中国陆地生态系统植硅体碳汇潜力估算研究进展
2022-04-01盛茂银王霖娇
殷 婕,盛茂银*,王霖娇
(1.贵州师范大学 喀斯特研究院,贵州 贵阳 550001;2.国家喀斯特石漠化治理工程技术研究中心,贵州 贵阳 550001;3.贵州省喀斯特石漠化防治与衍生产业工程实验室,贵州 贵阳 550001)
0 引言
单硅酸作为有效硅的一部分,在被高等植物吸收后,不断沉淀在植物细胞中而形成了非晶质二氧化硅矿物颗粒[1-2],称为植硅体(Phytolith)。植硅体有着抗氧化,耐高温等特性,并且其本身理化性质稳定[3-4]。因此,在植物生长,分解过程中植硅体相比其他矿物颗粒更不易分解,而得以稳定保存在土壤或者沉积物中。
在植硅体形成过程中,会不断吸收生态系统中的有机碳,一部分有机碳被吸收后封存于植硅体内而形成植硅体闭蓄有机碳(Phytolith-occluded-carbon,PhytOC),简称植硅体碳[5-6]。这一部分碳由于植硅体不易分解,可以长期,安全的存在于植物中,待植物死亡分解后,成为不参与大气碳循环而储存在土壤中稳定的有机碳库[7],得以数千年至上万年储存于土壤中。所以植硅体碳作为陆地稳定碳汇机制之一,在全球气候变化中发挥着重要的作用[1]。
近几十年来,我国与植硅体碳汇潜力相关的研究范围逐步扩展到农田、森林、草原和湿地等多个生态系统[8]。关于土壤碳库的研究[9]、植硅体在土壤中迁移[10]以及凋落物对植硅体迁移的研究[11-12]都有涉及,研究发现植硅体碳含量受多方面因素影响,而表现出不同差异。胡宁等[13]的研究发现凋落物中的植硅体碳进入土壤后含量虽表现出降低的规律,但其分解对土壤有机碳的提高有促进作用。由此可看出不同植被,枯枝落叶甚至土壤中植硅体碳含量都不尽相同。为了准确估算陆地生态系统植硅体碳汇潜力,为全球碳循环研究提供参考,本文对中国陆地生态系统植被地上/地下器官、植被凋落物、土壤等环节植硅体碳汇潜力估算的研究成果进行综述,总结中国陆地生态系统上述不同环节植硅体碳汇潜力差异,揭示不同环节碳封存潜力特征,为进一步研究指明方向。
1 植被地上器官植硅体碳汇潜力
植物作为植硅体产生的载体,不同植物产生的植硅体含量不一样。由于受到植硅体本身所具有的特性以及外部环境的影响,植硅体碳含量及封存潜力存在差异[5,14]。Zhang等[15]研究中提到蕨类植物的植硅体含量较高,而被子植物中单子叶禾本科的植硅体含量高达10%~15%[16],朱浩宇等[17]在研究中也提到,单子叶植物的合成植硅体能力要高于双子叶植物。因此,关于植硅体方面的研究多集中在植硅体合成能力较高的植物,即禾本科植物,如竹子、水稻、甘蔗[18-20]等。植物中的植硅体在细胞内不断沉淀而赋存于各器官中。有研究显示,大多数植物的叶片植硅体含量最高[3],因此,地上部分的植硅体碳汇潜力研究多集中于叶片部分。
应雨骐等[21]以亚热带森林生态系统中的典型树种为背景,通过计算不同树种中叶片的碳封存速率,对植物地上部分植硅体碳汇潜力进行估算得出:枫香一年的植硅体碳汇潜力为0.48×106t CO2、而苦槠仅为0.01×106t CO2;针叶林中,杉木每年可封存0.06×106t CO2、马尾松每年可封存0.13×106t CO2。Song等[22]对我国典型森林生态系统的地上部分植硅体碳汇潜力进行估算,寒温带和温带针叶林每年的植硅体碳汇潜力为0.18×106t CO2、针叶和阔叶混交林为0.04×106t CO2、亚热带常绿与落叶阔叶林为0.27×106t CO2、亚热带常绿阔叶林为0.28×106t CO2。以上两人研究结果均发现,在不同土地覆被类型下,针叶林的植硅体碳汇能力相对较低,这主要是因为针叶林植硅体含量较低原因,这与王丹等[23]的研究结果相似。从以上结果还发现,在气候较为湿润的地区植硅体碳汇潜力均较高,可见植硅体碳汇潜力可能与水热条件和植被覆盖类型有关。
竹子本身作为富硅禾本科植物,其植硅体固碳能力被众多学者认可[24-25]。Song等[22]研究发现中国森林生态系统每年植硅体碳汇潜力为1.70×106t CO2,而竹林的植硅体碳汇潜力就占全国30%,因此未来中国若采用竹子为植树造林工程对象,那每年仅地上部分的植硅体碳汇潜力就可达5.60×106t CO2。在竹子各个器官的碳汇潜力研究中,尹帅等[26]分析了绿竹和麻竹地上部分不同器官下的植硅体碳汇潜力差异,发现这两类竹子的枝和叶相比于杆碳汇潜力更高,这和冯晟斐等[27]、张金林等[28]以及孙凯等[29]的研究结果一致,最后尹帅等[26]还估算出我国绿竹林地上部分植硅体每年的碳汇潜力在0.19×104t CO2。


表1 中国草地生态系统植硅体碳汇潜力
高寒草甸和荒漠草原的碳封存潜力相差较大的原因主要是植硅体的碳汇潜力与地表净初级生产力之间呈现极显著的正相关[31,34],而荒漠草原的地表净初级生产力低于较湿润的草原类型,因此导致草甸的植硅体碳汇潜力较高。
农作物中,Li等[19]对水稻地上部分的植硅体碳含量进行计算后,估测得出中国水稻每年的植硅体碳汇潜力为3.88×106t CO2。Song等[35]对其他农作物研究发现玉米的碳汇潜力显著高于甘蔗、棉花等经济作物,并通过研究农作物各器官植硅体含量,发现茎、叶鞘、叶的含量都较多(表2),这与Li等[19]的研究结果相似,植硅体含量高是作物有较高的植硅体碳汇潜力的基础。左昕昕等[36]通过计算旱作植物粟和黍的植硅体碳封存速率和10年来的平均粮食产量,估算出我国旱作农业每年的植硅体碳汇潜力为2.37×106t CO2。

表2 中国农田生态系统植硅体碳汇潜力
由以上研究对比发现,植物地上部分植硅体碳汇潜力在整个生态系统的长期固碳减排中发挥着重要作用,其中森林生态系统和草原生态系统的碳汇潜力都不可小觑。目前植物不同器官内的植硅体及植硅体碳含量已有相关研究,但仍主要集中在富硅植物,因此还需加强其他植物各器官含量计算,以此加强对整个生态系统碳汇能力的认识。
2 植被地下部分植硅体碳汇潜力
现有植硅体碳汇潜力研究中,大多数学者还是集中于植物的地上部分,这可能是因为植物地下部分比较难获得。有研究学者指出植物中(水稻)根系的植硅体碳汇量相比茎叶较低[19,37]。但即使植物地下部分植硅体碳汇潜力低于地上部分,其地下部分的植硅碳汇量也是植硅体碳汇中不可或缺的一部分。
通过植硅体碳汇潜力的计算公式可知,碳汇潜力和净初级生产力息息相关[19]。有学者研究发现,在温带干旱和半干旱地区,多数草地的地下净初级生产力是地表净初级生产力的19倍左右[38],且地上部分植硅体封存的有机碳含量只是生态系统中的小部分,仅占2%~4%[30]。Qi等[30]通过估算得出,典型草原每年的地下部分植硅体碳汇潜力在0.31~0.60×106t CO2,确实比地上部分碳汇量高。这主要是因为草地生态系统的地下部分多为多年生,而地上为一年生,因此其封存碳的累积时间不同,而上文中提到水稻根系植硅体碳汇潜力比茎叶低,这可能与水稻的根系和茎叶都为一年生有关。
通过植硅体碳储量计算公式可知,植硅体碳储量=植硅体碳含量×生物量[39],并且生物量较净初级生产力更易获得,因此大多数学者在研究竹类地下部分时主要还是估算其植硅体碳储量。陈晨[39]研究6种散生竹发现其地下部分植硅体碳储量占整株植物的10%,且该6种散生竹地下部分植硅体碳储量为0.09×106t C,而丛生竹的地下部分植硅体碳储量为0.89×105t C。Huang等[40]通过研究发现雷竹地下部分植硅体碳储量占雷竹总植物部分的49.6%,本文再依据陈晨[39]论文中全国雷竹分布面积数据,对其雷竹地下部分植硅体碳储量进行计算,得出其植硅体碳储量为0.21×104t C。通过植硅体碳汇潜力计算公式[41],植硅体碳储量×3.67(C 转换为 CO2的系数),计算得出陈晨研究的6种散生竹地下部分每年植硅体碳汇潜力为0.33×106t CO2,丛生竹为3.26×105t CO2,Huang研究的雷竹地下部分植硅体碳汇潜力为0.77×104t CO2。但这主要是本文作者通过转换系数计算得出,可能与实际研究结果有差异。
通过以上研究对比发现,植物地下部分的植硅体碳汇潜力相对于地上部分较少,但可能是地下部分的研究相对欠缺。这些研究主要集中于植物地上部分,缺少对植物地下部分植硅体碳汇潜力研究,这影响了对植物各部分植硅体碳汇潜力差异的对比,也制约了对整个生态系统植硅体碳汇潜力的精确评估。因此需进一步加强植物各个环节植硅体碳汇变化的相关研究,对比植物各个器官的植硅体碳汇潜力差异。
3 植被凋落物植硅体碳汇潜力
土壤中的植硅体碳在积累过程中主要依靠凋落物的大量输入,凋落物是指植物地上部分生产并归还到地表的所有有机质的总称[42],主要包括凋落叶、繁殖体和树枝等组分。凋落物不断分解后,其内的植硅体碳被逐渐释放到土壤中作为地球碳库不可或缺的一部分,从而间接提高了土壤中植硅体碳的含量[43]。因此尽管凋落物中植硅体碳含量在生态系统中所占比例小,但它作为腐屑食物链的重要物质基础,是物质循环和能量流动的重要环节[44]。
一般认为,凋落物各器官中,叶片所占比重最大,枝条次之[45-46],因此凋落物的植硅体碳汇研究主要集中在叶片方面。Li等[18]对35个种属竹子的叶片凋落物进行植硅体碳汇潜力估算,发现中国竹子凋落物每年的植硅体碳汇潜力0.29×106t CO2。Huang等[40]研究发现通过鲜叶输入到土壤中的植硅体碳含量远小于通过凋落物输入的量,而雷竹凋落物植硅体碳汇潜力为41.45 kg/hm2,本文通过陈晨[39]论文中的全国雷竹分布面积数据,计算得出雷竹凋落物每年的植硅体碳汇潜力为0.67×104t CO2。胡晓薇等[47]估算了毛竹林每公顷凋落物的植硅体碳汇潜力,本文再通过陈晨[39]论文中的面积计算得出毛竹林凋落物每年的植硅体碳汇潜力为0.05×106t CO2,这与雷竹的碳汇潜力相差较大,这可能是毛竹林在我国分布面积较大的原因。此外,应雨骐等[21]以中国亚热带的杉木林、阔叶林、针阔混交林面积,估算得出中国亚热带杉木林、阔叶林、针阔混交林每年的地表凋落物植硅体碳汇潜力分别为0.12×106t CO2、0.57×106t CO2、0.28×105t CO2。而在农作物方面,Li等[19]通过对水稻的碳汇潜力进行估算后发现,若收割稻子后将剩余部分全部用做堆肥,那么中国在1950—2010年60年间通过水稻封存了2.37×108t CO2。
对比以上研究发现植硅体的碳汇潜力从植物到凋落物,主要呈下降趋势,这与何珊琼等[48]的研究结果相似,但凋落物对植硅体从植物向土壤转化,并增加植硅体碳汇潜力的作用还是积极的。因此在之后的研究中,不能忽视凋落物的碳汇潜力,并且要更加注重植被-凋落物-土壤这一过程中植硅体碳汇潜力的变化趋势,以便为大尺度的生态系统碳汇变化研究提供有力依据。
4 土壤植硅体碳汇潜力
土壤作为一个巨大且具有挥发性的潜在碳库,不但可以缓冲大气中二氧化碳浓度的增加,也可通过土壤内碳库微小变化而引起大气碳浓度的显著变化[49],因此分析计算土壤中的植硅体碳汇潜力就显得尤为重要。而植硅体碳主要被封存于植硅体中,植硅体通过植被-凋落物-土壤这一环节进入土壤后,通过枯枝落叶返回到土壤中的植硅体有92.5%被植物再吸收,7.5%进入土壤库[20]。尽管进入土壤部分的植硅体占比很小,但由于植硅体不易被分解,其在土壤中不断积累,封存于植硅体内的有机碳也不断累积,使土壤中的植硅体碳在经过2 000多年的分解后,还能占土壤总有机碳的82%[50-51]。由此可见,土壤植硅体碳在稳定全球气候,减缓温室效应方面发挥着重要作用。
4.1 不同土地覆被土壤植硅体碳汇潜力
地表植物群落在土壤植硅体碳积累过程中发挥着重要作用[52],因此不同覆被类型下土壤植硅体碳汇潜力就存在差异。综合前人研究结果,在不同森林类型下,毛竹林的土壤植硅体碳汇潜力为2.41×106t CO2[21,39],针阔混交林的碳汇潜力为2.11×106t CO2[21-22],常绿阔叶林为7.74×106t CO2[21-22],马尾松为4.22×106t CO2[21],杉木为2.45×106t CO2[21]。而在不同温度带下,土壤植硅体平均碳储量中,寒温带最多(表3)。

表3 不同温度带下森林植硅体平均碳储量
出现上述情况的原因可能是落叶松作为寒温带地区主要优势树种,凋落量大,因为植硅体的特性而导致叶片中的植硅体碳随着叶片的凋落归还于表层土壤,这与Blecker等[54]的研究结果一致。以竹子为主要覆被类型的土壤碳汇潜力研究中,雷竹的土壤植硅体碳汇潜力为5.29×106t CO2,占整个雷竹生态系统碳储量的97%[39,40],这与冯晟斐等[27]研究结果相似,说明土壤也是竹林生态系统植硅体碳的主要储存库。
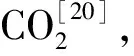
以上研究不仅表明了不同土地覆被类型之间土壤植硅体碳汇潜力的差异,也进一步说明土壤极高的碳封存潜力。植硅体碳通过凋落物释放到土壤的过程是一个长期过程,各个环节都有可能发生变化,何珊琼等[48]的研究就表明植物部分的植硅体碳汇高于土壤部分。但从以上研究对比发现,土壤中的植硅体碳汇潜力明显高于植物部分,且也有研究表明[38,48]植物中植硅体比土壤中植硅体更具活性,对环境的响应敏感,受外界和自身影响较大,因此其包裹的有机碳在进入土壤前有所释放,但土壤中的植硅体碳相对稳定,从而导致土壤中的植硅体碳汇量相比植物部分大。研究结果的不同表示,需进一步加强植硅体碳在各个环节中的变化研究,以便系统分析出现这一情况的原因。
4.2 植硅体碳在不同土壤剖面的迁移
由于植硅体体积很小,气候因素和土壤理化性质等都对典型土壤中的植硅体迁移、保存有影响,即土壤pH、土壤有机质含量、土壤有效硅含量及降水量等[58]。因此针对土壤植硅体碳垂向分布规律,早期已有学者展开了相关工作[59-60],刘利丹等[61]通过研究发现植硅体与有机质的剖面分布一样,呈现出土壤表层富集的趋势,这与Blecker等[54]在草地生态系统研究中土壤植硅体含量随剖面深度而变化的趋势相似,同时植硅体碳的浓度也呈现逐渐降低的变化趋势,即随着土壤深度的增加而减少。这主要是因为植物枯枝落叶首先进入土壤表层,且植硅体不易被分解掉,可以长期保存在土壤中,使其包裹的有机碳在土壤表层中慢慢积累,但由于受降水等因素影响,富集在表层的植硅体碳会向土壤深层不断迁移。并且在陆地生物泵的作用下,被植物循环利用并维持在一个含量相对稳定的状态,Fishkis等[10]的研究发现植硅体碳向深层土壤迁移的距离可达到2.2 m。
综合上述研究结果,今后应加强土壤内植硅体碳含量研究,同时应加强土壤各土层中植硅体碳汇潜力差异研究,以便进一步分析植硅体碳在土壤各土层中植硅体碳汇潜力的变化趋势,为整个生态系统的碳汇变化提供有力依据。
5 结论与展望
1)现有研究主要集中在森林、草地、湿地生态系统等人为干扰较少的地方,但城市生态系统中的行道树和景观植物等缺乏研究。城市作为巨大的碳源地,进一步研究城市生态系统内的植物碳汇潜力将对未来景观发展具有指导意义。
2)目前研究对于林下植被的植硅体碳汇潜力研究相对缺乏,而增加森林各层次中的植硅体碳汇潜力,将进一步提高森林的植硅体碳汇潜力。今后研究中应进一步加强林下植被的碳汇潜力研究,为造林、再造林过程中,林下植被的选择提供方向,以便进一步增强森林生态系统的碳汇潜力。
3)目前高大乔木的相关研究主要集中于亚热带,热带等环境,并且其根系部位一直研究较少,因此今后应加强单一树种,以及乔灌木等相对矮小植物研究。植物不同生境下的植硅体碳汇潜力也有不同,但关于此的研究相对缺乏,因此加强此方面的研究将补齐生态系统碳汇研究,进一步了解植物各部分,各生境植硅体碳汇潜力差异变化,对整个生态系统植硅体碳汇的精确评估具有重要意义。
4)西南喀斯特地区作为脆弱生态环境区,环境容量小[62-63],与其它亚热带森林碳汇能力存在差距,但目前关于该地区植硅体碳汇潜力研究较为缺乏。因此,挖掘喀斯特森林生态系统碳积累的自然过程和维持机制,开展西南喀斯特亚热带森林典型群落植硅体碳封存潜力的研究,将对科学估测亚热带森林生态系统长期固碳潜力提供重要依据。
