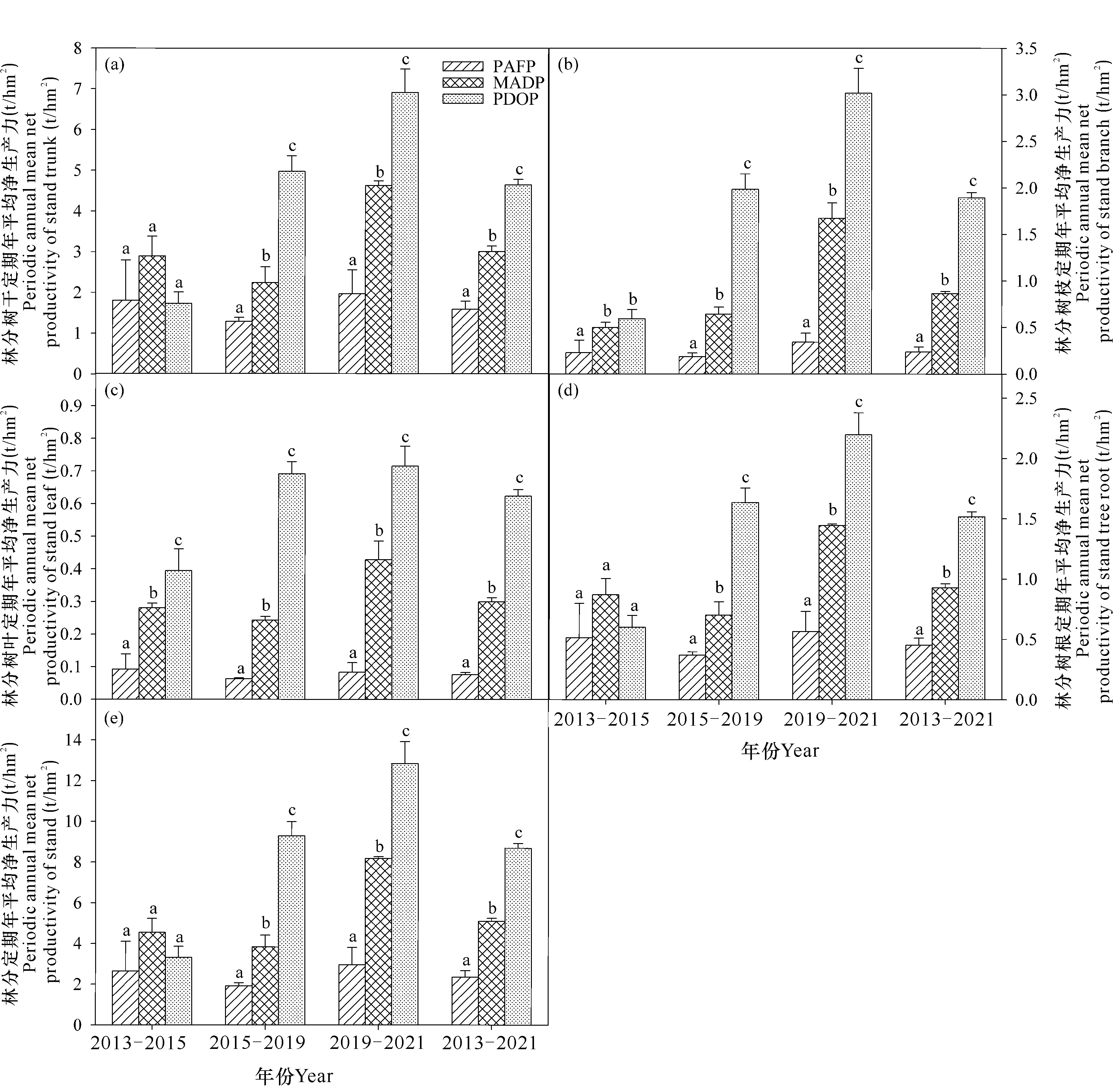
不同小写字母表示同一时期不同林分间差异显著(P<0.05)
3 讨论
3.1 林分类型对岩溶森林生物量的影响
林分生物量的形成和积累与生物气候条件、立地生境、林分状况、树种性质、林分年龄以及人为干扰等因素有关[12]。岩溶地区生态环境恶劣,森林生物量明显低于同区域的非岩溶森林[20],属于低生物量的森林生态系统[11,12,13-18]。有研究表明,喀斯特天然林的生物量是随着林分年龄的增加而增加,从幼龄林、中龄林、近熟林、成熟林到过熟林,林分生物量分别为50-60 t/hm2、90-110 t/hm2、120-140 t/hm2、160-240 t/hm2和331.46 t/hm2[11-16];而喀斯特地区18 a生马尾松天然林和19 a生马尾松人工林乔木生物量分别为103.46 t/hm2、140.55 t/hm2[18]。本研究中,3种修复林分的生物量均随着林分年龄的增长而显著增加,以PAFP的生物量最低,其值为(32.72±2.22) t/hm2,其次是MADP,其值为(51.02±1.53) t/hm2,PDOP最高,其值为(73.77±2.53) t/hm2。本研究表明,修复10 a的林分生物量明显低于保存较好的原生性森林[11,12],也明显低于18-19 a生松林[18],PAFP的生物量明显低于同区域的次生幼林,MADP略高于同区域的次生幼林,而PDOP却显著高于同区域的次生幼林[17]。3种修复林分的生物量是当地黄荆灌丛的2-4.6倍[13],说明采用豆科固氮树种修复石漠化可以显著提高林分生物量,且PDOP的生物量持续修复作用明显优于PAFP。本研究结果为石漠化植被修复中的树种和林分选择提供了科学依据。
3.2 树种和林分类型对岩溶森林生物量分配的影响
由于林分所处的环境,以及林分树种组成、年龄、密度、林木生长级别不同,生物量的分配也不同[4]。岩溶地区与非岩溶地区、针叶树种与阔叶树种、常绿树种与落叶树种、优势木与劣势木、人工林与天然林以及不同年龄阶段林分各器官的生物量分配明显有区别[14,20,21]。由于岩溶森林生态系统生境异质性强、树种组成复杂多样、同一树种个体数少、个体分化明显、树枝和树叶生物量估测方程精度低等[12,15],关于岩溶森林生物量分配的研究成果较少。刘之洲等[18]对岩溶地区18-19 a生针叶林的生物量分配研究发现,马尾松天然林的树干(64.15%)和树根(11.56%)生物量分配比例高于马尾松人工林树干(57.54%)和树根(9.41%),而马尾松人工林的树枝(25.83%)和树叶(7.22%)生物量分配比例却高于马尾松天然林树枝(17.47%)和树叶(6.82%)。朱守谦等[12]的研究也发现,岩溶广东松林地上生物量分配比例以树枝最高,为51.05%,其次才是树干,为41.43%,树叶的生物量分配比例仅为7.52%。
本研究表明,不同树种各器官生物量的分配比例存在显著差异,而且随着林分年龄的增加,各器官生物量分配比例也在发生明显的变化。本研究中,10a生林分,PAFP各器官生物量分配比例为树干(68.56%)>树根(19.44%)>树枝(8.05%)>树叶(3.94%),而PDOP却表现为树干(53.24%)>树枝(21.37%)>树根(17.48%)>树叶(7.91%)。本研究还发现,随着林分年龄的增长,不同树种不同器官生物量分配比例的变化规律并不一致,PAFP树干、树叶和根系生物量分配比例随着年龄的增加而显著下降,而树枝生物量分配比例却显著增加;PDOP树干和树枝生物量分配比例随着年龄的增长而显著增加,而树叶和树根却显著下降。同时发现,混交林分不同器官生物量的分配比例处于两种纯林(PAFP和PDOP)之间。上述结果表明不同林分不同树种各器官生物量的分配规律会随着林分年龄的增加而发生改变,揭示了岩溶地区PDOP和PAFP器官生物量随年龄变化的分配规律。因此,在未来森林生物量和碳储量研究中应充分考虑这种变化,以提高估测精度。
3.3 树种和林分类型对岩溶森林生产力的影响
关于岩溶森林平均净生产力的研究较少,有研究表明茂兰喀斯特顶极森林和次顶极森林乔木层的年度净初级生产力分别为7.06 t/hm2和5.09 t/hm2[15],而贵州喀斯特地区马尾松天然林、马尾松人工林乔木层年平均净生产力分别为5.73 t/hm2和7.39 t/hm2[18](作者根据其林分生物量和年龄修订)。本研究中,2013-2021年,林分定期年平均净生产力以PDOP (8.68 t/hm2)最高,PAFP (2.36 t/hm2)最低,MADP (5.09 t/hm2)居二者之间。可见,除PAFP外,PDOP的净初级生产力明显高于岩溶顶极森林和马尾松人工林,MADP也与岩溶次顶极森林持平。说明采用豆科固氮树种修复10 a的林分净生产力达到甚至超过了岩溶顶极和次顶极森林乔木层的净生产力,PAFP、MADP、PDOP在2019-2021年的平均净生产力比2015-2019年分别提高53.65%、113.32%、38.36%,实现了岩溶石漠化生态系统生产力的快速修复和提升。
岩溶顶极森林地上部分净初级生产力以树叶的占比最高(39.75%),其次是树干(34.68%),树枝仅占6.91%;而次顶极森林中树叶净初级生产力的分配比例更高,达68.57%,树干的比例下降为22.59%,树枝为8.84%[15]。这与天然森林树木的生产结构较人工林变化大,岩溶森林小生境的特殊性、多样性以及组成树种的复杂性有关,从而造成岩溶天然森林树枝、树叶生物量及其垂直分布的多变和异常[12]。岩溶地区马尾松天然林树干、树枝、树叶平均净生产力分别占地上部分生产力的72.58%、19.72%、7.69%,马尾松人工林树干、树枝、树叶平均净生产力分别占地上部分生产力的63.53%、26.40%、7.92%[18]。本研究中3种修复林分地上部分净初级生产力分配格局与岩溶地区天然顶极、次顶极森林不同[15],与马尾松天然林、人工林相似[18],这是由树种特性和林分结构不同所致。
本研究还发现,不同林分、不同树种和不同器官的定期平均净生产力随着林分年龄的增加存在明显的变化,PAFP树叶的年平均净生产力非常低,仅为0.08 t/hm2,只占PDOP (0.62 t/hm2)的12.90%和MADP (0.30 t/hm2)的26.67%。PAFP树叶的生产能力显著下降,光合面积持续减少,这可能是随着时间推移PAFP逐渐衰退的重要原因。同时也表明顶果木是一个喜肥的树种,在早期施肥的情况下,生长表现非常好,但后期停止施肥后,生长缓慢。因此,造林应选择在土壤较肥沃的地方。
4 结论
研究表明,3种修复林分生物量均随时间的推移而显著增加。随着林分年龄的增长,不同林分、不同树种各器官生物量均显著增加,但变化规律有所不同。PAFP树干、树叶和树根生物量分配比例显著下降,树枝生物量比例显著增加;PADP却表现为树干、树枝生物量分配比例显著增加,树叶和树根生物量比例显著下降;MADP各器官生物量分配比例居两者之间。3种修复林分在2015-2019年、2019-2021年和2013-2021年的林分定期平均净生产力均存在显著差异,表现为PDOP>MADP>PAFP,3种修复林分间差异显著;经过10 a的修复,豆科固氮树种人工林的林分净生产力达到甚至超过了岩溶顶极、次顶极森林乔木层的净生产力,实现了岩溶石漠化生态系统生产力的快速修复和提升。
