乳脂肪球膜蛋白组成及其功能特性
2022-04-11李贺腾飞
李贺,腾飞
(1.江苏师范大学,江苏 徐州 221116;2.江苏省内皮细胞靶向性心血管药物创制工程研究中心,江苏 徐州 221116;3.蓝河营养品有限公司,长沙 410200;4.Blue River国际羊乳营养研究中心,长沙 410200)
0 引言
乳脂肪球膜(Milk fat globule membrane,MFGM)是由磷脂,鞘脂及膜蛋白组成的3层膜结构,具有抗菌、抗癌、提高免疫力、抗少肌症以及维持正常大脑功能等多种生物学功能[1]。MFGM独特的营养价值除了与MFGM磷脂有关外,还主要与MFGM蛋白有关[1-2]。膜蛋白成分复杂,已鉴定出500余种MFGM蛋白组分,其作用组分及其作用机制尚未完全清晰[3]。微量的MFGM蛋白对维持正常生理功能起到重要的调控作用,是MFGM中最具开发价值的蛋白资源[4-5]。目前,MFGM由于其较高的乳化性和稳定性,以及促进神经认知发育的生物学作用,已被广泛应用于婴幼儿配方乳粉中[6-7]。本文对MFGM蛋白的营养组分和生物学功能方面进行了综述,为乳源MFGM蛋白资源的进一步开发和利用提供理论参考。
1 MFGM的组成结构及分布
乳脂肪是乳中以乳脂肪球(Milk fat globule,MFG)形式存在的一种组成和结构最为复杂的脂质,其含量约占总乳的3%~5%[1]。MFGM主要由膜蛋白、中性脂和极性脂等组成,其含量约占MFG总质量的2.07%~2.99%[8]。膜蛋白和脂质是MFGM的主要组分,分别占MFGM含量30.26%~41.65%和51.69%~66.34%[9]。牛乳、水牛乳以及牦牛乳等哺乳动物乳是婴幼儿配方乳粉的主要原料,MFGM中各组分的含量受物种、地域、泌乳期以及环境等因素影响,如表1所示。MFGM在MFG中的占比不同,其中水牛乳(约2.99%)>牛乳(约2.76%)>骆驼乳(约2.35%)>牦牛乳(约2.11%)>山羊乳(约2.07%);各组分在MFGM的占比也有差异,山羊MFGM中蛋白(约30.26%)含量最低,总脂质(约66.34%)占比最高,骆驼MFGM中水分(约7.88%)含量最高,总脂质(约51.69%)含量最低,牛乳MFGM中极性脂质,如:磷脂酰胆碱(Phosphatidylcholine,PC)、磷脂酰乙醇胺(Phosphatidylethanolamine,PE)和鞘磷脂(Sphingeomyelin,SM)含量相对较高,且磷脂含量和比例与人乳最为接近[10]。

表1 不同乳源MFGM的组成[11]
1.1 MFG和MFGM的形成
MFGM是在MFG分泌过程中形成的一种包裹在MFG表面的三层生物膜,其产生过程如图1所示。在哺乳动物乳腺上皮细胞粗面型内质网膜的作用下合成甘油三酯(Triglyceride,TAG),并作为离散的脂滴释放到细胞质中,被极性脂质和来自内质网的蛋白质等多种成分组成的单层膜包被。细胞内的TAG通过融合成不同尺寸的脂滴,将其转运至细胞的顶端质膜分泌出来[12]。在分泌期间,脂滴完全被细胞双层膜包被,从而在分泌时形成完整的厚度为10~20 nm的三层膜结构[13]。

图1 MFGM分泌示意图[14]
1.2 MFGM的结构
MFGM在乳中含量很低,约占0.08%~0.24%,占总脂肪的2%~6%,占MFG的2.07%~2.99%[9]。MFGM主要由磷脂和蛋白质组成(>90%),还含有少量中性脂质(约5%)、神经节苷脂(约3%)和胆固醇(约2%)等成分[15]。MFGM中的极性脂主要由PC(约35%)、PE,(约30%)、SM(约25%)、磷脂酰肌醇(Phoshhatidylinositol,PI,约5%)及磷脂酰丝氨酸(Phosphatidylserine,PS,约3%)组成[1]。蛋白质和脂质在MFGM中的分布是不对称的,MFGM自内而外,PE、PS以及PI主要位于MFGM内表面,磷脂、软磷脂、鞘磷脂、脑苷脂以及神经节苷脂主要位于MFGM外[13];根据MFGM蛋白的位置可分为:整合型、外周型以及与膜结合松散型蛋白。与膜结合松散的蛋白与甘油三酯具有很强亲和力的脂肪亲脂蛋白(ADPH)位于内侧极性脂单层膜上,黄嘌呤氧化酶/脱氢酶(XDH/XO)位于单层膜内表面,与跨膜蛋白-嗜乳蛋白(BTN)与XO/XDH相邻,并紧密连接形成超分子复合体使内外层膜相连,具有稳定MFGM的作用[10]。MFGM蛋白中的其它高丰度蛋白,如过碘酸稀夫(PAS6/7)和糖基化水平极高的黏蛋白1(MUCI)分别位于MFGM的最外侧和MFGM表面[16]。MFGM整体上呈现为由磷脂、膜蛋白、糖脂以及富含胆固醇与SM的脂筏域等构成的疏松网状结构。

图2 MFGM的结构[8]
1.3 MFGM蛋白的组成及分布
质谱分析技术结合生物信息学分析在蛋白质组成分析和结构预测方面发挥着巨大的作用[1,17]。蛋白质的生物学功能在很大程度上取决于空间结构,因而通过对蛋白质结构进行预测分析,可了解蛋白质的生物学功能,系统地阐明营养补充调节代谢的分子机制[18]。Vanderghem[19]等人基于双向电泳(Two-dimensional gel electrophoresis,2D电泳)分离MFGM蛋白,并结合MALDI-TOF质谱技术,建立了MFGM蛋白在MFGM上的分布模型,研究了7种高丰度MFGM蛋白的组成和分布。由于2D电泳存在对极端等电点、高分子量以及膜结合蛋白鉴定的局限性,姬晓曦[11]等人基于iTRAQ蛋白组学技术研究牛乳、山羊乳、水牛乳、牦牛乳和骆驼乳5种乳源MFGM差异蛋白质组,共鉴定总蛋白336种,通过建立MFGM蛋白差异表达谱,揭示了不同乳源MFGM蛋白的组成及潜在的生物学功能。杨梅[20]等人基于SDS-PAGE和LC-MALDITOF技术分析人乳和牛乳MFGM差异蛋白质组,共鉴定MFGM蛋白315种,人乳和牛乳中特异性MFGM蛋白分别为1230种和173种;基于Gene Ontology(GO)注释功能分析,人乳中37%的MFGM蛋白能够参与代谢相关的生物学过程;55%的MFGM蛋白生物学功能与结合作用有关;34%的MFGM蛋白组成与细胞器生物合成有关。Lu[21]等人基于不同乳源MFGM差异蛋白质组共鉴定出MFGM蛋白,如图3所示:人乳(312种),牛乳(554种),羊乳(175种)以及牦牛乳(143种),其中与人乳MFGM共有蛋白分别为:牛乳(166种)>羊乳(95种)>牦牛乳(76种)。在人乳、牛乳,山羊乳以及牦牛乳MFGM中特异性蛋白数分别为137种,317种,23种和16种。骆驼乳和牦牛乳由于地域和分布的局限性产量很低,与羊乳相比,牛乳中MFGM蛋白种类丰富且更接近人乳,是摄取MFGM蛋白的最佳来源[1]。

图3 人、牛、山羊和牦牛MFGM中蛋白质的维恩图[1]
综上,MFGM蛋白质组成复杂,在不同物种之间具有高度的差异性。就MFGM蛋白组成而言,与羊乳相比,牛乳中MFGM蛋白与人乳之间相关性更高;就分子功能和GO分析而言,人乳MFGM富含内质网蛋白,而牛乳MFGM富含质膜蛋白,参与最多的生物学过程包括:细胞过程、定位、转运以及信号转导等。虽然MFGM蛋白的分析手段不同,可鉴定的蛋白质数量差异较大,但是无论基于何种测定方法,均证明分子量分布在47~225 ku范围内的7种蛋白,如图4所示:MUCI(160~200 ku)、XDH/XO(150 ku)、PAS III/IV(约98 ku)、CD36(76~78 ku)、BTN(68 ku)、PASⅥ/Ⅶ(45~52 ku)以及GTP结合蛋白(28~38 ku)为MFGM蛋白中的高丰度蛋白,并且MFGM的许多生物学功能,如乳化性、稳定性、增强免疫力以及与神经认知发育等与这几种蛋白有直接关系[2,9]。
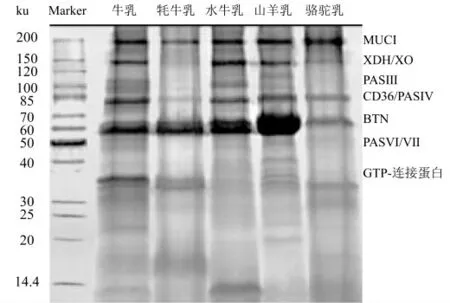
图4 不同乳源MFGM蛋白的SDS-PAGE图[22]
2 MFGM蛋白的性质
MFGM蛋白不仅是乳中最重要的营养成分,而且在乳制品加工过程中具有维持乳制品的胶体特性、泡沫性、稳定性以及稳定离子和矿物质结构等有重要作用[19,23]。MFGM蛋白因其在免疫调节,肠道健康以及改善认知能力上发挥功能作用,将其作为天然的乳化剂和活性成分添加至婴幼儿配方乳粉中成为新型婴幼儿配方乳粉新的研究热点[14,24]。因此,准确认识MFGM蛋白的乳化性、稳定性、消化性以及安全性和耐受性对MFGM蛋白相关功能产品的开发具有重要意义。
2.1 MFGM蛋白的乳化性和稳定性
MFGM是一种天然乳化剂,MFGM中磷脂-蛋白层作为表面活性剂,能防止脂肪球的聚集和絮凝,维持乳体系中水-油相以稳定形式存在[13,19]。MFGM的稳定性受脂肪球大小、极性脂在脂肪球表面的覆盖率、膜蛋白在膜上的定位和脂肪球表面发生的化学反应等多种因素影响。据报道在婴儿配方奶粉中添加1.6%的MFGM,可显著降低脂肪球的粒径,提高脂肪的物理性质和氧化稳定性[25]。在相同浓度条件下,与MFGM磷脂相比,MFGM蛋白制备的水包油型乳化剂粒径更小,ζ-电位相对较高[2,26]。由于MFGM中的特异性蛋白对乳体系稳定性的作用相对更大,因此MFGM蛋白在乳液的稳定性中得到广泛的研究和应用。MFGM蛋白具有亲水亲油性,能够吸附到液滴表面。在相同的浓度条件下,与乳中的其他优质蛋白相比(如:酪蛋白和乳清蛋白),由于MFGM蛋白分子量较大,使其乳状液表面覆盖率高,更易于吸附在水-油界面上,形成一层黏弹性吸附膜,产生静电斥力,阻止聚集,增强乳液稳定性[25,27]。由于MFGM蛋白的种类丰富,且多数为膜结构紧密的整合蛋白,其乳化作用是基于何种蛋白组分尚未明确[3]。
2.2 MFGM蛋白的消化稳定性
MFGM具有生物活性的前提条件是其在胃肠道消化过程中的稳定,MFGM在胃肠道运输过程中可能以多种形式存在,MFGM周围的碳水化合物可以影响MFGM蛋白质的消化性[2]。Le[23]等人通过SDSPAGE结合考马斯蓝和高碘酸希夫两种染色方法,评估MFGM体外模拟消化稳定性并通过质谱技术对消化液中多肽进行定性和定量分析。与非糖基化蛋白相比,糖蛋白对内源蛋白酶抗性最高;MUCI消化抗性最强,经胃肠道消化系统作用后,仍能够检测到部分MUCI以完整的蛋白形式存在;CD36、XO和PAS6/7在胃肠道消化性具有较强的抗性,在未去除MFGM脂质的情况下,脂质可以保护MFGM糖蛋白免受胃肠道消化,如:PAS6/7经过胃肠道消化后,绝大部分蛋白以完整的形式存在[25,27]。因此,还需进一步深入研究MFGM糖蛋白在胃肠道中的稳定性,以期探索其对人体健康的调控作用。
2.3 MFGM蛋白的安全性和耐受性
MFGM极性脂乳化的脂滴超微结构更接近于母乳,且消化过程更符合婴儿的脂肪消化途径[14,28]。特别是对于脏器功能发育不全和免疫力较低的婴幼儿而言,富含MFGM的配方粉能够缓解喂养不耐受问题,促进肠道有益菌群增殖,提高钙和镁的吸收率,能满足婴幼儿能量需求,对各器官的健康发育具有促进作用。Breij等人[28]基于223名婴儿临床研究发现,喂养含有人工模拟MFGM成分和结构的配方乳粉未发生与腹泻和呼吸道相关的不良现象,婴儿生长状况与人乳喂养无显著性差异;此外,基于1500名孕妇临床研究发现,在妊娠前后无论是否摄入复杂的MFGM磷脂都能够满足胎儿的生长。Billeaud[24]等人给予出生14 d的新生婴儿,喂养标准配方乳粉、富含MFGM磷脂乳粉以及富含MFGM蛋白的乳粉4个月,研究发现MFGM磷脂和MFGM蛋白组婴儿的体脂比、代谢标志物和免疫标志物之间无显著性差异,婴儿对MFGM磷脂和蛋白是耐受的,在婴幼儿配方粉中添加MFGM磷脂和蛋白是安全的,且能促进婴儿的生长发育。此外,Li等人基于体外细胞实验证明了MFG-E8作用浓度低于200μg/mL时,对Caco-2细胞不具有细胞毒性[29];根据凝集素在临床上靶向药物的用量,以及人体用量和大鼠之间药物等效剂量比值系数计算,确定并证明了16 mg MFG-E8能够有效促进大鼠腓肠肌的修复和再生,用量是安全和耐受的[30]。
3 MFGM蛋白的功能特性
牛乳是人乳的主要代用乳,在世界上产量最高[1]。尽管MFGM的大多数有益作用与极性脂质有关,但近年随着MFGM功能性蛋白研究的不断增加。MFGM蛋白的营养组分及其在健康中作用的相关研究已不再局限于细胞和动物实验,已广泛开展了临床研究[6-7]。牛乳MFGM蛋白中含有MUCI、XDH、CD36、MFG-E8以及BTN等多种生物活性蛋白,赋予了牛乳MFGM抗癌、增强免疫力、降低胆固醇、改善血脂、修复肠道、增强骨骼肌质量和功能以及促进神经认知发育等功能,使牛乳MFGM蛋白兼具营养和保健的双重功能[4,31]。
3.1 抗癌
有关MFGM蛋白的抗癌活性最早见于2002年,Vissac发现并证明了乳腺癌易感基因1(BRCA1)和乳腺癌易感基因2(BRCA2)是MFGM蛋白中具有抗癌功能的活性蛋白,能够参与DNA的修复过程[32]。随后研究发现,MFGM蛋白中低剂量的脂肪酸结合蛋白(FABP)可通过抑制癌细胞的生长,起到抗癌作用,但是相关作用机制尚未明确[3,13]。MFGM蛋白中的β-葡萄糖苷酸酶可以参与肠道葡萄糖醛酸苷的降解,形成葡萄糖苷酸,中和肝脏细胞中会刺激结肠癌形成的毒性物质,进而对结肠癌发生和发展起到预防和缓解的作用[3,31]。近年来,随着蛋白质组学技术的发展和对MFGM蛋白研究的深入,Li[3]等人采用体外热处理和特异性水解来修饰MFGM分子和超分子结构,研究发现酶解的MFGM对HT29细胞活性显著降低,与热处理之间存在关系,并提出MFGM中不仅磷脂具有抑制HT29细胞活性,MFGM蛋白也同样具有显著的抗结肠癌作用。姬晓曦[22]等人基于蛋白质组学技术分析了5种不同乳源MFGM蛋白组成,研究外源性添加MFGM蛋白对HT-29结肠癌细胞和MCF-7乳腺癌细胞的抑制作用,揭示了MFGM蛋白能够通过介导线粒体凋亡信号通路促进HT-29细胞和MCF-7细胞凋亡,并认为MFG-E8、长链酯酰辅酶A合成酶、三磷酸腺苷结合转运蛋白G、糖基化依赖细胞黏附分子1是MFGM蛋白中促进细胞凋亡的的主要功能成分。其作用机制为MFGM蛋白通过引起细胞线粒体膜电位下降,触发线粒体介导的凋亡途径,细胞内抗凋亡因子Bcl-2表达量下调,促凋亡因子Bax表达量上调,细胞膜稳定性下降,通透性增加,细胞膜形成孔隙并释放细胞色素C及其他凋亡因子于细胞质中;进而导致Caspase-9活化,上调Caspase-3表达量,从而促进细胞凋亡。
3.2 增强免疫力
MFGM蛋白中的XDH、MUCI、MFG-E8以及FABP具有抑制F4ac阳性大肠杆菌对猪肠上皮细胞的黏附作用[13,19]。过氧化氢被认为是牛奶中乳过氧化物酶系统的底物,XDH对细菌结合没有保护作用,但是XDH可通过形成过氧化氢抑制细菌分泌,抑制大肠杆菌和肠炎沙门氏菌的生长[3]。MUCI是由MUCI基因编码的能够与膜结合的上皮黏蛋白,是膜相关黏蛋白以及膜结合黏蛋白[1,33]。由于其含有大量的唾液酸使乳腺上皮载有负电荷,能够阻止乳腺导管内壁的上皮细胞粘连保持开放状态,参与上皮细胞调控过程[33]。体外抑制病原体生长的能力证明了150μg/L MUCⅠ和MUC4可有效抑制肠炎沙门氏菌SL1344的侵袭,其作用效果显著优于高浓度(约1000倍,含有150 mg/L的MUCⅠ和MUC4)的牛血清白蛋白、全脂牛奶、脱脂牛奶以及MFGM[34]。此外,与人乳和羊乳相比,牛MFGM富含抗菌蛋白,因此,相对于人乳和羊乳增强免疫力更具优势[1]。
3.3 促进肠道发育
MFGM蛋白中MUCⅠ、MFG-E8、XDH生物活性成分,已被证明可以维持肠道屏障的完整性。Ying[35]等人基于大鼠实验评估MFGM对短肠综合征大鼠肠屏障功能的保护作用,通过对小肠切除的大鼠补充MFGM,其肠组织中MUCⅠ的表达量增加,下调NLRP3和Caspase-1的表达,降低白细胞介素-1β(IL-1β)的水平,抑制NLRP3炎性体信号通路激活增强大鼠肠屏障完整性[33,35]。MFG-E8表达量的变化是评价溃疡性结肠炎的重要指标之一,是肠道内环境平衡的主要营养因子。MFG-E8在促进肠道发育和维持肠道稳态中的作用主要表现为:①激活PKC信号通路促进肠上皮细胞迁移,增强损伤上皮细胞的修复功能[35-36];②识别凋亡细胞表面外翻的磷脂酰丝氨酸“eat me”信号,将凋亡细胞与巨噬细胞表面上的αvβ3整联蛋白连接,增强巨噬细胞对凋亡细胞的吞噬作用,维持肠道内环境稳态[36];③抑制核转录因子-κβ(NF-κβ)的转录活性,下调促炎因子TNF-α和IL-1的表达,缓解肠道黏膜炎症[35-36]。
3.4 改善血脂
牛乳中MFGM蛋白对脂质代谢的影响可能与其活性成分有关,CD36是维持脂质体内平衡所必需的,其缺乏可能导致血脂异常的发生[8,37]。在甘油三酯合成过程中,抑制FASN,ACSL1和ACACA的表达能够减少脂肪生成,有助于治疗系统性和组织性高血脂症[3]。不同乳源MFGM蛋白组成和性质上存在显著性差异,TMEM120B、ACSS2、ApoC3和ERLIN2是牦牛乳MFGM中的高丰度蛋白,其中ERLIN2蛋白通过调控固醇调节元件结合蛋白-1c(SREBP-1c)mRNA和蛋白的表达,稳定胆固醇的含量,改善脂质代谢过程,减少脂质滴的形成[3];TMEM120B、ACSS2和ApoC3能够诱导脂肪细胞的分化,促进甘油三酯的形成[3,38]。ACADVL是牛乳MFGM中的特异性蛋白,可降低体内脂质堆积;相反,MFGM中的特异性蛋白RETN和LBR能够诱导胆固醇合成和脂质堆积[3]。此外,与牛乳相比,牦牛乳MFGM蛋白具有较高的减低脂质积累的作用可能是由于其ERLIN2含量高于牛MFGM蛋白中ACADVL的含量所导致的。
3.5 预防少肌症
少肌症是一种随着年龄增长而导致骨骼肌质量、力量以及功能衰退的老年退行性疾病[39]。在老年人群中发病率极高,其中60~70岁人群的发病率为5%~13%,80岁以上发病率高达50%以上[7,30]。Harii[6-7]等人基于32名健康成年男性和女性为临床研究对象,通过外源性补充MFGM结合抗阻力运动,证明了补充MFGM可以逆转肌肉质量和功能,且对人体是安全的。Harii[40]基于小鼠实验证明了MFGM能够促进少肌症小鼠骨骼肌神经肌肉接头合成的功能性因子Dok-7和肌肉特异性激酶(MuSK)的mRNA表达,促进肌肉质量增加;Li[30,41]等人基于体外成肌细胞实验和大鼠体内实验证明了MFG-E8能够结合表皮生长因子受体,通过激活PI3K/Akt信号通路促进细胞增殖与分化促进成肌细胞增殖。关凯方[42]等人基于体内大鼠和体外成肌细胞实验证明了MFG-E8能够通过激活/ERK信号通路介导细胞周期关键调控因子Cyclin D1、Cyclin E1及CDK2的表达,增加线粒体数量,促进成肌细胞增殖,从而缓解肌肉萎缩,达到修复受损肌纤维的目的。
3.6 促进神经认知发育
母乳是婴幼儿营养的黄金来源,与母乳相比,配方乳粉中缺乏与神经发育相关的重要乳源功能因子[1,28]。随着婴幼儿配方乳粉的升级换代,MFGM作为一种油-水-界面物质,能够参与脂肪的消化、吸收和代谢,促进婴儿神经系统和免疫系统发育,提高婴幼儿的认知和免疫功能[28]。国内外大型乳企相继推出了富含MFGM成分的第五代婴幼儿配方乳粉,其脂肪球结构与母乳更相似,与普通配方乳粉相比,富含MFGM配方乳粉脂肪的消化和餐后脂质代谢水平更优,在神经发育上更接近母乳[14,28]。MFGM中的唾液酸(来自神经节苷脂)、鞘磷脂和胆碱是促进神经发育的重要组分。目前,临床研究发现健康的婴儿补充含有MFGM蛋白含量(4%)的配方乳粉,其认知能力评分相对配方奶粉更高、更接近母乳水平,因此提出MFGM蛋白同样具有促进婴幼儿神经认知发育的作用,但是有关其活性蛋白组分还有待进一步研究。近年来,MFG-E8在缓解老年神经退行性疾病(如阿尔茨海默氏病、中风以及脑外伤)中的研究受到广泛的关注。MFG-E8在预防和治疗阿尔兹海默症方面的作用主要表现为:①识别细·胞上的磷脂酰丝氨酸信号,锚定到细胞表面整合素αvβ5,启动细胞内CrkIIDOCK180-Rac1信号通路,激发吞噬细胞中的细胞骨架重排,参与吞噬凋亡细胞和潜在毒性物质,减少神经炎症,维持大脑正常生理功能[43-44];②介导PI3K,NF-κβ和信号通路级联反应抑制小胶质细胞外Aβ过表达[36];③逆转Aβ1-42诱导的M1促炎因子标记物(如:TNF-α、IL-1β和CD86)和M2抗炎因子标记物(如:Arg-1、IL-10和CD206)的表达,介导NF-κβ和PI3K/Akt信号通路,促进M2型小胶质细胞的积累和抑制M1型小胶质细胞的产生,调节Aβ过表达引起的脑内神经毒性[36,45]。
4 结束语
多年来国内外学者已对乳源MFGM的营养特性和加工特性进行了深入研究,并应用于婴幼儿配方乳粉中。近年来,由于MFGM中活性蛋白种类丰富,很多活性蛋白仅需微小剂量就能够对人体生理起到至关重要的调控作用,因此越来越多的MFGM蛋白及其在体内的作用机制被不断地挖掘和研究。牛乳是人们日常营养来源的健康乳品,与羊乳和骆驼乳MFGM蛋白相比,牛乳MFGM蛋白种类丰富,与人乳更接近;与大豆蛋白、酪蛋白以及乳清蛋白等其他优质动植物蛋白相比,微量的MFGM蛋白即可起到预防疾病发生和发展相似的作用效果。因此,乳源MFGM蛋白具有重要的开发和应用价值。
