西南岩溶地区天然次生林群落不同层次的构建机制*
2022-04-12周晓果孙冬婧温远光陆志成朱宏光
周晓果,孙冬婧,温远光,**,王 磊,陆志成,朱宏光
(1.广西科学院生态环境研究所,广西南宁 530007;2.广西大学林学院,广西森林生态与保育重点实验室,广西南宁 530004;3.北京林业大学生态与自然保护学院,北京 100083)
受人类高强度活动和不合理开发利用的影响,我国西南岩溶地区的森林资源锐减,生态功能退化,绝大部分地区沦为次生性的灌木林和灌草丛[1],部分地区石漠化[2],严重影响西南岩溶地区的生态安全和经济社会的可持续发展[2,3]。进入21世纪,国家明确将西南岩溶地区石漠化综合治理作为重大生态工程实施并上升为国家战略[4],极大地促进了天然次生林的形成和恢复。天然次生林是原始林经过采伐或多次破坏后自然恢复起来的森林[5]。我国天然次生林约占全国森林总面积的46.2%,约占全国森林总蓄积的23.3%,在水源涵养、水土保持、珍贵木材资源储备以及生物多样性保护等方面具有重要的作用[6-8]。然而,由于天然次生林多为中、幼龄林,林分结构不稳定,大部分天然次生林由于遭受多次破坏沦为低质低效林[6,7]。森林群落不同层次的物种组成决定着林分的结构、质量和演替方向[9]。因此,深入开展岩溶地区天然次生林群落不同层次的物种组成及构建机制研究,对加快天然次生林培育、持续提升林分质量具有重要意义。
干扰后森林结构的进展演替一般从密度、胸高断面积、冠层高度和冠层群落多样性等方面进行评估[1]。早期的研究表明,天然次生林恢复过程中的群落组成受到多种因素的影响。例如,植物繁殖体的迁移和扩散影响种子的到达和定居[10],植物物种的生态特性影响植物对外界干扰和环境变化的响应和适应[1],而种间和种内关系则影响植物群落的生长、发育和组成[11]。此外,群落环境的变化影响资源的分布和有效性,导致土壤N、P的限制和pH的变化[12]。最重要的是,扰动历史和土地利用方式强烈地影响土壤性质[12,13]、残余繁殖体(如种子库、芽和幼树)的存在以及当前植物群落的物种组成[14]。这导致了天然次生林演替过程中微生境和小气候的复杂性。与此同时,冠层中的大型木本植物通常具有气候主导的生态位,林下植物具有土壤和微生境主导的生态位[15],从而导致森林群落不同层次植物组成的差异和林分质量的不同。最近的研究表明,森林群落不同层次的植物组成受不同构建机制的影响[16]。然而,目前对岩溶地区天然次生林群落不同层次的构建机制尚不清楚,量化不同群落层次结构和生态环境因子对森林群落不同层次构建作用的影响意义重大。
本研究以西南岩溶地区天然次生林为研究对象,根据《中华人民共和国林业行业标准 主要树种龄级与龄组划分》(LY/T 2908—2017)[17],研究对象分属青冈栎幼龄林(恢复期为20-30 a)、中龄林(恢复期为40-50 a)和近熟林(恢复期为60-70 a),探究岩溶地区天然次生林群落不同林层的构建机制及其主控因子,为岩溶地区森林生态保护、修复和管理提供科学依据。
1 材料与方法
1.1 研究区域和样地概况
研究区域位于广西南宁市马山县(23°24′-24°02′ N,107°41′-108°29′ E),属南亚热带季风气候区。年平均气温21.3℃,1月平均气温12.1 ℃,7月平均气温28.2 ℃。年平均降水量1 667.1 mm,4-9月为雨季,10月至翌年3月为旱季。该地区岩溶地貌广布,属于滇黔桂石漠化片区。土壤主要是石灰岩土,土层浅薄,一般在40 cm以内,分布不均,岩石裸露率为20%-50%[18]。
2019年5月,在查阅相关资料和询问当地群众明确研究区域土地利用历史和林分年龄的基础上,选择代表性的幼龄林、中龄林和近熟林为研究对象。幼龄林是指森林中的林木遭到不同程度的砍伐破坏后,由原来的林木伐根萌蘖迅速生长、层次结构尚无明显分化,林分年龄在40 a以下的林分。中龄林主要是指层次结构明显分化,林分年龄为40-60 a的林分。近熟林是指群落的组成和结构接近当地顶极群落,年龄在60-80 a的林分。幼龄林乔木层优势种为青冈栎(Cyclobalanopsisglauca,重要值128.5)、光叶海桐(Pittosporumglabratum,重要值42.8)、小果化香(Platycaryaglandulosa,重要值31.1),灌木层优势种为矮棕竹(Rhapishumilis,重要值144.0)、龙须藤(Bauhiniachampionii,重要值35.5)、美丽胡枝子(Lespedezaformosa,重要值20.8),草本层优势种为粽叶芦(Thysanolaenalatifolia,重要值172.5)、薄叶卷柏(Selaginelladelicatula,重要值66.1)、白茅(Imperatacylindrica,重要值32.1)。中龄林乔木层优势种为青冈栎(重要值92.4)、苦木(Picrasmaquassioides,重要值32.7)、海红豆(Adenantheramicrosperma,重要值21.2),灌木层优势种为青冈栎(重要值29.7)、龙须藤(重要值22.4)、网脉酸藤子(Embeliarudis,重要值18.8),草本层优势种为井栏边草(Pterismultifida,重要值92.0)、薄叶卷柏(重要值86.3)、灰绿耳蕨(Polystichumanomalum,重要值33.0)。近熟林乔木层优势种为青冈栎(重要值88.4)、海红豆(重要值55.4)、南酸枣(Choerospondiasaxillaris,重要值47.8),灌木层优势种为西南红山茶(Camelliapitardii,重要值76.1)、青冈栎(重要值51.3)、海红豆(重要值37.7),草本层优势种为团叶鳞始蕨(Lindsaeaorbiculatavar.orbiculata,重要值90.9)、斜羽凤尾蕨(Pterisoshimensis,重要值87.6)、灰绿耳蕨(重要值64.8)。
在上述3种林分的典型地段,分别设置9个20 m×20 m的样方,将每个样方细分成4个10 m×10 m的小样方,样方间相隔300 m。不同林分样地的地形和0-20 cm土层土壤特征见表1。
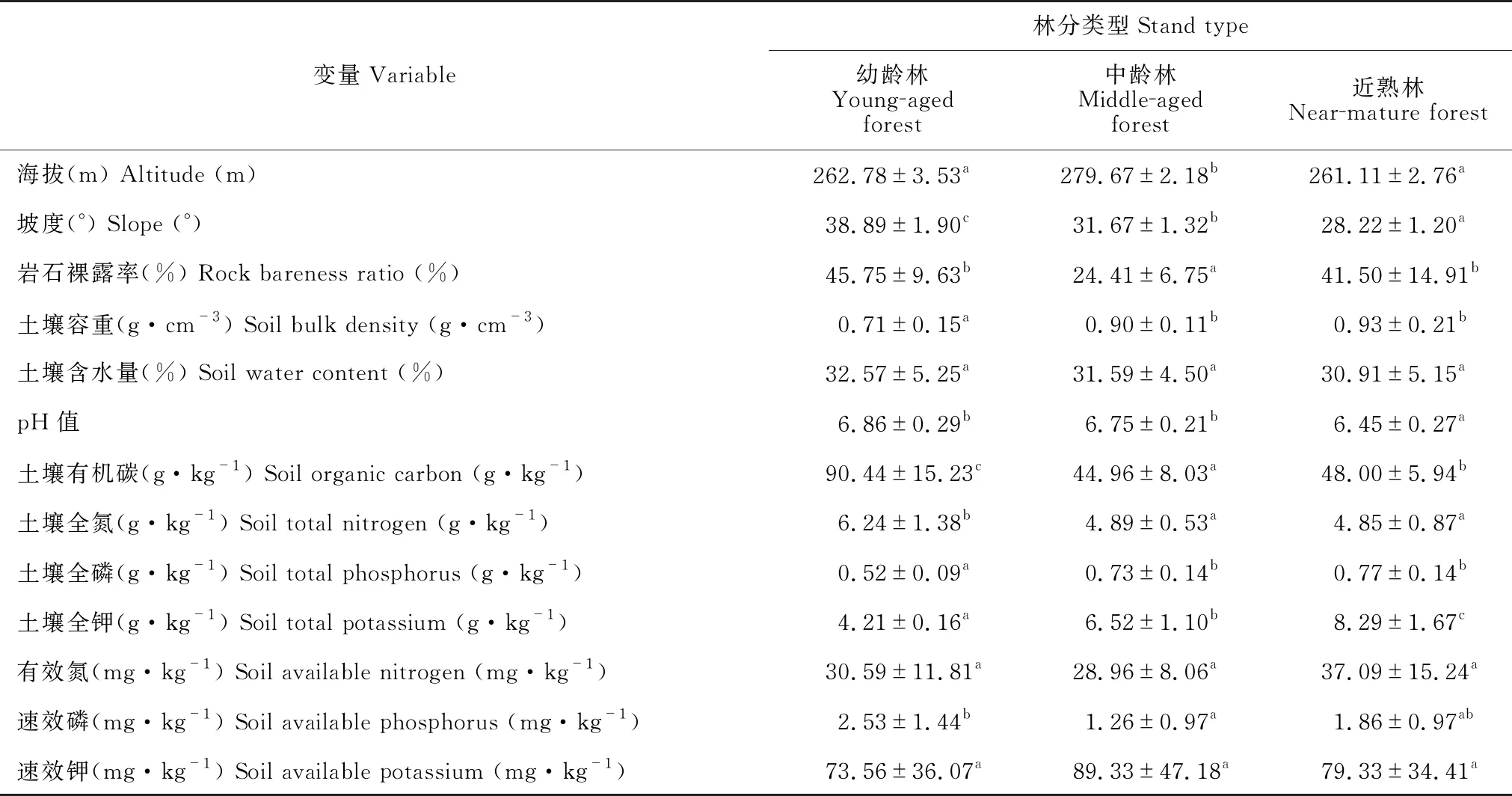
表1 不同林分样地的地形和土壤(土层0-20 cm)特征
1.2 群落调查和生物量估算
在每个样方,对胸径(DBH)≥2 cm的林木进行全面调查,记录物种名、胸径、树高(H)和冠幅。将所有乔木分为林冠层(DBH≥5 cm,H≥5 m)和林下层(DBH<5 cm,H<5 m)。在调查过程中,分别记录个体数、萌芽个体数,以每棵树的萌芽个体数来表征演替过程中的再生能力。将各层的总个体数和萌芽个体数相加作为茎干数,计算茎干密度和平均单株茎干数。
根据样方调查数据,采用相对生长法计算树木地上生物量(TAB)[19]:
TAB=0.0755×(DBH2×H)0.8941,
式中:DBH为胸径(cm),H为树高(m)。
1.3 群落多样性、重要值和谱系多样性计算
(1)基于调查数据,统计群落调查样方内的物种组成,计算物种丰富度指数(R)和Shannon-Wiener指数(H′)[20]:
R=S,
式中,S是样方中出现的物种数;Pi=ni/N,其中ni为样地内某种的个体数,N为样地内总个体数。
(2)计算每个林冠层和林下层各物种的重要值(IV)[21]:
IV=100×(Ra+Rf+Rc)/3,
式中,Ra为相对丰度,是每个样方林冠层(或林下层)某一物种的个体数与样方中所有物种个体数的比值;Rf为相对频度,为每个样方中林冠层(或林下层)某一物种的频度与样方中所有物种的总频度之比;Rc为相对显著度,为每个样方林冠层(或林下层)某一物种的胸高断面积与样方中所有物种的总胸高断面积之比。Ra、Rf和Rc用百分数表示。
运用植物系统发育数据库Phylomatic version 3(http://phylodiversity.net/phylomatic/),基于Ge-nBank (http://www.ncbi.nlm.nih.gov/genbank)中陆地植物的基因序列构建进化树进行群落谱系多样性和结构分析。使用谱系多样性指数(Faith′s PD)度量每个样方的系统发育alpha多样性[22]。用净种间亲缘关系指数(Net Relatedness Index,NRI)量化每个样方内物种间的谱系关系,NRI衡量种间平均谱系距离(MPD)的标准化效应大小[23]:
NRI=-1×(MPDobs-MPDrand)/sdMPDrand,
式中,MPDobs为MPD的观察值,MPDrand为随机群落的预期MPD(n=999),sd MPDrand是随机群落MPD的标准差。Faith′s PD和NRI利用R 3.5.2中的picante包计算。
1.4 土壤样品采样与分析
在每个20 m×20 m样方中随机设置9个采样点,用内径为8.5 cm的不锈钢土钻采集0-20 cm土层的样品,混合均匀后去除根系、石砾等,过2 mm孔径筛后自然风干,用于土壤理化性质的测定。
采用环刀法测定土壤容重(Soil Bulk Density,SBD),采用重量法测定土壤含水量(Soil Water Content,SWC),采用水土比(2.5∶1)电位法测定土壤pH值,采用重铬酸钾外加热法测定土壤有机碳(Soil Organic Carbon,SOC),采用凯氏定氮法测定土壤全氮(Total Nitrogen,TN),有效氮(Available Nitrogen,AN)采用氯化钾溶液浸提法测定,采用酸溶-钼锑抗比色法测定土壤全磷(Total Phosphorus,TP),采用0.5 mol·L-1NaHCO3法测定速效磷(Available Phosphorus,AP),采用火焰光度法测定土壤全钾(Total Potassium,TK),采用乙酸铵浸提法测定土壤速效钾(Available Potassium,AK)[24]。
1.5 统计分析
采用单因素方差分析(One-way ANOVA)检验同一层次不同林分物种多样性指数、土壤性质以及林分结构(密度、高度、盖度、重要值、生物量)差异的显著性,采用最小显著性差异法(Least Significant Difference,LSD)进行多重比较(P<0.05)。在方差分析之前,所有检验变量的原始数据均使用Shapiro-Wilk检验进行正态性检验,并使用R 3.5.2的Bartlett检验进行方差齐性检验。采用独立样本t检验对同一林分不同层次间上述各指标进行差异显著性检验(P<0.05)。
采用R 3.5.2的vegan包,基于不同林分各层次间的Bray-Curtis距离,用主坐标分析(Principal Coordinates Analysis,PCoA)评估植物群落结构的总体差异。基于Bray-Curtis相异性测度,用置换多元方差分析(Permutational Multivariate Analysis of Variance,PERMANOVA)检验不同林分垂直群落物种组成的差异。
以植物群落组成为响应变量,土壤理化因子和林分结构特征因子为解释变量,用冗余度分析(Redundancy Analysis,RDA)确定影响植物群落不同层次组成变异的主控因素。本研究采用各层次中物种的重要值来表征群落组成。在进行RDA分析前,对植物重要值矩阵进行Hellinger转化,对土壤和林分结构参数进行标准化,以避免单位尺度差异造成的偏差。用vegan包的ordistep函数向前选择能够解释植物群落组成变化的土壤和林分结构变量,获得最优模型。最后,用varpart函数对最优模型中的土壤物理特性、化学特性及林分结构特征因子进行方差分解分析,并用韦恩图展示3类因子的纯效应和共同效应。
2 结果与分析
2.1 森林结构特征
不同林分林冠层和林下层的个体密度、萌芽个体密度和茎干密度存在明显差异(P<0.05,表2)。在林冠层中,沿幼龄林、中龄林到近熟林,个体密度显著增加,而萌芽个体密度和单株平均茎干数显著降低;各林分林下层的3种密度和单株平均茎干数均显著降低。对于幼龄林,林下层3种密度极显著大于林冠层(P<0.01)。近熟林林冠层3种密度值极显著大于林下层(P<0.01)。就中龄林而言,林冠层和林下层3种密度没有显著差异(表2)。无论是林冠层还是林下层,幼龄林的单株平均茎干数都显著高于中龄林和近熟林(P<0.05)。
3种林分林冠层的胸高断面积和地上生物量均显著大于林下层(表2)。从幼龄林、中龄林到近熟林,林冠层的胸高断面积和地上生物量显著增加,而林下层则显著降低。林冠层和林下层的林冠高度均随森林恢复而显著增加。3种林分林冠层的林冠盖度均极显著高于林下层(P<0.01)。在林冠层,林冠盖度在近熟林达到最大值,而在林下层,林冠盖度在中龄林达到最大值。
研究还发现,由幼龄林、中龄林到近熟林,物种丰富度和Shannon-Wiener指数在林冠层和林下层均呈现出先增加后降低的趋势(表2)。在林冠层,中龄林的Shannon-Wiener指数显著高于幼龄林和近熟林(P<0.05),中龄林的物种丰富度显著高于幼龄林(P<0.05),但与近熟林差异不显著。
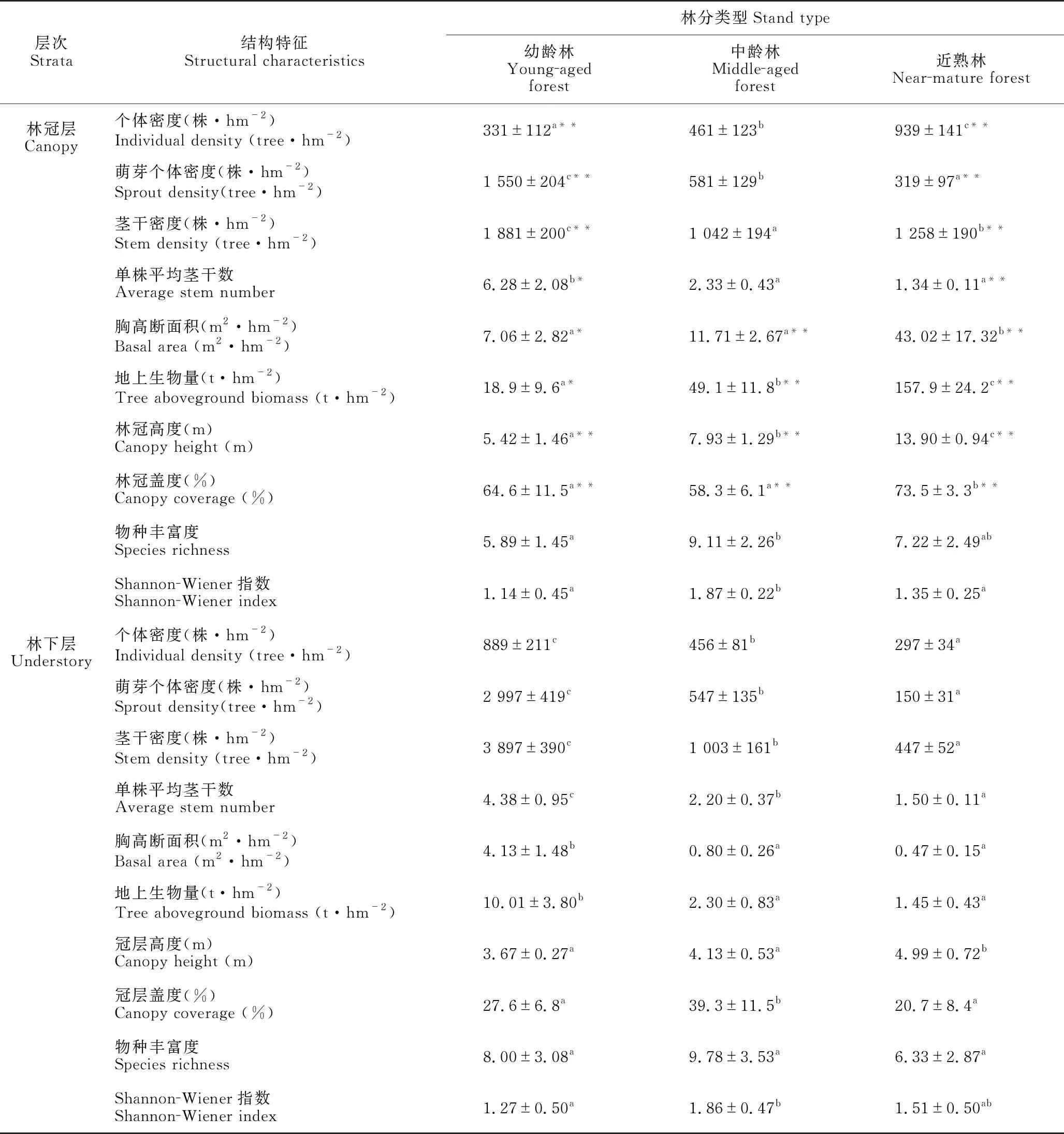
表2 3种林分林冠层和林下层的结构特征
2.2 植物群落组成和谱系结构
PCoA分析(图1)表明,幼龄林的林冠层和林下层群落组成很相似(PERMANOVA,R2=0.238,P=0.076)。第一轴将中龄林林下层与其他群落层次分开,解释了37.77%的变异量;第二轴将幼龄林、中龄林和近熟林的林冠层和林下层分开,可解释的变异量为17.32%。林冠层和林下层Faith′s PD沿幼龄林、中龄林到近熟林均表现出先增大后减小的特征[图2(a)]。随着演替的进行,林冠层和林下层植物群落谱系结构均从聚集(NRI>0)过渡到发散(NRI<0)[图2(b)]。在中龄林,林冠层植物群落结构为谱系聚集,而林下层植物群落为谱系发散。
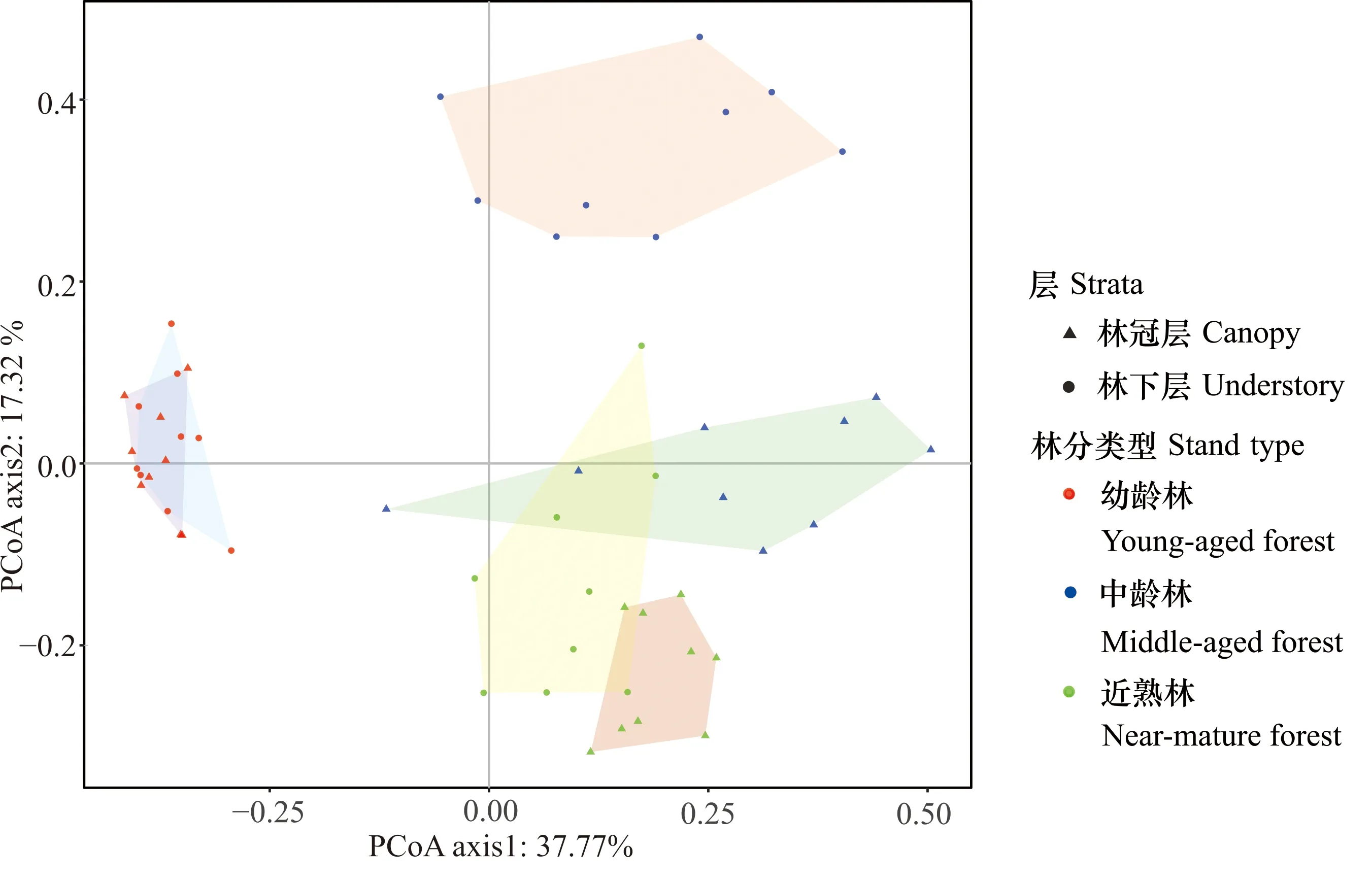
图1 不同林分林冠层和林下层植物群落的主坐标分析

不同小写字母表示不同林分间差异显著(P<0.05)
2.3 植物群落物种组成变化的驱动因子
RDA分析结果表明,随着演替的进行,不同森林林冠层、林下层植物群落物种组成的变化主要受土壤化学性质、土壤物理性质、林分结构特性及其综合效应所驱动[林冠层RDA模型:调整R2=0.458,P=0.001,图3(a);林下层RDA模型:调整R2=0.294,P=0.001,图3(b)]。土壤和林分结构特性解释的变异量在不同层次之间有很大的差异[图3(c)、(d)]。在林冠层中,地上生物量(TAB)、土壤有机质(SOM)、岩石裸露率(RBR)、土壤容重(SBD)和林冠层盖度(CC)是解释方差最多的因素[图3(a),表3]。土壤化学性质、土壤物理性质、林分结构特性分别独立解释了10.7%、2.1%和3.0%的变异[图3(c)],三者的共同解释率为4.1%,而土壤化学性质和林分结构特性的共同解释率为15.7%。在林下层,超过一半(70.6%)的植物群落组成变化未得到解释。SOM、TAB、RBR和CC解释了大部分的方差[图3(b),表3]。土壤化学性质和林分结构特性的共同解释率为11.2%,高于两者单独解释的比例[图3(d)]。土壤因子和林分结构因子的共同解释率为5.4%。RDA结果表明,pH、SOM、TAB、CC、SBD和RBR是林冠层和林下层植物构成的共同驱动因素。
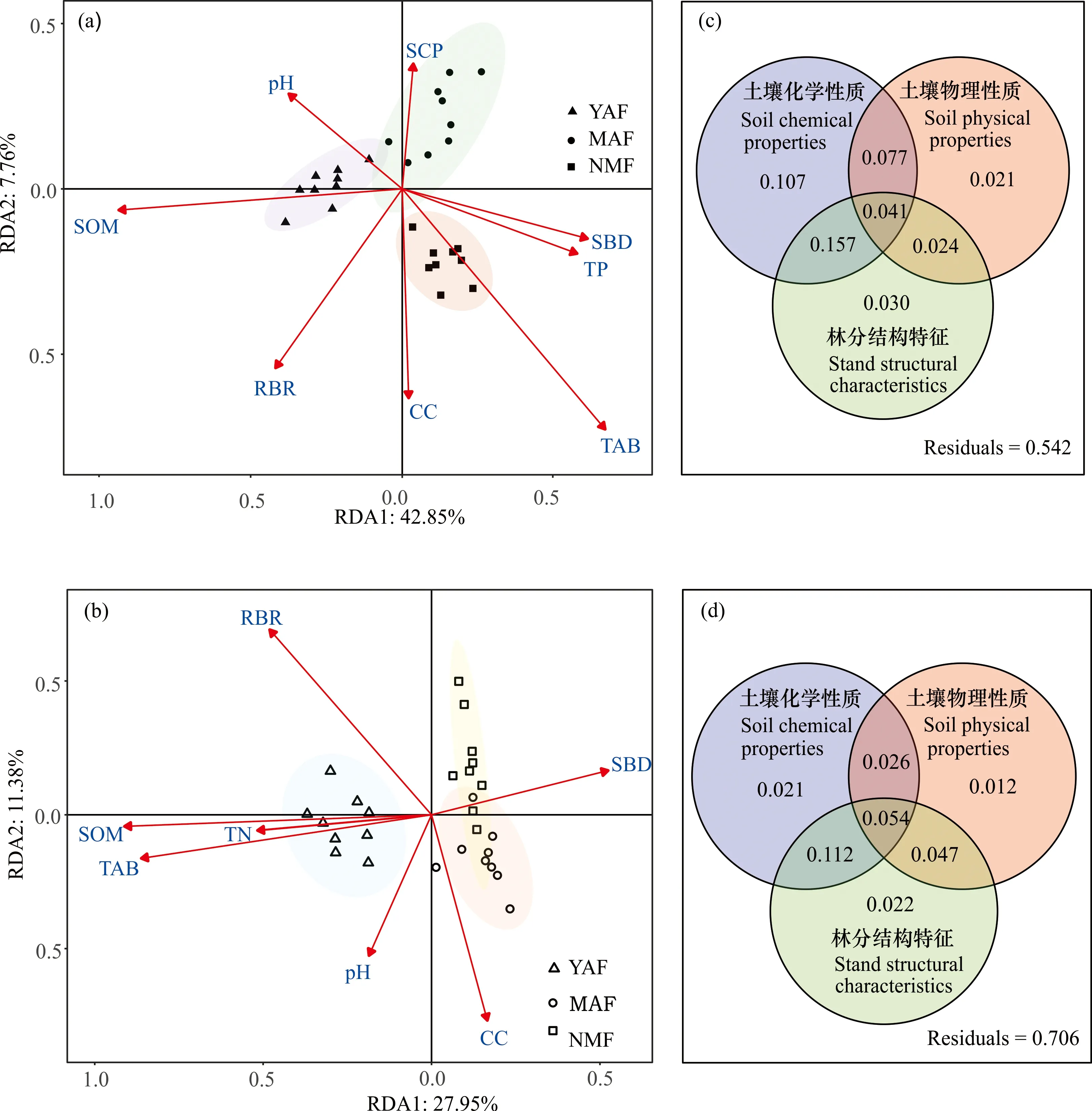
YAF:幼龄林;MAF:中龄林;NMF:近熟林;SOM:土壤有机质;pH:土壤pH;TP:土壤全磷;TN:土壤全氮;SBD:土壤容重;SCP:土壤毛管孔隙度;RBR:岩石裸露率;TAB:地上生物量;CC:冠层盖度。(a) 不同林分林冠层植物组成与土壤因子和林分结构特性的冗余度分析;(b) 不同林分林下层植物组成与土壤因子和林分结构特性的冗余度分析;(c) 土壤因子和林分结构特性对林冠层植物组成影响的方差分解;(d) 土壤因子和林分结构特性对林下层植物组成影响的方差分解
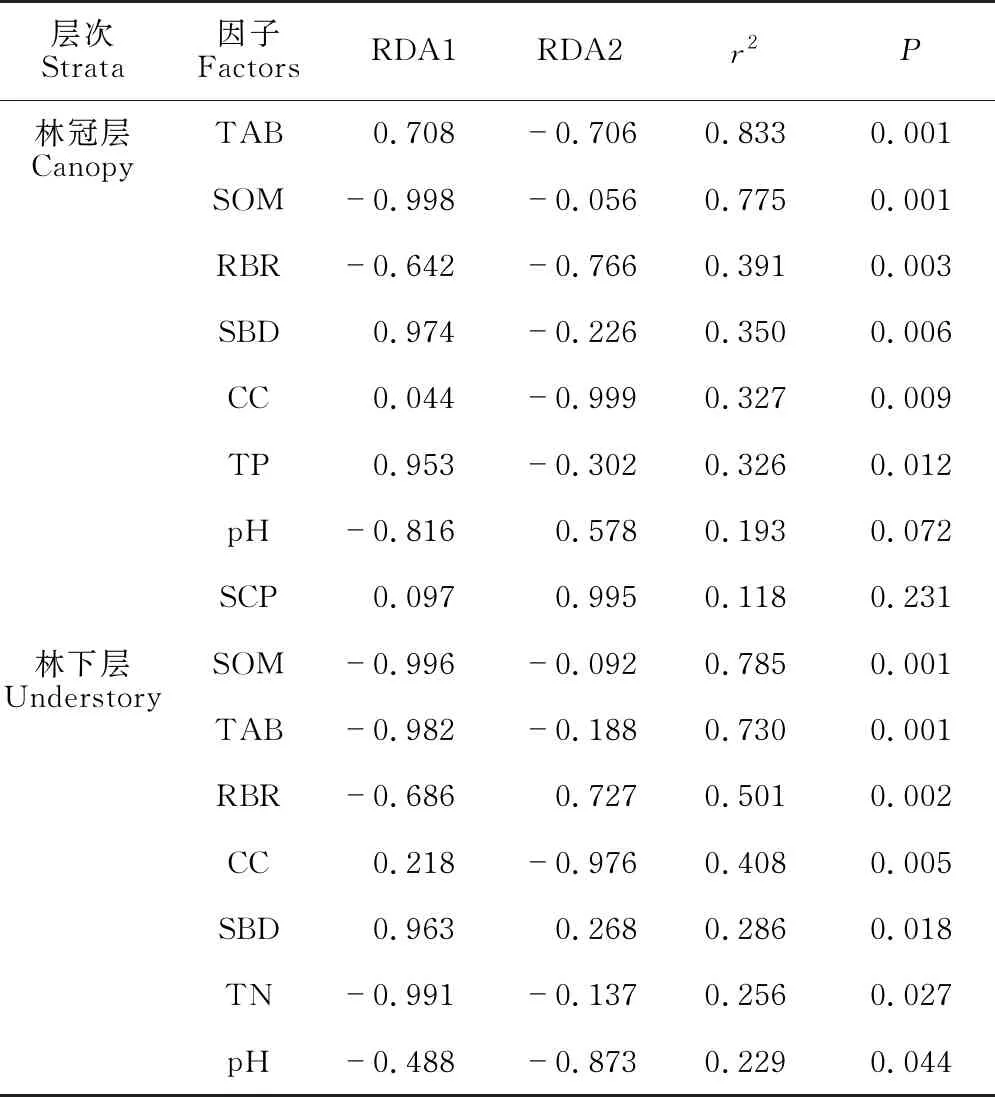
表3 林冠层和林下层土壤、林分结构特性及排序轴的相关性
3 讨论
3.1 不同森林林分结构的变化
本研究表明,由幼龄林、中龄林到近熟林,林冠层和林下层的结构特征之间存在显著差异(表2)。早期研究表明,林冠层和林下层的树木密度和胸高断面积均在演替中期达到峰值,而演替后期森林的密度最小[25]。也有研究发现,树种多样性的峰值可能出现在演替中期[26]或演替后期[27]。本研究中,森林结构指标,如树冠高度、个体密度、胸高断面积和林冠层生物量随森林恢复呈连续性递增,而个体密度、萌芽个体密度、茎干密度、平均茎干数、胸高断面积、林下层生物量沿幼龄林、中龄林到近熟林的恢复序列不断减小。此外,林冠层和林下层的物种丰富度和多样性峰值均出现在中龄林阶段(表2)。这些结果与之前的研究[28]一致,即幼龄林包含更高的萌芽个体和茎干密度,胸高断面积和树冠高度较低,乔木种类少,多样性指数低,以及树种多样性指数的高峰期在中龄林阶段。
萌生是植物对干扰响应的一种重要的适应方式,通常情况下,在遭受严重干扰后,具有较强萌生能力的物种能够存活,反之亦然[28]。可见,萌生过程在植被恢复和自然更新中具有重要作用[29]。有研究表明,除祼子植物外,几乎70%-80%的树种均具有不同程度的萌生能力,但不同的物种其萌生能力的强弱不同[28]。本研究发现,3种森林在萌生特性方面存在显著差异,尤其是幼龄林,其林冠层(6.28±2.08)和林下层(4.38±0.95)的单株平均茎干数最高。幼龄林的高茎干密度可能与以前的土地利用类型(如反复砍伐薪柴)有关。已有研究表明,在岩溶森林[1]和热带森林[30]中,树木受损后萌芽是林分萌生的一种重要形式。此外,在森林再生的早期演替阶段,萌芽可能会导致初始覆盖度的增加[30]。在岩溶地区,停止人为干扰后的一段时间内,茎干可以再次萌芽,但随着演替进程的延续,大部分茎干被单株茎干所替代。本研究揭示的幼龄林高茎干数特征与岩溶地区农业撂荒形成的其他森林[12,13]存在明显差异。这种强烈的再生特性对岩溶地区天然次生林的建立和演替起着重要作用。
3.2 不同森林群落不同层次物种组成的变化
研究表明,3种林分中,林冠层和林下层植物组成存在显著差异。PCoA分析发现,幼龄林林下层和林冠层的群落组成极为相似,而中龄林和近熟林不同林层的物种组成却明显不同(图1)。虽然近熟林具有最高的物种丰富度、Shannon-Wiener 指数和Faith′s PD,但是,林冠层和林下层的物种组成和构建却存在显著差异。这一结果与早期的研究结果[28]一致。早期研究发现,森林中不同层次受不同组装机制的影响[16]。本研究还发现,中龄林林冠层物种组成为谱系聚集,而林下层则为谱系发散,这可能是林冠层经常受到较严酷环境(如光照、水分限制等)的生境过滤,而林下层却具有较有利的生境,这符合压力—优势假说[31]。在近熟林不同林层中,林冠层植物组成在演替后期呈现出谱系发散格局,表明随着天然次生林的恢复,非生物环境胁迫降低,有利于亲缘关系远的物种在群落中共存。
3.3 驱动森林群落不同层次物种组成变化的主要因子
本研究表明,不同群落林冠层和林下层之间的植物组成存在显著差异,这是由土壤和林分结构特征及其相互作用共同驱动形成的。岩溶地区土壤pH值通常较高,钙质土壤含水量与pH值呈正相关[32]。岩石裸露程度也影响到土壤的蓄水能力[33]。土壤容重越低,透水性越强,保水能力越弱[34]。因此,在亚热带岩溶天然次生林中,土壤pH值、岩石裸露率和土壤容重共同成为植物分布的生境过滤器,这与其他研究[35]结果相似。除土壤性质外,乔木地上生物量和冠层盖度是决定森林群落不同层次物种组成差异的最重要因素。这在一定程度上与树木生物量(生物量高表明资源利用效率高和养分回归的凋落物产量高)[36],以及冠层盖度与林下植物的光有效性[37]有关。
本研究中,植被通过在幼龄林中重新萌芽和定居得以恢复。但幼龄林较高的基岩祼露率和较低的土壤容重不利于乔灌木在演替早期的定居,再者,尚无层次分化,导致林冠层与林下层物种组成很相似。在中龄林,随着土壤和冠层性质的改善,为植物提供了更多样的生境。在这一阶段,林冠层植物具有气候主导的生态位,而林下层植物则具有土壤和微生境主导的生态位,导致林冠层和林下层物种组成的差异。进入近熟林阶段,森林环境进一步形成,生境过滤作用减弱,生物因子作用增强,树木地上生物量和冠层盖度成为林冠层和林下层植物组成的主要驱动因素。
