Screening strategies for quorum sensing inhibitors in combating bacterial infections
2022-04-07LnLuMingxingLiGuojunYiLiLioQingChengJieZhuBinZhngYingyingWngYongChenMingZeng
Ln Lu,Mingxing Li,Guojun Yi,Li Lio,Qing Cheng,Jie Zhu,Bin Zhng,Yingying Wng,Yong Chen,Ming Zeng
aAntibiotics Research and Re-evaluation Key Laboratory of Sichuan Province,Sichuan Industrial Institute of Antibiotics,School of Pharmacy,Chengdu University,Chengdu,610000,China
bLaboratory of Molecular Pharmacology,Department of Pharmacology,School of Pharmacy,Southwest Medical University,Luzhou,646000,Sichuan,China
Keywords:
Quorum sensing inhibitor
Quorum quenching
Anti-infective agent
Screening strategies
A B S T R A C T
Interference with quorum sensing(QS)represents an antivirulence strategy with a significant promise for the treatment of bacterial infections and a new approach to restoring antibiotic tolerance.Over the past two decades,a novel series of studies have reported that quorum quenching approaches and the discovery of quorum sensing inhibitors(QSIs)have a strong impact on the discovery of anti-infective drugs against various types of bacteria.The discovery of QSI was demonstrated to be an appropriate strategy to expand the anti-infective therapeutic approaches to complement classical antibiotics and antimicrobial agents.For the discovery of QSIs,diverse approaches exist and develop in-step with the scale of screening as well as specific QS systems.This review highlights the latest findings in strategies and methodologies for QSI screening,involving activity-based screening with bioassays,chemical methods to seek bacterial QS pathways for QSI discovery,virtual screening for QSI screening,and other potential tools for interpreting QS signaling,which are innovative routes for future efforts to discover additional QSIs to combat bacterial infections.
1.Introduction
The emergence of antimicrobial resistance constitutes a major global health care concern and poses a significant challenge.Conventional antibiotic therapies targeting the synthesis of protein,DNA,RNA,and folic acid usually exert bacteriostatic or bactericidal effects on multiple targets,resulting in strong selective pressure on bacterial communities and subsequently giving rise to bacterial strains resistant to conventional antibiotics[1-5].In response to this,alternative strategies to antibiotics for bacterial infections are generating significant interest.Among these,some innovative alternative therapies,such as quorum sensing(QS)disruption,have been intensively studied[6-10].In terms of the extensive impacts on microbial physiology induced by QS,interference with QS has been demonstrated to be a promising approach to decreasing bacterial virulence and restoring antibiotic effectivity by controlling biofilm production,paving the way for future anti-infective treatment and dressing/coating agents in medical applications[11,12].
In the 1970s,Nealson found that the density of Vibrio fischeri(V.fischeri)and Vibrio harveyi(V.harveyi)was positively correlated with their bioluminescence,and confirmed that this phenomenon is controlled by the QS system in bacteria(first described as QS)[13].QS is an intercellular chemical communication process in a cell-density-dependent manner in which bacteria coordinate the expression of QS-mediated genes based on the exchange of small signaling molecules defined as quorum sensors or autoinducers(AIs).
Chemically,QS is based on the synthesis,sensing,and uptake of AIs[14].Once a particular threshold concentration of bacteria is reached,programmed changes that coordinate biological effects including biofilm formation,virulence secretion,swarming ability,sporulation,and protease production are motivated in a densitydependent manner.The signal molecules related to clinical pathogens have been reported and are listed in Table 1 and Fig.1:1)acyl homoserine lactones(AHLs),which exist in most Gram-negative bacteria except Vibrio harringiensis and Mucoccus flavus;2)autoinducer peptides(AIPs),which are produced by most Gram-positive bacteria;3)autoinducer-2(AI-2)furanones,which are utilized by more than 40 species of Gram-positive and Gramnegative bacteria for communication and transmission[15,16];4)autoinducer-3(AI-3),which is used by enterohemorrhagic Escherichia coli.Although the AI-3 structure is still not finely characterized,it has been found that epinephrine or norepinephrine could be considered as an alternative to AI-3;and 5)other signal molecules such as the Pseudomonas quinolone signal(Pqs)and ketopyridazines,which also play important roles in intraspecific QS.Some biosynthetic pathways regulating the generation of important QS signal molecules such as AHL and AI-2 involve numerous receptors and enzymes,which are summarized as:S-adenosyl methioninedependent methyltransferase,LuxS,AHL synthase,LuxR receptor,histidine kinase response regulated LuxN protein,and AI-2(LuxP and LsrB)receptor.For example,LuxI/LuxR are key regulatory factors in AHL-mediated QS signaling pathways.The AHL synthase(LuxI type)synthesizes AHL derivatives to generate QS signal molecules.The synthesized AHL entities subsequently diffuse in the adjacent environment and accumulate in a cell-density-dependent manner(Fig.2)[17].When the AHL level reaches the threshold,the AHL interacts with the LuxR-type receptor to activate target gene transcription in QS signaling pathways[18,19].
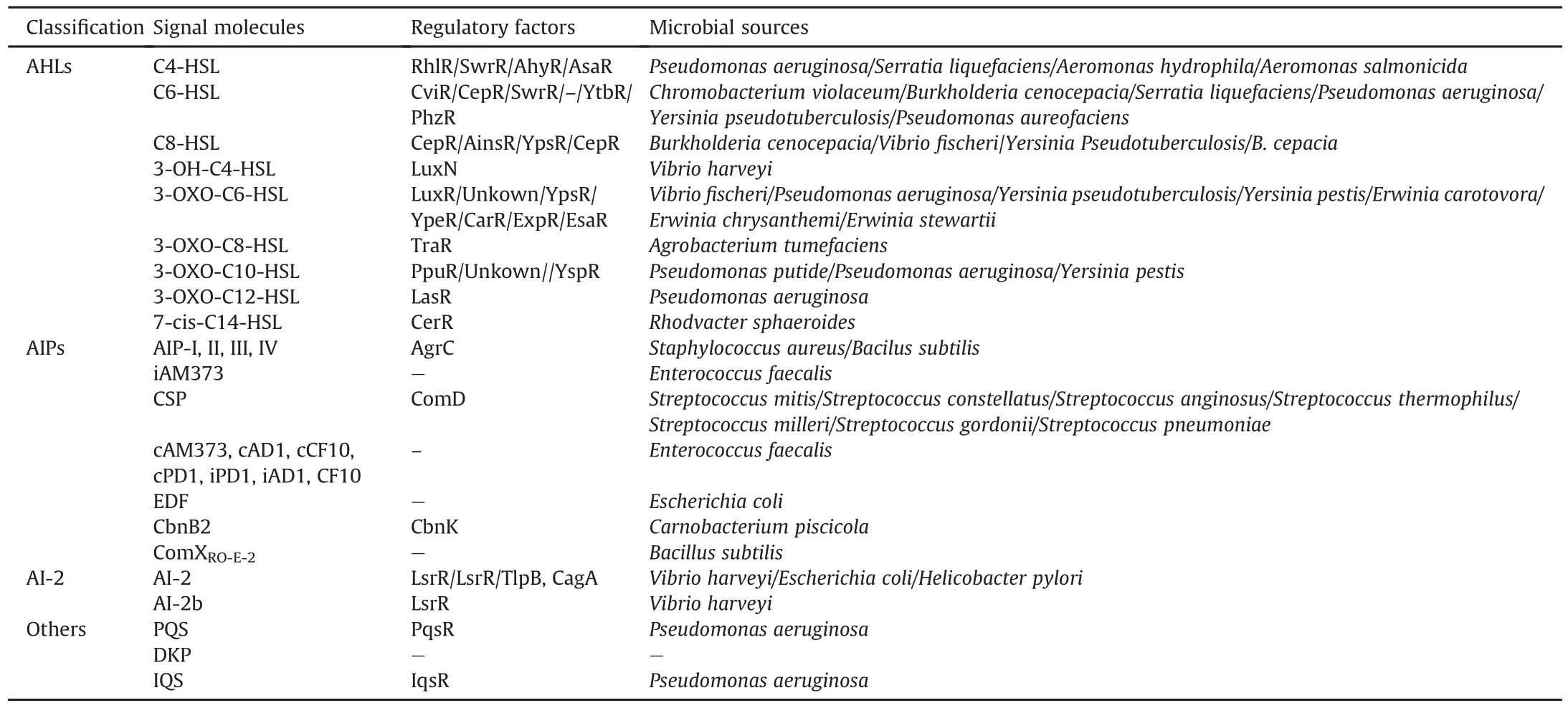
Table 1Classification of autoinducer molecules.
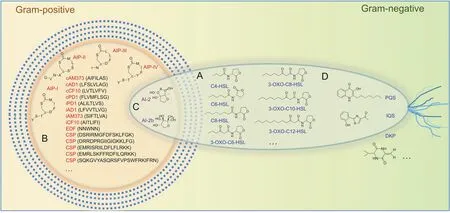
Fig.1.Representation of AI molecules.(A)Acyl homoserine lactones;(B)autoinducer-2 furanones;(C)autoinducer peptides;(D)others.AIs:autoinducers;AIPs:autoinducer peptides;HSL:homoserine lactone;PQS:quinolones;DKP:diketopiperazines;IQS:integrated quorum sensing signal.
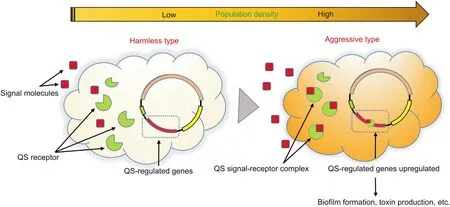
Fig.2.Cyclic process of signal molecules.QS:quorum sensing.
In Gram-negative bacteria,many QS circuits are based on the LuxR-LuxI homologs of V.fischeri that use AHL signal molecules.For example,Pseudomonas aeruginosa(P.aeruginosa)has two key QS systems,LasR-LasI and RhlR-RhlI,which make use of 3-oxo-C12-homoserine lactone and C4-homoserine lactone(HSL)as signal molecules,respectively.The QS is also mediated through the Pqs system,which senses Pqs(2-heptyl-3-hydroxy-4-quinolone).These three systems are hierarchically presented,because the Las system is at the top and LasR controls Rhl and Pqs QS circuits.The Pqs system,which intervenes between the Las and Rhl systems,can interfere with the levels of Las-and Rhl-controlled genes.The Las and Rhl systems coordinate various virulence factors,including exoproteases,siderophores,toxins,and biofilm formation,and apparently exert an influence on host immune systems.
Interference with QS,termed quorum quenching(QQ),was first discovered as an enzyme capable of degrading AHL signals to interrupt QS signaling in Erwinia carotovora[20].Specifically,QQ can be achieved by preventing bacteria from producing or perceiving AIs using quorum sensing inhibitors(QSIs)screened by different means[21],eliminating AIs by QQ antibodies,or extracellular enzymatic hydrolysis of AIs by QQ enzymes[22].To date,many QQ enzymes,QSIs,and QQ antibodies have been reported and extensively reviewed[18,23-25].Because QQ enzymes or antibodies can act remotely and independently without entering the bacterial cells to degrade AHL signals,they are probably less resistance-prone than QSIs,which have to interact with an intracellular target or a receptor on the bacterial outer membrane.However,stability remains a major constraint that usually limits the application of QQ enzymes and antibodies.Moreover,most QQ enzymes discovered such as lactonase are limited in quantity and variety[26].Compared with that of QQ enzymes,the biochemical nature of purified QSIs,originating from a host of organisms including microbes,plants,fungi,and animals in ecosystems,is highly diverse[27].For the discovery of QSIs,diverse approaches exist in-step with the scale of screening as well as specific QS systems.Among them,basic activity-based screening strategies consist of screening organisms,cells,and chemical libraries by QS bacterial reporters as well as QS-based phenotypes,including biofilm formation and virulence secretion.However,these natural QSIs may present a weak efficiency or possible toxicity in the targeted environment.A promising solution to overcoming these limitations is most likely to chemically design and synthesize molecules according to natural QSIs.Additionally,alternative strategies involve structure-based screening for target-oriented discovery of QSIs as well as other novel biotechnical tools for interpreting QS signaling.Overall,different methods,relying on the scale of screening as well as specific QS systems,are necessary for conducting QSI screening,which benefits the discovery and utilization of bioactive QSIs and validation of the effects of some QSI candidates.This review highlights the recent findings in strategies and methodologies for QSI screening,which are promising routes for QSI discovery to control bacterial biofilm formation and virulence to combat bacterial infection.
2.Activity-based screening with bioassays
In activity-based screening with bioassays,the detection systems rely on the testing signal generated by certain reporter proteins involved in colorimetric,bioluminescence,chemilumin escence,and fluorescence effects.Regarding QS bacterial reporters,two approaches are currently being applied,including those relying on the natural QS molecule-triggered responses(Fig.3A)and by constructing engineered strains based on genetic modification(Fig.3B).

Fig.3.Activity-based screening with bioassays.(A)The construction of engineering strains based on the naturally observed activity;(B)The construction of engineering strains based on genetic modification.The detection signal produced by QS is observed via colorimetric, fluorimetric,bioluminescent,and chemiluminescent analyses.
2.1.Natural indicator strain for QSI screening
The easiest approaches to potential QSI screening involve colorimetric tests with natural indicator strains such as violacein formation in Chromobacterium violaceum(C.violaceum),bioluminescence in Vibrio Ferdinans and Vibrio Harringtonii,the prodigiosin produced by Serratia marcescens,and the orange promethazine pigments produced by Staphylococcus aureus(S.aureus)(Fig.3A)[28].More specifically,C.violaceum,which produces the visible purple pigment violacein regulated by the CviI/CviR system,is a frequently used sensor strain for QSI screening.In addition,V.harveyi is a typical biosensor that regulates an easily observed QS-mediated phenotype termed bioluminescence and has wellcharacterized QS networks.Recently,it was reported that metabolite extracts of Gram-positive bacteria from diverse marine environments inhibit the bioluminescence produced by V.harveyi.
These studies usually started with an initial screen using an indicator strain or the combined tests against two QS reporter strains to estimate the anti-QS activity of candidates[29-33].Analysis of QS-controlled phenotypes,such as the virulence factor assay and biofilm formation assay,has significantly decreased the potential candidates.
However,quantitative analyses of bioluminescence and violacein interference do not seem to adequately demonstrate that these inhibitors specifically disrupt QS signaling pathways.Molecular observations,including gene expression and in silico molecular docking,as well as biochemical tests of inhibitor agents,will be needed to explore the mechanisms and action modes of the inhibitory effects.
Notably,for the preparation of extracts for QSI screening,chromatography analysis by liquid chromatography-mass spectrometry or high-performance liquid chromatography(HPLC)fractionation is utilized for the extraction or purification of anti-QS compounds.It is also expected that the extraction approach used for QSI isolation may not be optimal for all types of inhibitory compounds.In some cases,metabolites with high hydrophilicity may not be fully soluble in ethyl acetate.In addition,producing fermentation extracts or isolating metabolites for further biological evaluation is often a laborious process.
It has been found that many microbes generate metabolites or fermentation extracts that regulate QS-dependent phenotypes.In this regard,co-cultivation assays offer a rapid screening approach to testing bacteria that are chemically innovative for QSI discovery.In one instance,an Halobacillus salinus isolate C42 isolated from a co-cultivation assay termed Membrane Overlay was discovered to generate N-(2-phenylethyl)-isobutyramide,which effectively inhibited the QS phenotypes in several Gram-negative biosensors[34].This active compound represents a promising chemotype for inhibitors of QS phenotypes.These co-cultivation assays were applied to screen a host of organisms to inhibit QS phenotypes.
2.2.Construction of engineering strains for QSI screening
However,these colorimetric assays with natural strains cannot meet the requirement of large-scale QSI screening.To promote large-scale screenings,the QSI selector system,including a plasmid that involves a construct composed of a QS homolog gene such as LasI,LasB,and RhlA,as well as a phenotype or function regulated by a QS-mediated promoter,has been extensively developed[35-46].
2.2.1.QSIs-LasI biosensor
The QSIs-LasI biosensor emerged as an innovative strategy for QSI discovery,and diverse QSIs were identified by virtue of this biosensor.Using the P.aeruginosa QSIs-LasI biosensor combined with analysis of QS-controlled phenotypes and HPLC,equisetin in secondary metabolites from the marine fungus Fusarium sp.Z10 and four new alpha-pyrones from the secondary metabolites of Streptomyces sp.were discovered as the bioactive anti-QS components[37,38].In one instance,Wang et al.designed an efficient screening system involving a QSIs-LasI selector based on the PlasI-sacB reporter.Plasmid pMHLASI harboring PlasI-sacB and Plac-LasR transcriptional fusion was constructed and transformed into a P.aeruginosa LasI-RhlI double mutant PAO-MW1 to generate the selector QSIs-LasI.They tested the crude extracts from 65 marine fungi,and then an isolate,Penicillium Atramentosum QJ012,was selected to exhibit an inhibitory effect on the QS system.The thinlayer chromatography assay of the fungal extracts for bioautographic identification by QSIs-LasI demonstrated that the fungus produced several valuable compounds with anti-QS effects[39].
2.2.2.LasB-gfp and RhlA-gfp biosensor
The LasB-gfp and RhlA-gfp monitor strains contain an LasB promoter regulated by LasR,an RhlA promoter regulated by RhlR,and genes controlling an unstable green fluorescent protein.Using this biosensor,active compounds such as zeaxanthin and evernic acid were discovered to downregulate QS-related genes and inhibit biofilm formation in P.aeruginosa[40,41].Rasmussen et al.[42]constructed three novel QSI selectors for QSI screening,including QSIs1 phlA encoding the toxic gene product,the expression of which was controlled byLuxR;QSIs2,the LasR-and RhlR-regulating LasB promoter controlled expression of the sacB gene,which led to cell death in the presence of sucrose;and the QSIs3 system,which contained the npt and gfp genes,conferring kanamycin resistance and green fluorescence,respectively.By virtue of these QSIs systems,two potential QSIs,garlic extract and 4-nitro-pyridine-N-oxide,were identified.In addition to in vitro phenotypic assays,GeneChip-based transcriptome analysis and a Caenorhabditis elegans pathogenesis model were also used to evaluate the specificity of QS-regulated virulence genes and virulence in vivo.
2.2.3.AHL-biosensor and others
The application of biosensors for the detection of the inhibitory activity in AHL-mediated QS has been instrumental in the rapid and streamlined screening of the QSI in a broad spectrum of microbes,especially Gram-negative bacteria.The realization that structurally different AHLs exist,possessing diversity in the acyl side chain and having different activation profiles,has prompted the construction of the corresponding biosensors.These reporters could be utilized in further studies to characterize the AHL signals in QS in a broad spectrum of organisms.
Using AHL-biosensors to screen out from the QSI chemical library and AHL-analogs as well as secondary metabolites produced by microbes,marine Streptomyces and Flustra foliacea,a large number of novel QSIs including N,N′-alkylated imidazoliumderivatives,and some secondary metabolites were identified as potential QSIs[43-45].
In another study,coumarin was identified as a potential QSI using a novel screening system composed of three biosensors(Serratia marcescens SP15,C.violaceum DSM30191,and Agrobacterium tumefaciens NTL4 containing a plasmid pZLR4 carrying a traG:lacZ reporter fusion)to detect short,medium,and long AHLs,respectively.Subsequently,coumarin was demonstrated to significantly suppress biofilm formation and protease activity in other microbes and inhibit bioluminescence in Aliivibrio fischeri[46].All cases mentioned above are shown in Table 2[29-46].
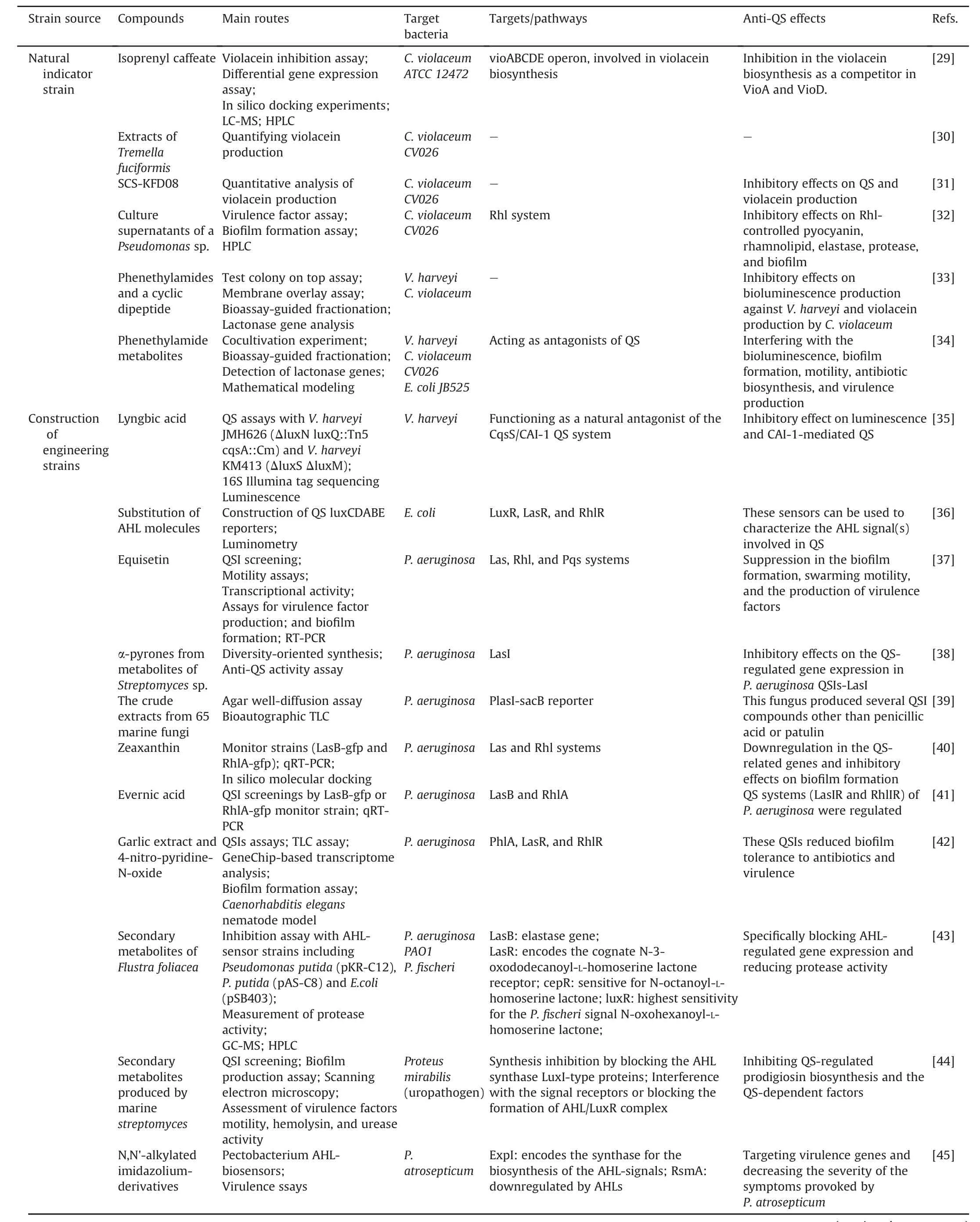
Table 2Activity-based screening with bioassays.
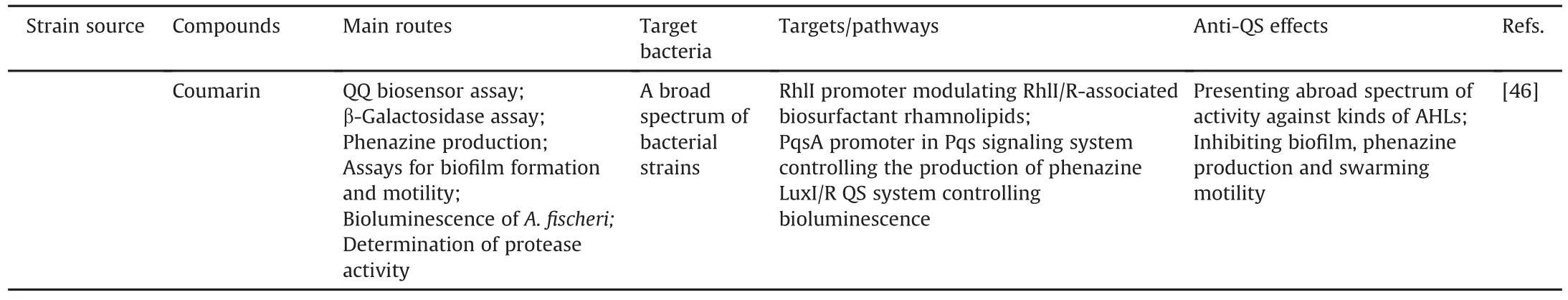
Table 2(continued)
2.3.Other activity-based bioassays
Other series of activity-based assays use known QS-mediated mechanisms for functional differentiation.The QS-regulated functions tested in these bioassays involve motility,pyocyanin or pyoverdin production,and biofilm formation in P.aeruginosa.In this regard,similar steps are applied in the preliminary stage,including diffusing potential QSIs into agar;measuring the area of pigment inhibition,which is visualized by the formation of a colorless but visible halo around the well;and the method of microbulion dilution,which quantitatively detects the level of pigment by measuring the optical density using a spectrophotometer[47,48].Numerous studies have described the discovery of QSIs based on biofilm formation.Commonly used qualitative methods to evaluate the characteristics of the biofilm in these cases involve Congo red agar,the microtiter plate assay,or the crystal violet assay.
Another novel approach used for QSI discovery is proteomic analysis,which is a two-step procedure involving an initial quantitative 2D-difference gel electrophoresis and subsequent protein identification in each spot by mass spectrometry.This method has been proven to be beneficial for the examination of the impact of various factors on a range of proteomic and metabolomic changes mediated by QS in bacteria[49].
Transcriptome analysis provides deep insights into QS modulation in response to anti-QS compounds[50,51].Comparing realtime polymerase chain reaction used in small-scale genes for QSI screening in early studies,this transcriptome analysis based on high-throughput sequencing methods achieved more quantitative analysis of comprehensive transcription profiles.The analysis included 1)total RNA extraction of bacterial samples,2)RNA reverse transcription into a complementary DNA library for transcriptome sequencing,and 3)comparative analysis of differentially expressed genes by various genetic techniques.The subsequent differential expression analysis by bioinformatics approaches is beneficial for revealing and finally characterizing global gene expression patterns in response to anti-QS compounds.
In vivo models are widely used to evaluate the efficiency of potential QSIs for antivirulence activity.The Caenorhabditis elegans model achieves in vivo high-throughput screening(HTS)and provides a better understanding of QS modulation and virulence regulation[52].Additionally,mammalian models with general immune systems are also commonly employed to evaluate the anti-QS activities of potential QSIs on bacterial infections[53].
3.Chemical approaches to interrogating QS pathways for QSI discovery
3.1.Solid-phase synthesis for the identification of small molecule QS modulators by interaction with their target proteins/receptors
Each compound yielded by traditional synthesis strategies requires individual purification,identification,and bio-activity evaluation,rendering the acquisition process cumbersome and timeconsuming.Unlike traditional synthesis methods,the update of combinatorial chemistry involving solid-phase synthesis significantly accelerates chemical synthesis and screening.Solid-phase synthesis,in which the reactant molecule is chemically bound to an insoluble material while reagents are added in the solution phase,is extensively applied in the high-throughput synthesis of biological molecules such as peptides,nucleic acids,oligosaccharides,and QSIs[54].
Geske et al.[55-57]constructed a library of AHL-analogs using a solid-phase synthesis technique.They adopted a novel synthetic strategy containing a series of microwave-assisted reactions followed by a CNBr cyclative-cleavage procedure on the polystyrene beads.Furthermore,through systematic screening of these analogs,numerous antagonists were identified to competitively bind to the LuxR-type protein.These compounds were found to attenuate QS-mediated phenotypes in bacterial culture as well as in eukaryotic host infections.
Novel solid-phase synthesis techniques such as macroarrays and microarrays were studied for the discovery of QSIs[58-64].In small molecule macroarrays,libraries of varying sizes were formed by the spatial synthesis of small molecules on planar polymeric supports.This platform,with advantageous properties such as economics,stability,and compatibility,was proven to be a practical approach to the large-scale synthesis of small molecules including QSIs[65,66].
Not limited to macroarrays,microarray techniques were also investigated and utilized for the combinatorial synthesis of QSIs[67].Sharing the analogous underlying principle with macroarray synthesis,the size and diversity of microarrays are apparently smaller and simpler[68,69].Moreover,macroarrays and microarrays simultaneously provide platforms not limited to chemical synthesis,but many on-support biological tests are involved,further facilitating the discovery of bioactive anti-QS molecules.
3.2.Affinity chromatography approaches for identifying QS receptors based on ligand-receptor interactions
Not limited to synthetic AI analogs as QS modulators,ligand-receptor interactions in QS signaling have also been studied for target identification of QS receptors.Affinity chromatography based on ligand-receptor interactions,with its exquisite specificity,has been proven to be a valuable tool for the purification and identification of biomolecules.In affinity chromatography,one molecule is immobilized to a support,whereas its binding counterpart is soaked in the mobile phase.Under alternate conditions,molecular recognition between the two molecules allows the counterpart to be isolated from the eluent and separated from the support.This highly selective separation approach is of significant value in the identification,isolation,purification,pretreatment,or analysis of many biologically relevant molecules.
To identify AHL-binding receptors,Spandl et al.[70]synthesized biotin-labeled N-3-oxo-hexanoyl L-homoserine lactone(OHHL).The biotin tag,with powerful affinity and recognition to avidin,permits the ligand OHHL to be readily immobilized to an avidinfunctionalized support.Biological tests and 2D-difference gel electrophoresis have also been pursued to investigate the target for OHHL.In a similar work,affinity resins were utilized to identify putative receptors for AHLs in eukaryotic cells.Two piperazinemodified N-(3-oxo-dodecanoyl)-L-homoserine lactone(OdDHL)derivatives were synthesized and linked to agarose resin[71].Subsequently,two proteins were successfully obtained by affinity assay by virtue of these OdDHL-modified resins from mammalian cell lysates.Such resins in affinity-based methods could be ultimately valuable for studying novel AHL-binding proteins.
3.3.Antibodies for quenching QS signaling
Another approach that has gained considerable attention for interrupting QS signaling is to apply antibodies to quench QS by competing with or interrupting QS signal molecules[72].The AIs or derivatives thereof are delivered into an animal to activate monoclonal antibody formation.The yielded antibodies,which are substantially specific for the AI molecules,could catalytically inactivate or effectively sequester AIs by acting as the QS quenchers,resulting in the QS signaling interruption[73].Usually,generating antibodies such as QQ needs a molecular template(or hapten)to immunize.Small QS molecules such as AHLs,with hydrolytic instability and nonproteinaceous nature,cannot complete antibody-based immune reactions by themselves because they are unlikely to be appropriately presented by the mammalian immune system.The antibody-based strategy has promising applications in future vaccine research and development.
In recent years,numerous studies have reported the design and evaluation of QQ antibodies to sequester AIs in vitro and in vivo[72,74].Using an immunotherapeutic strategy for QQ,Janda and coworkers[75]pioneered eliciting immune responses against the synthetic 3-oxo-AHL analog RS2 in mice,and isolated an anti-AHL antibody(RS2-1G9).The RS2-1G9 efficiently inhibited QS phenotypes and QS pathways in P.aeruginosa by sequestering 3-oxo-C12-HSL.Furthermore,they utilized this approach to raise antibodies against a similar analog of AIP-IV.In this case,the native thioester in the AIP macrocycle was more hydrolytically stable using an ester bond.The antibody obtained,AP4-24H11,was shown to be an active QSI in S.aureus cultures[76].
Additionally,antibody-based QQ also involved other strategies,such as raising catalytic antibodies for degrading and thus inactivating AIs.Marin et al.[77]utilized this approach to screen and evaluate catalytic antibodies for lactonase activity.An antibody(XYD-11G2)capable of hydrolyzing OdDHL from an antibody library was identified and demonstrated to suppress QS in P.aeruginosa cultures.
3.4.Biotic polymers/polymeric materials
Materials-based approaches have also gained considerable attention for QS modulation.The first strategy is the sequestration of the AI signal by non-native polymeric materials.By computational modeling and prior affinity studies,Piletska et al.[78]selected a series of monomers with binding capacity with an AHL signal named OHHL produced in V.fischeri.Polymers based on one monomer,named itaconic acid(IA),showed higher affinity to OHHL and thus suppressed QS of V.fischeri.Subsequently,they generated a polymer production composed of a template molecule and polymers synthesized by utilizing molecularly imprinted polymers[79].The formed IA-based polymer production served as high-specificity binding pockets to sequester QS signaling and biofilm formation in P.aeruginosa.In another study,2-hydroxypropyl-βcyclodextrin(HP-β-CD)was reported to downregulate the levels of QS-controlled genes in Serratia marcescens and P.aeruginosa.Then,cross-linked HP-β-CD was utilized to form sheets of 2-hydroxypropylcellulose to form an AHL-sequestering material.Subsequently,these HP-β-CD-linked sheets were reported to suppress the levels of QS-dependent genes when co-cultured with the two abovementioned strains.
Another strategy is to utilize polymer materials for the spatial accumulation of QS modulators and thus to prevent the release of QS modulators into the surroundings.Poly lactide-co-glycolide is employed as a material to regulate the release and delivery of AHLs.Both the AHL concentration and the time of AHL release could be readily controlled without affecting its biological activity[80].These studies indicated a novel material-based strategy for modulating QS signaling that can be applied in a series of contexts(e.g.,biofilm prevention and coatings on the surface of clinical implants).All those mentioned above are shown in Table 3[56,57,64,66,69,76-78,80].
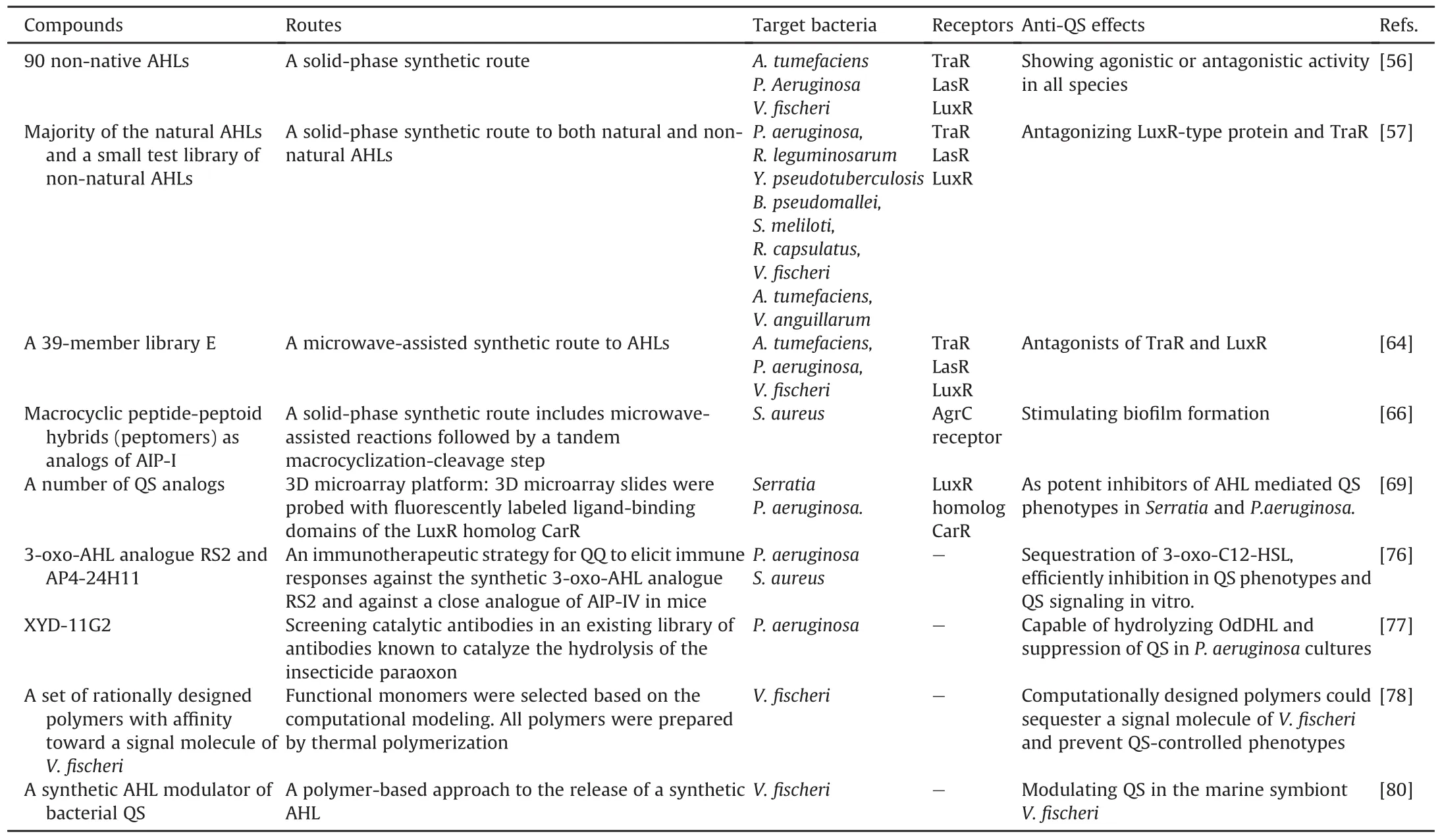
Table 3QSI discovery by chemical techniques.
4.Virtual screening for QSI screening
As described above,limited QSIs have been identified using traditional methods from natural resources.Computer-assisted screenings,especially structure-based virtual screening(SB-VS),have been developed as an efficient paradigm for lead discovery that fits in well alongside HTS to promote large-scale screenings,which can complement traditional methods.
The SB-VS approach,which is used extensively in the pharmaceutical industry,has also been applied for novel QSI discovery that antagonized AHL sensing in recent years:1)against Las,Rhl,and Pqs systems:the screening of natural compounds and recognized drugs/compounds by in-silico docking analysis and SB-VS in P.aeruginosa[81,82];2)against the LasR and RhlR receptor proteins:the identification of five active compounds using an SB-VS of 1920 natural compounds[83];3)against LasR receptors:two compounds discovered by virtual screening and pharmacophore-and structurebased approaches and six novel potential QSIs screened with parameters including Lipinski's rule,and toxicity as well as absorption,distribution,metabolism,excretion(ADME)from the ZINC database[84,85];4)with the crystal structure of LuxP:the screening of cinnamaldehyde derivatives[86],boronic acid derivatives[87],and traditional Chinese medicine(TCM)databases[88]by SB-VS;5)other targets/systems:the identification of baicalein in 51 bioactive components from TCMs against TraR protein[89],the discovery of eight lead-like molecules from the ZINC database by combined analysis including virtual screening,molecular docking as well as ADME and toxicity studies against FabI which plays the important role of enoyl-acyl carrier protein reductase related to the synthesis of 3-oxo-C12 HSL[90],the screening of most plant compounds and all NSAIDs to find the Z-phytol and lonazolac molecules which can suppress the SdiA as a homolog of LuxR in the QS pathway[91].
To sum up,SB-VS approaches represent an economical and rapid alternative to the traditional methods and HTS approaches.Screening strategies for SB-VS can be limited to commercially available compound libraries to avoid time-consuming and expensive chemical syntheses and utilized to dock compounds from known drugs/natural product libraries to circumvent problems in complicated testing stages,owing to toxicity or poor ADME properties,which would substantially reduce the costs,compared to conventional resource-consuming HTS processes.However,SB-VS approaches,relying on the prediction by the docking software,have one major problem of false positives and false negatives.As more novel and advanced algorithms are developed,false hits may be minimized.Moreover,experimental validation is required to combine SB-VS for in vitro inhibition efficacy.
The SB-VS approaches promote the identification of targeted QSIs,and future directions include extending SB-VS to more novel targets and performing combined tests with quantitative structureactivity relationship studies to promote rational drug design.All cases mentioned above are shown in Table 4[81-91].
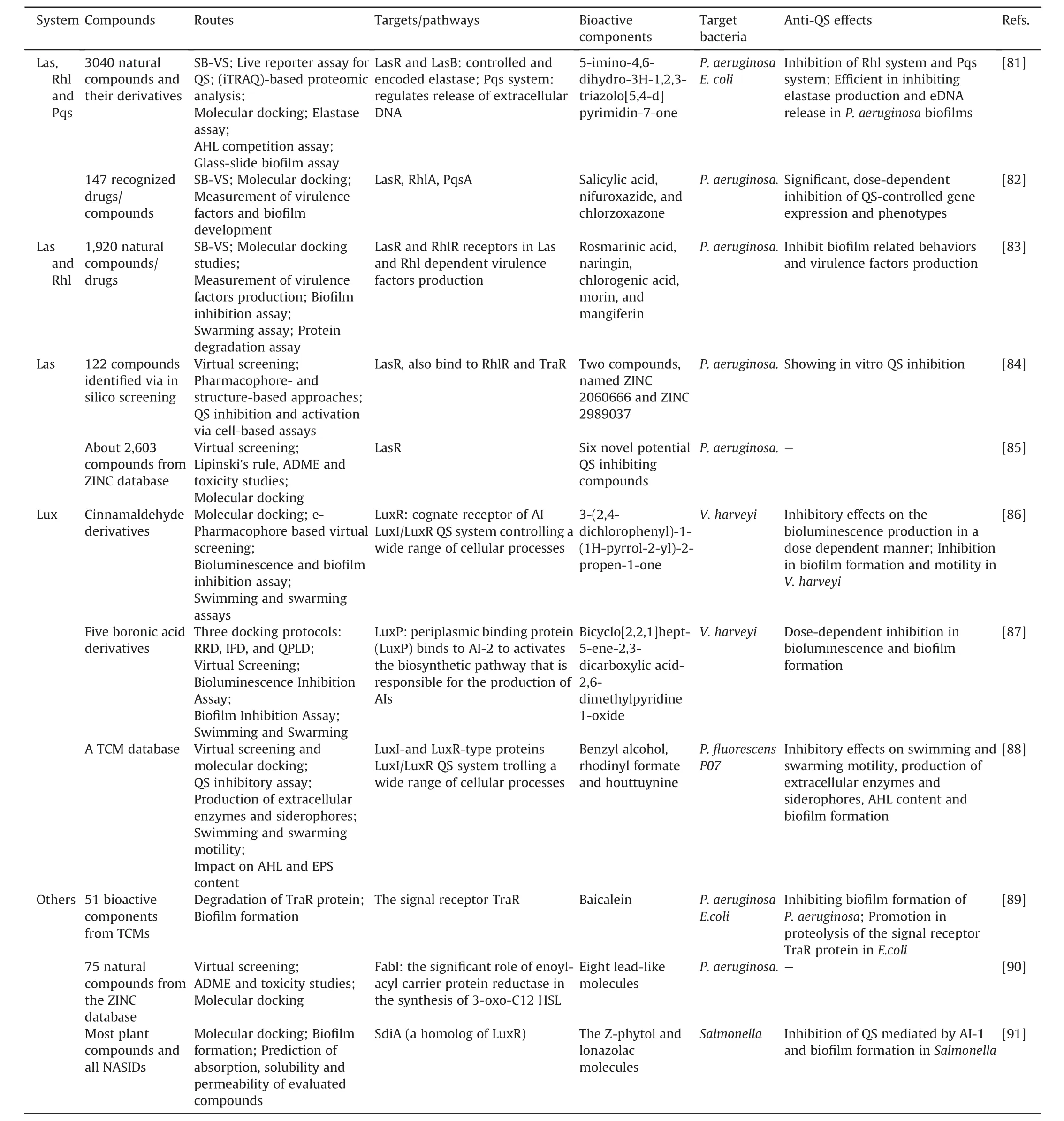
Table 4Virtual screening for QSI screening.
5.Other potential tools for interpreting QS signaling
5.1.Three-dimensional(3D)printing
A microscopic 3D printing strategy was established to organize multiple populations of bacteria within essentially any 3D geometry.Using this approach,a distinct core-shell arrangement,which was composed of a single species at vastly differing densities as well as polymicrobial communities including motile and nonmotile bacteria,was found in microbial populations.For example,Connell et al.[92]also demonstrated that a picoliter-volume aggregate of S.aureus had the ability to produce substantial resistance to β-lactam antibiotics by nesting a unique core-shell arrangement enclosed with a shell composed of P.aeruginosa.
5.2.Immobilized hybrids
Using an operationally simple dip-and-rinse procedure,Gomes et al.[93]synthesized natural product hybrids featuring an AHL inducer as well as a nitrodopamine and subsequently immobilized the hybrids onto biocompatible TiO2surfaces.The formed immobilized hybrids were demonstrated as effective QS activators by gradually releasing them from the TiO2 surface.This approach is a straightforward and useful strategy for interrupting the QS signaling pathway by the preparation of coated surfaces in medical devices in a wide variety of bacterial species.
5.3.Biological nanofactories
Hebert et al.[94]constructed a biological nanofactory to regulate the behaviors of bacteria on the surface of human epithelial cells.The biological nanofactory construct activated the surface to produce the bacterial QS signaling molecule,AI-2.Biological nanofactories include several functional subassemblies:the cell targeting module with the targeting element CD26 antibody,the fabrication,and the synthesis modules.The biosynthesis component implemented is the fusion protein His6-protein G-LuxS-Pfs-Tyr5(HGLPT).The HGLPT consisted of both synthesis and fabrication domains,which facilitated flexible self-assembly of the functional units. The HGLPT utilized the substrate S-adenosylhomocysteine to synthesize AI-2 in a two-step process using enzymes such as Pfs and LuxS.This tool would be beneficial for modulating signaling activities associated with the microbiome present in the human gastrointestinal tract and other environments.
6.Challenges in QSI discovery and some criteria for QSIs
To date,many recently authorized patents focused on the discovery of new bioactive agents,analogs,and approaches interfering with QS signaling highlight the application of QS inhibition in the fields of plant pathogens,aquaculture,antifouling,and medical devices(Table 5)[95-111].However,very few QSI compounds have reached human clinical trials.Until now,QSI therapeutic studies for clinical QS-induced bacterial infections are still undergoing preclinical phases and have not been extensively developed.In total,there are only a few clinical trials on azithromycin(a QSI for preventing P.aeruginosa ventilator-associated pneumonia and treatment of cystic fibrosis)(Table 6)[112].Some beneficial QSI effects,such as the ability to prevent biofilm formation,bacterial virulence,and innovative medical devices equipped with QSI molecules have attracted the most attention.Various inhibitors/agents capable of QS-controlled biofilm formation are ongoing clinical evaluations(Table 7)[112]and have been published with outcomes[113-118].Moreover,QSIs were primarily designed for functionalizing catheters.The increasing range of QSIs fuel efforts to develop innovative medical devices and diversify their applications.

Table 5Recently authorized patents on the discovery of new bioactive agents,analogs and approaches interfering with QS signaling.
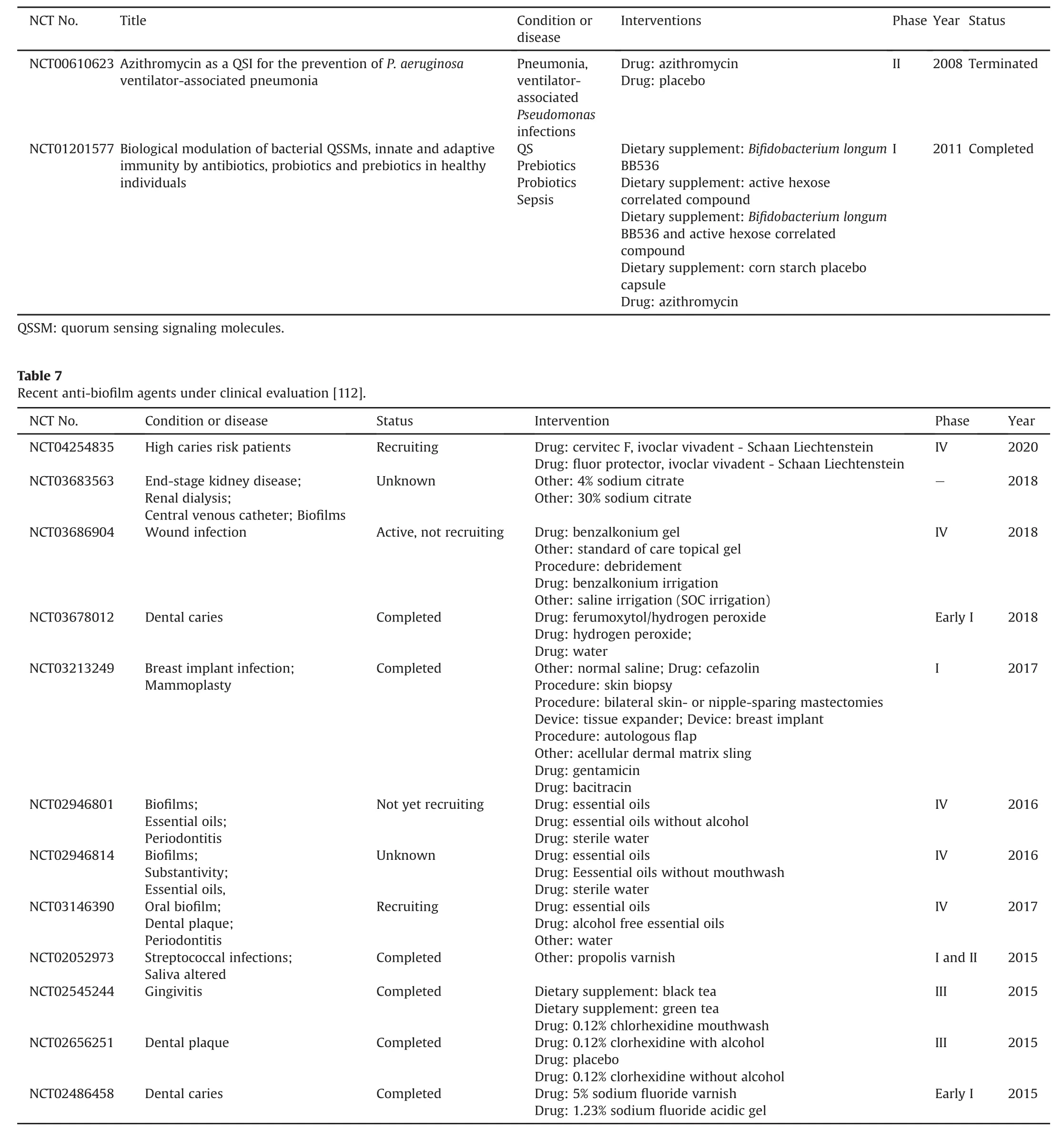
Table 6Quorum-sensing inhibitors in clinical evaluation[112].
Unlike antibiotics,QSIs act by interfering with QS-mediated gene expression controlling virulence traits that are not necessary for bacterial growth,which probably produces a limited selective pressure to develop bacterial resistance.However,some studies have pointed out the risk of inducing selective pressure and developing resistance in QS disruption conditions by anti-QS agents[119,120].However,the literature manifestly indicates that resistance to QS inhibition may emerge,but likely at a much lower level than that which conventional antibiotics could induce with their massive use.To date,there have been few reports regarding the potential QQ-related resistance mechanisms that various bacteria could induce to support this possibility,but it cannot be completely ignored.
At present,only a few QSIs were finely clarified for the anti-QS mechanisms supporting the related anti-QS activity at a subcellular level and functionally evaluated regarding their biological role in vivo.In this regard,QSIs may not be as active as antibiotics,and are in realistic conditions,which becomes a major bottleneck for their potential for antibacterial treatments to reach clinical trials.In some cases,QSIs have been described as having potential medicinal effects on bacterial communities in the human body[121].The impacts on the hose microbiome were quite likely to influence human health and induce metabolic disturbance[122].
Overall,to prevent the negative influence on hosts and minimize the development of QSI resistance,some criteria for QSIs should be considered:1)low molecular weight;2)high stability and specificity;3)no side effects on hosts and no interference with the host microbiome;and 4)unlikely to develop drug resistance.
7.Conclusions
Over the past two decades,numerous studies have reported that QS circuitry and potential QSIs have significant impact as antiinfective agents against a host of bacteria.The discovery of QSIs and their initial success represent a promising target for the design of novel anti-infective drugs.The discovery of QSIs is appealing in large part to expand the anti-infective therapeutic arsenal to complement classical antibiotics and antimicrobial agents.With regard to QSI discovery,some problems that need to be solved,including the QSI molecular target identification,the molecular targeting delivery,the QSI cytotoxicity evaluation at the organism,cellular,and molecular levels.These queries on the above points are certainly promising trails for further efforts to discover QSIs and diversify their applications.
CRediT author statement
Lan Lu:Project administration,Funding acquisition,Writing-Original draft preparation,Supervision;Mingxing Li:Investigation,Writing-Reviewing and Editing;Guojuan Yi and Li Liao:Conceptualization,Methodology,Software;Qiang Cheng,Jie Zhu and Bin Zhang:Conceptualization,Investigation;Yingying Wang,Yong Chen and Ming Zeng:Visualization.
Declaration of competing interest
The authors declare that there are no conflicts of interest.
Acknowledgments
This study was funded by the National Natural Science Foundation of China(Grant No.:81803812).
杂志排行

Journal of Pharmaceutical Analysis的其它文章
- Capillary electrophoresis methods for impurity profiling of drugs:A review of the past decade
- Antiviral,immunomodulatory,and anticoagulant effects of quercetin and its derivatives:Potential role in prevention and management of COVID-19
- Anti-inflammatory and analgesic properties of Moroccan medicinal plants:Phytochemistry,in vitro and in vivo investigations,mechanism insights,clinical evidences and perspectives
- Assessment of the binding interactions of SARS-CoV-2 spike glycoprotein variants
- Identifying the molecular basis of Jinhong tablets against chronic superficial gastritis via chemical profile identification and symptomguided network pharmacology analysis
- MTBSTFA derivatization-LC-MS/MS approach for the quantitative analysis of endogenous nucleotides in human colorectal carcinoma cells