桐梓河浮游植物优势种生态位与种间联结性分析
2022-04-01李秋华马一明
李秋华,马一明
(1.贵州师范大学 贵州省山地环境信息系统和生态环境保护重点实验室,贵州 贵阳 550001;2.贵州省国际合作研究基地水生态国际联合研究中心,贵州 贵阳 550001)
0 引言
对不受管制、自由运行的河流,河流连续体的概念可谓是一个框架,其之中的生物群落结构具有纵向连通性功能。有研究显示,为调控河流的流量及排放,许多地方在沿河道上进行蓄水筑坝(全球湖泊生态系统和近58 000座大坝超过15 m高或蓄水>300万m3[1-2]),原生河流的生态过程及多样性在这些筑坝的干扰下发生了改变[3-4],为此,河流中的非生物和生物也可能发生了转移。
浮游植物的生态群落是一个由生物体和物种相互作用而形成的复杂的网络结构。一个特定的浮游植物种群不是孤立存在,而是在相应的栖息地共存,并在时间、空间上与其他种群有联系。Héctor等[5]和刘艳等[6]的研究结果均显示,生态位的宽度、位置与物种的分布有关。此外,物种间的关系(对有限资源的竞争和相互促进)也决定了群落的结构及动态,主要表现为生态位和种间关联[7-8],可见,生态位宽度和种间关联能反映种间竞争及群落的稳定程度。
众所周知,修建在长江流域上游的山区河流大坝,不仅在防洪中发挥着重要的作用,同时其转化的清洁能源水电资源,在改善环境、提高经济效益等方面也做出了巨大贡献[1,9-10]。然而,大坝对河流中水生生物在地点间的扩散却形成了阻碍(有限或过度扩散可不同程度地改变群落动态),从而改变生物的多样性模式。为深入了解不同水文时期大坝阻隔对浮游植物群落生态位和种间关联的影响,选择云贵高原梯级筑坝河流——桐梓河为研究对象,通过浮游植物的生态位变化模式,拟探讨浮游植物优势种的生态适应性、种间关联及浮游植物群落结构的演替关系,进而为水质管理和河流水生态系统的恢复提供理论参考。
1 材料与方法
1.1 调查方法
桐梓河是赤水河最大的支流,属长江水系支流,流域海拔高程介于484~1 843 m,面积3 348 km2。桐梓河属典型的雨源型山区河流,径流的时空分布变化与降水基本一致,年际变化较大,年内分配不均匀。据田义超等[11-12]研究显示,该流域内人类的活动有水土资源的开发利用、植被恢复、水利工程建设等,人类活动的变化规律可归结为土地利用的变化,且是人类活动的主要表现形式。本研究于2020年8月(丰水期)、2020年11月(枯水期)和2021年4月(平水期)对桐梓河浮游植物群落结构进行调查,在主河道共设25个采样点(见图1),每个采样点只对表层进行采样。

图1 桐梓河采样点分布
1.2 样品的采集与处理
浮游植物定量样品:在每个采样点的表层取1.5 L的水样后,立刻加入15 mL的鲁哥氏液固定;带回实验室静置24~48 h后用虹吸法浓缩至30 mL,再使用0.1 mL浮游生物计数框在10×40倍光学显微镜下进行观察、鉴定和计数。鉴定参考的主要资料为《中国淡水藻类:系统、分类及生态》[13]和《中国内陆水域常见藻类图谱》[14]。
1.3 数据分析及处理
由Mcnaughton优势度指数(Y)计算优势度[15],公式为:
(1)
式中:ni为样点中第i种浮游植物的个体数,N为样点中所有浮游植物的总数,fi为该种属在各样点中出现的频率。
Levins的生态位宽度指数能确定一个群落中所利用的各种不同资源的总和[16]。其公式为:
(2)
式中,Bi为物种i的生态位宽度;Pij为物种i在给定地点j中的占比;r为采样点。如果物种具有较高的B值,表明该物种的生态位宽度更高,其分布更广泛、更均匀、规模更大;反之物种的B值越低,则说明物种分布越少,且不均匀。当物种i的个体以相等的数目利用每一资源状态时,B值最大化,即该种具有最宽的生态位;当物种i的所有个体都集中在某一个资源状态下时,B值最小,该种具有最窄的生态位。
用Pianka指数计算生态位重叠值(Oik)[17],公式为:
(3)
(4)
(5)
式中,Oik为重叠值,Pij和Pkj分别表示种i和种k在第j个站点中的个体数占该种所有个体数比例,r为资源状态总数。Oik取值范围为[0, 1],表示种i与种k的资源利用曲线的重叠指数。当ΔOik>0,说明该种处于发展期;ΔOik<0,该种则处于衰退期;ΔOik=0,该种是中性的;R表示生态响应速率。
采用Schluter的方差比率法(R)检验物种间总体关联性[18],公式为:
(6)
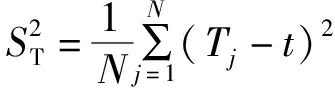
(7)
(8)

关联系数(AC)用于量化每连个物种间的种间关联[7],公式为:
(9)

(10)

(11)
式中:a为1对种共同出现的采样点数,b、c为单个种出现的采样点数,d为均不出现的采样点数。AC范围为[-1, 1];AC=1表示物种间最强的正种间关联,AC=-1表示物种间最强的负种间关联,而AC=0表示物种对是完全独立的。
2 结果与分析
2.1 浮游植物优势种群落结构特征
通过实验室镜检分类鉴定,3个水文时期桐梓河共有浮游植物7门75种(见图2)。其中,绿藻门物种数最多,为33种(占比44.0%),硅藻门次之,为24种(占比32.0%),蓝藻门10种(占比13.3%),前3类种群占总种数的89.3%,为主要的群落组成结构,其他种群占比较少,甲藻门和裸藻门各为3种(分别占比4.0%),隐藻门和金藻门各为1种(分别占比1.3%)。

图2 桐梓河浮游植物物种类组成比例
优势度前10的物种为优势种。由表1可见:桐梓河3个水文时期共鉴定出浮游植物优势种5门15种,硅藻门为主要的优势种群,其中包括梅尼小环藻(Cylotellameneghiniana),小型异极藻(Gomphonemaparvulum),尖针杆藻(Synedraacusva)和舟形藻(Naviculasp.),以及隐藻门中的啮齿隐藻(Cryptomonaserosa);优势种的种类和优势度在不同的水文时期均不同,其中梅尼小环藻在3个水文时期的丰度最高,平均丰度为3.82×105cells·L-1,平均优势度为0.303;部分藻类具有较强的季节适应性,仅在某一水文时期成为优势种,且丰度和优势度皆占比较小。

表1 浮游植物优势种丰度(D,×104 cells·L-1)及优势度(Y)
不同水文时期浮游植物优势种丰度和优势度的变化趋势(见图3):整体来看,丰水期浮游植物优势种的平均丰度最高(1.53×105cells·L-1),其次是平水期(9.49×104cells·L-1),最后是枯水期(5.18×104cells·L-1),优势种的优势度与水文期具有相同的趋势;与丰度趋势相反的是,丰水期优势种间的相对丰度差异最大,枯水期时最小,这说明河流水量越大,浮游植物群落具有明显的差异性,群落结构越不稳定。
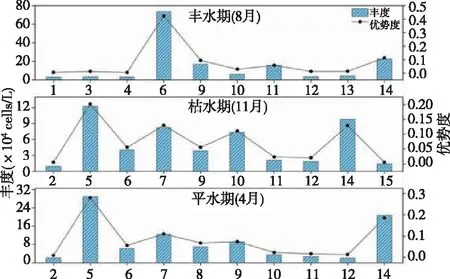
图3 不同水文时期的优势种丰度(D)及优势度(Y)
2.2 浮游植物生态位
2.2.1 优势种生态位宽度
3个水文时期浮游植物优势种生态位宽度的变化情况(见表2):15个优势种生态位宽度变化的范围为[2.04, 14.90],其中在平水期的微小曲壳藻的生态位最宽(14.90),丰水期的微小四角藻的生态位宽度最窄(2.04);微小曲壳藻、扁圆卵形藻、小型异极藻和舟形藻的生态位宽度相对较高,生态位宽度值大于7.0,说明该类优势种利用资源能力强,具有较广的生态适应幅度;蹄形藻、梅尼小环藻、线型菱形藻、尖针杆藻和啮齿隐藻,其生态位宽度小于第一类,对生境有一定的要求,对环境资源具有较强的选择性,在合适的生境下能较好生存;普通小球藻、湖生卵囊藻、微小四角藻、弧形短缝藻、盾形多甲藻和分歧锥囊藻,仅在某一时期成为优势种,该类特种只在环境较适合的水文时期出现,对水环境条件变化较为敏感,对生态环境的指示作用明显。

表2 浮游植物优势种生态位宽度(Bi)
对比3个水文时期各个物种的生态位宽度(图4)发现:丰水期间浮游植物的平均生态宽度最低(Bi=2.95),窄生态位(Bi<2)的物种有33种,占比最高(55.9%),这可能是过快的水流速度影响了浮游植物稳定群落结构的形成,使得多数种群表现出窄生态位情况;在枯水期和平水期,低水流量下出现了部分稳定的水体生境,浮游植物群落向广生态位转移,但受大坝阻隔大部分物种的生态位宽度值在4~8之间,形成了一定的生态隔离。

图4 不同水文时期浮游植物生态位宽度(Bi)
2.2.2 优势种生态位重叠
利用生态位重叠指数分析不同水文时期对浮游植物优势种的重叠关系(见表3):不同种类的物种对3个时期表现出的生长适应有明显差异;15个优势种的生态位重叠值Oik变化区间为[0.008, 0.918];重叠种群的最小值出现在丰水期(微小四角藻-小型异极藻),2个物种间基本不相交,最大值在平水期(尖针杆藻-线型菱形藻),说明在平水期2个群落之间大部分重叠。
ΔOij的大小展示的是不同环境条件下不同种属的生态习性变化及发展衰退状况,为更准确地确定不同时期优势种对生境条件的生态响应,结合生态位宽度计算生态响应速率R(见表3):蹄形藻、梅尼小环藻和啮齿隐藻在枯水期和平水期表现出衰退的群落模式,微小曲壳藻、小型异极藻和线型菱形藻在枯水期和平水期表现出发展的群落模式;丰水期的优势种群与枯水期和平水期表现出相对的生长模式。基于R值进一步确定不同时期优势种对生境条件的生态响应状况,发现:在生态位重叠程度最大时均处于衰退状态,且衰退空间较大。

表3 浮游植物优势种生态位重叠(ΔOik)和生态响应速率(R)
2.3 浮游植物优势种的种间联结性
桐梓河3个水文时期的VR值均大于1,且统计量W均大于(df)(见表4),表明浮游植物优势种间总体上均呈显著正关联。

表4 浮游植物优势种总体关联性
联结系数(AC)分析结果显示(见图5):丰水期、枯水期和平水期AC≥0.5的种对分别为8,3,3对,分别占总对数的17.8%,6.67%,6.67%,说明种对间正联结的程度较低;0≤AC< 0.5的种对分别为17,15,18对,分别占比37.8%,33.3%,40.0%,说明种对间正联结的程度较弱。-0.5≤AC< 0 的种对分别为6,10,5对,分别占比13.3%,22.2%,11.1%。AC≤-0.5的种对分别为14,17,19对,分别占比31.1%,37.8%,42.2%,说明种对间负联结的程度较高。
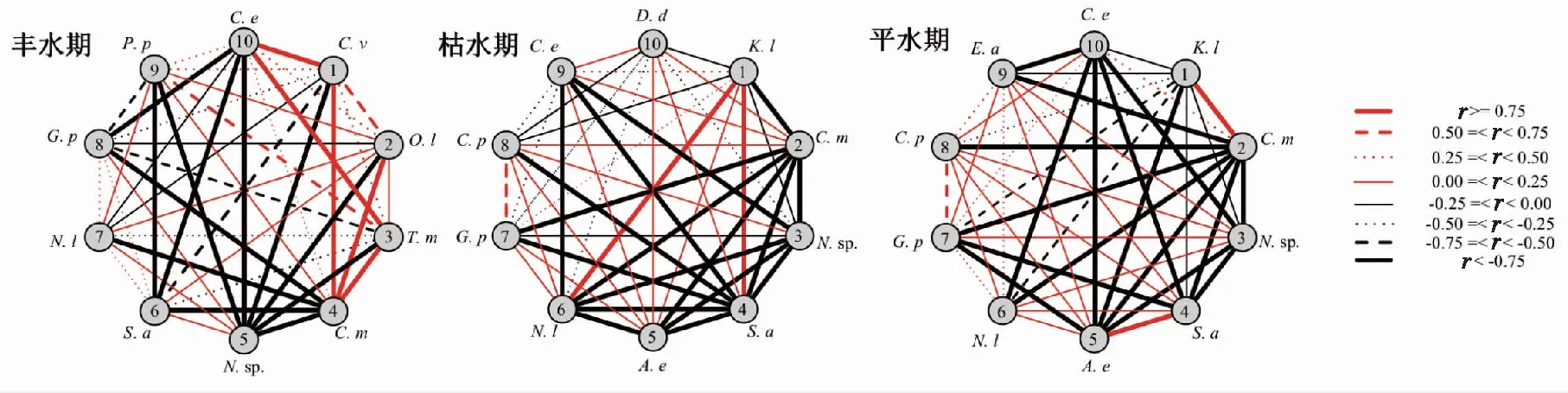
图5 浮游植物优势种间AC检验
3 讨论
有研究发现,生态位宽度可反映浮游植物群落中各物种的地位及作用[19-20]。本研究结果表明,硅藻门中的微小曲壳藻、扁圆卵形藻、小型异极藻和舟形藻生态位宽度较大,在群落环境中起着重要作用,即在群落中的竞争能力和利用资源能力方面较强。李兴等[21]通过监测生态位宽度较大的藻类状况,判断出河流水质的本质变化;杨文焕等[22]研究显示,生态位宽度较小的藻种,往往会在特定的时间和空间条件下出现,对生境的变化比较敏感,生境特化程度高,可作为水域的指示种(利用同一物种在不同水文时期下的生态位宽度变化的特性,该物种作为水体的指示种具有一定的意义);De souza等[23]研究发现,在不同水文时期,因温度、水流量大小不同,大坝的阻隔会对浮游植物的结构产生明显影响。本研究显示,硅藻在低温条件下能较好地生存,相似水温下优势种的组成也比较相似,温度的差异决定了优势种的组成结构。在对比研究流域内浮游植物种群生态位宽度的结果时,本研究发现:在丰水期,浮游植物群落的平均生态位宽度最低(Bi=2.95),窄生态位的物种占比最高(55.9%),这是因为在雨季河流的水流量和流速较大,浮游植物群落受环境的影响较大,多数种群难以适应不稳定的水体生境,故不能形成稳定的群落结构,进而导致窄生态位的物种占比较高;在枯水期,浮游植物群落的平均生态位宽度(Bi=4.08)高于丰水期,这是因为大坝阻隔使得河流的部分水体流速缓慢,较为稳定的水体给大多浮游植物群落提供了一定的空间资源,加之部分河段断流形成的短时间的地理隔离,进而使得该时期浮游植物群落的平均生态位宽度高于丰水期;在平水期,浮游植物群落的平均生态位宽度(Bi=4.13)最高,该时期大坝对浮游植物群落的水生环境影响相比丰水期和枯水期最小。
有研究显示,生态位重叠的结果与物种的生态位宽度有较大的联系,而这些有着相似生态特性的物种会共同开发同样的生境资源,进而导致浮游植物群落发生显著的生态位重叠特征[24-26]。本研究发现,在丰水期,多数优势种之间的生态位重叠值较低(重叠值高于0.5的物种对占比17.8%)。说明优势种占据的生境资源不在同一空间,受外界干扰严重(即水流量较大时对浮游植物群落的破坏较为严重),使得不同优势种难以形成同一稳定的生存环境[27]。与生态位宽度表现出相似结果的是枯水期比丰水期具有更高的生态位重叠值(重叠值高于0.5的物种对占比26.7%),生态位重叠值在平水期最高(重叠值高于0.5的物种对占比31.1%)。在生态位重叠分析中,本研究发现不同优势种表现出的生长状态不同,梅尼小环藻、小型异极藻和线型菱形藻在丰水期表现出的是积极的生长模式,枯水期和平水期是衰退状况,舟形藻和啮齿隐藻则在丰水期表现出衰退状况,枯水期和平水期则相反。说明不同优势种对不同水文时期的适应能力不同,从这些物种的生长情况可判断出生境变化带来的生态影响[22]。
联结系数(AC)是对3个水文时期优势种物种的联结性进行具体测度,进而得到更加精确的结果[28]。在AC分析中,3组水文时期状态下的物种之间整体表现出负关联(正关联和负关联的平均值分别为0.318和-0.736),正关联占比分别为51.1%、40.0%和46.7%,表明研究流域内大部分物种在研究期间具有相似的生物学特性或生境需求,而生态位重叠值较低,说明物种之间的共用资源充足、竞争较小[29]。一般来说,群落正向演替程度提高,群落结构和组成种类数会趋于稳定,物种间也会趋向正关联,这与AC统计结果相吻合。在湖泊生态系统中,往往生态位重叠值高的物种间的正联结占比较高,但在大坝阻隔的桐梓河的河流生态系统中,却表现出相反的结果,这可能是该河流中物种的总丰度较低,不存在资源竞争情况。求锦津等[30]研究显示,物种间的群落结构模式完全受河流的水流大小影响,外界影响的因素差异导致了物种的关联方式不同。此外,在优势种之间的联结性检验结果中,本研究还发现同一种群很难适应3种不同的水流量状态,生境的改变直接引起了群落结构的改变,这可能与季节的变化有关,但AC值显示物种间长期存在高度负关联,相比丰水期,枯水期对浮游植物群落的隔离更加严重。
