基于CCM机制的水生碳泵效应协同富营养化缓解研究进展
2022-04-01陈文瑾陆苹茹
陈 波,陈文瑾,陆苹茹,赵 敏
(1.贵州财经大学,贵阳 贵州 550025;2.中国科学院地球化学研究所,贵阳 贵州 550081)
0 引言
近年来由于人类活动的影响,大气中 CO2含量大幅增加,全球碳循环和全球气候发生变化。由于全球化石燃料的燃烧和土地利用变化等人类活动增强,大气CO2浓度已经从280 ppm升高至约415 ppm,近两百年的时间增加了48%[1]。大气、海洋、陆地生态系统之间原来的碳通量交换平衡已经打破,正在向新平衡过渡。新的碳循环平衡点,取决于人类活动和地球系统之间碳交换的物理、化学和生物过程[2]。大气CO2浓度升高,进而引起的温室效应,对全球气候变化、环境生态结构甚至人类自身的生产生活都有深刻影响。
现代碳循环研究表明,在全球尺度上估算出的大气碳源量大于碳汇量,碳源、汇收支不平衡,此部分“遗失碳汇”去向成为近年研究关注的重点[3-7]。随着研究的系统化和深入,内陆水体(包括河流、湖泊和水库等)在全球碳循环中的作用得到越来越多的关注。
尽管淡水面积只占地球陆地的很少一部分,只占到全球面积的约3.7%[8],但淡水生态系统,特别是河流、湖泊和水库在区域和全球碳循环中的极端重要性,近年来获得了广泛的关注[9-10]。全球内陆水体向大气释放CO2的量与海洋吸收的 CO2量相当,同时全球内陆水体沉积物埋藏的有机碳则超过海底有机碳汇[10]。据 Cole 等人[9]的估计,全球内陆水域每年从陆地景观获得的碳至少达到19亿t,其中2亿t被埋藏在水体沉积物中,8亿t返回大气,其余9亿t进入海洋,这与湖库的高OC沉积速率和高保存度有关,湖库是海洋的50倍[11]。而生物碳泵效应作为陆地水生生态系统长期稳定碳酸盐风化碳汇形成的关键过程。
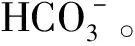
总之,基于CCM机制的水生碳泵效应是陆地水生生态系统碳汇形成的关键, 该机制影响水生生态系统的碳汇量和营养元素利用耦合效率,使得高DIC(溶解无机碳)地区的喀斯特陆地水生生态系统成为全球遗失碳汇的重要研究对象。
1 碳汇研究进展
全球变化科学的一个重要问题是全球大气 CO2的收支不平衡[3,13],即每年存在着大约10亿t的陆地遗失碳汇(Missing carbon sink),其位置、变化和机制仍然不清楚,且存在较大的争论。现在多数学者通过对δ13C-△14C值的测量、全球二氧化碳分布和总碳循环模式的研究后,认为全球的“遗失碳汇”可能存在于北半球的陆地圈中[14]。陆地生态系统的源与汇的不确定性,给未来大气中二氧化碳浓度的估算带来不确定性,也影响了对未来全球气候变化预测和评估[15]。
我国已是世界能源消费和CO2排放大国,2020年化石燃料燃烧排放CO2高达10.7 Gt,是美国化石燃料燃烧CO2排放(4.7 Gt)的两倍多[1]。作为一个负责任的大国,中国政府对于CO2的减排给予了高度重视。党的十九大报告中多次提到生态效益、减排等相关内容[16];中共中央总书记、国家主席、中央军委主席习近平在全国生态环境保护大会上明确提出“要实施积极应对气候变化国家战略,推动和引导建立公平合理、合作共赢的全球气候治理体系,彰显我国负责任大国形象,推动构建人类命运共同体。”[17]。2020年9月22日,国家主席习近平在第七十五届联合国大会一般性辩论上发表讲话,“中国将提高国家自主贡献力度,采取更加有力的政策和措施,二氧化碳排放力争于2030年前达到峰值,努力争取2060年前实现碳中和”[18]。基于理论计算和野外观测,刘再华等提出了可能存在一种由全球水循环产生的重要的CO2汇(以溶解无机碳-DIC的形式)[19]。这一研究发现,为寻找“碳失汇”提供了一个新的方向, 即将水循环、碳酸盐岩风化和水生光合作用综合考虑,对区域尺度乃至全球尺度的碳汇进行重新估算。
近20年来,对于估算岩溶碳汇量的观测研究取得了重要的成果,长期的定位观测从多角度揭示了岩溶动力系统中碳循环的运行机制,并运用多种方法估算了岩溶作用消耗大气CO2的量,其中中国岩溶作用碳汇为1.77×107t·C/a,全球岩溶作用碳汇为6.08×108t·C/a,后者约占全球碳循环模型中遗失碳汇总量的1/3[20]。随着人们对生物参与岩溶地质作用的认识[21-23],对岩溶碳汇有了新的认识,基于考虑水-岩-气-生相互作用的碳酸盐岩风化碳汇模型[19]的岩溶碳汇量显然比单纯的计算碳酸盐岩风化所产生的无机碳通量要更大[24-25]。
生物泵是海洋科学家研究海洋碳循环时提出的概念,是指由有机物生产、消耗、传递、沉降和分解等一系列生物学过程构成的碳从表层向深层的转移[26-32]。生物泵是以一系列生物为介质,通过光合作用将部分DIC转化为有机碳,之后在食物网内转化、物理混合、输送及沉降将碳从真光层传输到深层中的过程[26,30]。这样, 通过生物活动增加了海洋中碳汇的强度。
陆地水体作为全球水体的重要组成部分,相比海洋而言,其覆盖面积虽不及全球的1%[33],但因其类型的多样性,包括了众多如湖泊、河流、沼泽、溪流以及人工水库等类型,同时陆地水体中所富含的水生生物,尤其是水生光合生物,它们每年通过光合作用所固定的碳却是巨量的[19]。因此,虽然陆地水体的面积小,但其生态系统能影响区域的碳收支[9]。
内陆水体碳收支的计算主要包括4个方面:对流域碳输入通量的估算,对流域下游碳输出通量的估算,对水—气界面碳通量的估算,以及对流域内碳沉积通量的估算。由水-气界面的碳通量研究表明,水的化学性质能影响二氧化碳的平衡,软水的缓冲能力较小,硬水较大。水体中初级生产者通过光合作用水中的DIC,产生有机物和氧气,其中部分有机物沉积于水体中,因此水生系统的初级生产力对水体碳循环有重要意义。湖泊的初级生产力与水体的营养条件呈正相关,表明湖泊的营养条件对其碳收支水平有密切的关系。其影响因素包括水体的光辐射、温度、二氧化碳在水体中的平衡与转化速率、营养的含量水平、浮游植物群落特征和水动力条件等等。湖泊初级生产力水平决定了湖泊的碳循环特点和碳收支强度,即湖泊中的生物碳泵功能有着重要的作用[33]。
相比传统的陆地水生生态系统的观点,Cole等[9]提出了新的更准确估算内陆水体中的碳收支的方法:
I=G+S+E
(1)
其中I表示输入内陆水体的总碳量,G表示内陆水体向大气释放的CO2量,S表示内陆水体碳储量的变化,E表示内陆水体通过排泄释放的碳量。而Raymond等[34]则更新了Cole等人估算碳收支的方法,认为内陆水体中的碳收支变化受到流量控制,并提出了脉冲分流概念(Pulse-shunt concept)。
大气CO2浓度增加,并不一定能增加陆地生态系统的碳汇,Song等[35]认为,提高CO2的浓度并不能增加半干旱地区的草地碳汇,因为大气CO2浓度升高的同时加剧了草地土壤呼吸,抵消了草地的碳汇能力。而与陆地生态系统不同,土壤呼吸的提升能增加土壤水中溶解无机碳的浓度,同时影响岩石风化过程,最终在陆地水生生态系统中的水生植物作用下影响陆地水生生态系统碳汇格局,并产生有效碳汇[19]。DIC转运到海洋或内陆湖泊, 被浮游植物和沉水植物固定和利用,形成溶解有机碳(DOC)和颗粒有机碳(POC),再通过混合和传输作用,被底栖微生物转换成难以分解的惰性沉积物(SOM),有效沉积下来,成为长久碳汇[36]。
2 水生光合生物固碳机制

(2)
2.1 DIC 施肥效应
2.2 基于CCM的水生光合作用固碳机制
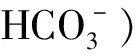
(3)
随着CO2被Rubsico固定,平衡向右移动,加速了藻类的钙化[39]。
在高DIC环境下,为了能适应其低CO2aq条件和实现水体中光合作用速率最优,演变出了CCM机制。自从1980年发现这一重要无机碳利用机制[42],在低CO2aq环境中通过CCM机制利用DIC的研究得到越来越多的关注,关于CCM机制的研究取得了重要进展[43-46]。但关于诱导CCM机制产生机理、制约CCM机制功能的主要原因等研究较少。

图1 水生植物对DIC的吸收利用途径
Rubisco作为催化CO2固定的关键酶,尽管Rubisco对CO2的亲和力低于外界大气CO2浓度,但许多光合作用的有机体具有一种能在细胞内提高CO2浓度的机制—CO2浓缩机制(CCM),依靠该机制可以在低浓度CO2环境中也能达到较高的光合速率,这种机制广泛存在于从细菌到高等植物的很多进行光合作用的有机体中[40,47]。
3 岩溶水生碳泵效应与生态系统修复
3.1 基于CCM机制的C-N-P共沉积效应
3.2 沉水植物群落恢复与富营养化改善
因此,基于CCM机制的生物碳泵效应可以作为氮磷去除,特别是磷去除的一个生态手段。Brothers等人[62]研究了浅水湖泊富营养化致生物结构演替对碳埋藏的影响,随着沉水植物的消失,水底中溶解氧下降使得埋藏有机碳的矿化速率降低,一定程度上增加碳埋藏效率,大型沉水植物向浮游植物的转变可能有利于有机碳的埋藏。另外有研究者发现,浮游植物由于繁殖世代很短,容易形成新鲜有机质,对埋藏有机碳有激发效应,加速了底泥有机质的分解和CO2释放[63]。沉水植物在CCM机制的驱动下,除了能发挥固碳和稳碳的作用,对富营养化水体的生态系统的修复能起到关键作用。例如,童昌华等[64]发现狐尾藻和微齿眼子两种菜沉水植物对水中总氮、总磷和硝态氮去除效果最好。任文君等[65]发现沉水植物对富营养化湖水、底泥具有一定的耐受能力,在试验条件中生长良好,长度与生物量都明显增加,并能有效地降低水体中的总磷、氨氮及总氮浓度。李必才等人[66]利用白洋淀优势沉水植物轮藻(Charasp.)作为建群种,研究其修复白洋淀富营养化水体的效果。结果发现轮藻覆盖度为80% 时,水体中叶绿素a、氨氮、磷酸盐和总磷含量减少最明显。
4 结论及展望
陆地水体与大气、土壤、岩石和海洋都有接触,吸收大气中的CO2同时也向大气排放CO2和CH4;土壤中的有机物和无机物淋溶经地下水和地表径流进入水中,在运移过程中,土壤中DIC、DOC、POC、PIC相互之间发生物理化学和生物转化,其中部分沉积在陆地水体中;地表上的岩石,在物理化学和生物风化的作用下溶解,除一部分向大气释放外,很大一部分是被搬运到水体中被陆地水生生态系统利用;河流中的碳,包括DIC、DOC、POC、PIC,在河口处向海洋排泄,使海水中的碳量增加。每年大约有0.6 Pg的碳被埋藏在内陆水体中,这相当于陆地生物和土壤固定碳的20%,而且这并不包括长时间尺度上的存在于河漫滩和滨海的碳汇效应[14]。生物碳泵效应作为陆地水生生态系统一种稳碳和固碳过程,是形成长期稳定碳酸盐风化碳汇的重要机制,是碳循环的重要环节。陆地水生光合生物主要通过CCM机制在有限CO2水体中提升光合固碳能力,DIC施肥效应进一步放大这一固碳机制,增加碳酸盐风化碳汇量。陆地水生生态系统中耦合营养元素与生物碳泵效应能够有效对富营养化水体环境进行改善和修复,在稳碳和固碳过程中同步实现富营养化水体的治理。
同时,随着全球CO2浓度在不断上升,促进了碳酸盐岩风化,使地表溶解更多的DIC。被溶解的DIC进入陆地水体,其中一部分被转化为其他形态的碳,一部分被水生生物固定下来,沉积到水体底部,其他的排向海洋,再被海洋水生生物利用固定[6,19]。基于CCM机制的C-N-P共沉积,在提升陆地水生生态系统的固碳能力的同时,通过调整生态系统中化学元素计量比,实现水环境综合治理,改善陆地水域环境。
总之,鉴于陆地水生生态系统的复杂性,综合考虑碳的各种存在形式,把它们来源和去处以及它们之间相互转化关系了解清楚,生物碳泵效应作为一种稳碳和固碳过程,是形成长期稳定碳酸盐风化碳汇的重要机制,是碳循环的重要环节。陆地水生生态系统中耦合营养元素与生物碳泵效应能够有效对富营养化水体环境进行改善和修复,在稳碳和固碳过程中同步实现富营养化水体的治理。这对精确地对陆地水生生态系统碳汇量进行估算,研究不同气候和土地利用对碳汇量的影响和揭示生物碳泵效应与水环境改善的相互作用机制和水生生态系统在全球碳循环中的作用具有重要意义。
