急性脑梗死发生过程中lncRNA TALNEC2的作用与机制
2022-03-31李岳勇蒙兰青邱绍财
李岳勇,蒙兰青,黄 清,李 东,邱绍财
急性脑梗死的临床治疗主要是早期静脉溶栓治疗,时间窗在发病后3.0~4.5 h内[1]。但大多数脑梗死患者不能及时接受溶栓治疗,预后较差。因此,进一步研究脑缺血的病理生理机制具有重要且迫切的临床价值。研究[2-4]表明,长链非编码RNA(long non-coding RNA,lncRNA)在大脑中高度表达,与多种神经疾病有关,如自闭症谱系障碍和癫痫。TALNEC2作为一种促进细胞损伤的lncRNA,研究发现胶质母细胞瘤中TALNEC2增加[5]。此外,心肌缺血患者血清和缺氧损伤的H9c2细胞中TALNEC2增加[6]。然而,TALNEC2在急性脑梗死中的作用尚未阐明。该研究通过体外脑微血管内皮细胞(brain microvascular endothelial cell,BMEC)暴露于缺氧-葡萄糖剥夺(oxygen-glucose deprivation,OGD)模型,确定TALNEC2在急性脑梗死中的表达和作用,并探讨TALNEC2基因敲低对小鼠大脑中动脉阻塞(middle cerebral artery occlusion,MCAO)模型神经功能恢复的影响,旨在为探索急性脑梗死的治疗靶点提供新的见解。
1 材料与方法
1.1 BMEC培养与OGD模型建立将成年雄性C57BL/6J小鼠(雄性,体质量25~30 g,10周龄,北京维通利华实验动物技术有限公司)的大脑皮层均质化,过滤,然后用胶原酶B,胶原酶/Dispase酶(瑞士Roche公司)依次消化,并在40% Percoll溶液中离心。收集含有微血管的第2条带并将其置于胶原涂层的培养皿上。小鼠BMEC(3代,基于因子Ⅷ的表达,纯度>95%,表现出缓激肽受体功能)在使用前生长到85%~95%的融合率。为了在体外模拟缺血样条件,将小鼠BMEC培养物转移到温度控制(37±1)℃的厌氧室(美国Forma Scientific公司),其中含有5% CO2和95% N2的混合气体,并用去氧无糖Hanks平衡盐溶液(美国Invitrogen公司)代替培养基,细胞在厌氧室中保持16 h,对照组小鼠BMEC不暴露于OGD。
1.2 细胞转染广州锐博生物科技有限公司合成了si-TALNEC2、si-NC、pcDNA、基于pcDNA的TALNEC2过表达载体(TALNEC2)、miR-19a-3p模拟物(miR-19a-3p)和miR-NC(miR-NC)。建立OGD模型前,使用Lipofectamine 3000(美国Invitrogen公司)将上述慢病毒或载体转染入细胞中,孵育48 h。
1.3 小鼠MCAO模型的建立采用C57BL/6小鼠建立MCAO模型。将小鼠随机分为4组:溶媒对照组(Sham)、MCAO组、MCAO+siRNA阴性对照组(MCAO+si-NC)和MCAO+siRNA抗TALNEC2组(MCAO+si-TALNEC2),每组12只。在建立MCAO模型前24 h将si-TALNEC2或si-NC慢病毒载体注入小鼠左心室。MCAO手术参照文献方法[7]建立,麻醉后,从颈外动脉至颈内动脉插入5-0外科尼龙单丝缝合线以闭塞大脑中动脉。MCAO手术60 min后,取下缝合线。Sham组采用相同的手术方法,无MCAO。于MCAO后1、3、7和14 d进行神经功能评分,取脑组织进行TTC染色分析。
1.4 神经功能评分和梗死体积的测量在手术后的1、3、7和14 d进行了足误测试以评估MCAO治疗后小鼠的神经功能,低神经评分显示神经功能缺损[7]。在手术后14 d,将小鼠麻醉,收集大脑,切割成2 μm厚的切片,并用2%TTC溶液(美国Sigma公司)在37 ℃黑暗中染色15 min。使用图像实验室软件(美国Bio-Rad公司)分析梗死组织(梗死组织呈白色)的面积。
1.5 qRT-PCR分析使用TRIzol试剂(美国Invitrogen公司)分离总RNA,并使用M-MLV逆转录试剂盒(美国Thermo Fisher公司)或TaqMan microRNA逆转录试剂盒(美国Applied Biosystems公司)将其逆转录为cDNA。然后,使用SYBR green(Applied Biosystems)将cDNA用于qRT-PCR分析。以β-actin或U6小RNA为内源性对照,用2-ΔΔCt法计算RNAs的相对表达。引物序列见表1。

表1 引物序列
1.6 Western blot实验从脑组织或BMEC细胞中提取的蛋白质用BCA蛋白质分析试剂盒(上海Beyotime公司)定量,在98 ℃变性10 min,然后通过SDS-PAGE凝胶电泳分离并转移到聚偏二氟乙烯膜(美国Millipore公司)上。在室温下用5%脱脂牛奶封闭膜1 h,然后在4 ℃下用抗Bcl-2(1 ∶1 000稀释,英国Abcam公司)、Bax(1 ∶2 000稀释,英国Abcam公司)、JNK(1 ∶5 000稀释,英国Abcam公司)和β-actin(1 ∶5 000稀释,英国Abcam公司)一级抗体孵育过夜。随后,在室温下与辣根过氧化物酶偶联二级抗体(1 ∶10 000稀释,英国Abcam公司)孵育2 h,并与增强化学发光显色底物(上海Beyotime公司)相互作用。用图像实验室软件(Bio-Rad)对蛋白质信号进行分析。
1.7 ELISA试验采用ELISA试剂盒(美国Sigma公司)测定脑组织匀浆或细胞培养液中炎性细胞因子[白细胞介素(interleukin,IL)-1β、IL-6和肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)]的释放量。
1.8 细胞凋亡采用流式细胞仪检测Annexinv-FITC/PI凋亡检测试剂盒(美国Sigma公司)检测细胞凋亡。用PBS洗涤处理后的BMEC细胞,加入200 μl结合缓冲液中再悬浮,然后在黑暗中用10 μl Annexin V-FITC和5 μl PI双重染色15 min。用流式细胞仪(美国Becton Dickinson公司)分析凋亡细胞。
1.9 荧光素酶活性测定用starBase预测miR-19a-3p与TALNEC2(全长572 bp,第358~383位碱基存在miR-19a-3p的潜在结合位点)或JNK(全长663 bp,第472~511位碱基存在miR-19a-3p的潜在结合位点)的结合位点,设计成对PCR扩增引物(TALNEC2 F:5′-AUUUGACUAAGUCAAGUUGCUC UCA-3′,R:5′-UUGGGUAGUACCUUAGAAGAGU-3;JNK F:5′-TCAAAGGGGGAGAGGGGAGAATGTTTG TAACAGGGTAGAGTGGAGGGGTGGGGGGAGCCCT-3′,R:5′-AGGGCTCCCCCCACCCCTCCACTCTACCC TGTTACAAACATTCTCCCCTCTCCCCCTTTGA-3′),构建含有miR-19a-3p应答序列的TALNEC2和JNK的3′UTR片段。扩增含有野生型(WT)或突变型(MUT)结合位点的TALNEC2或JNK序列,然后插入pmirGLO载体(美国Promega公司)以构建荧光素酶报告载体(TALNEC2-WT,TALNEC2-MUT,JNK-WT或JNK-MUT)。使用Lipofectamine 3000将20 ng荧光素酶报告载体、15 ng对照载体和40 nmol/L miR-19a-3p或miR-NC共转染BMEC细胞。转染48 h后,使用荧光素酶检测试剂盒(美国Promega公司)测定荧光素酶活性。
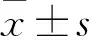
2 结果
2.1 TALNEC2基因敲低对MCAO模型脑损伤的抑制作用为探讨TALNEC2在急性脑梗死中的作用,建立小鼠MCAO模型。在MCAO后1、3、7和14 d,小鼠脑组织中TALNEC2的表达异常增强(F=38.042,P<0.001)(图1A),与MCAO+si-NC组比较,MCAO+si-TALNEC2组的TALNEC2丰度降低(图1B)。观察各组神经损伤情况,结果显示:与Sham组比较,MCAO处理增加了梗死体积,降低了神经功能评分,而TALNEC2的敲低逆转了这一结果(图1C、D)。MCAO组Bcl-2蛋白表达受到抑制,Bax蛋白表达增加,而TALNEC2的敲低使其减弱(图1E)。此外,MCAO组脑组织中IL-1β、IL-6和TNF-α的蛋白水平升高,TALNEC2的敲低使其减弱(图1F)。

图1 敲低TALNEC2抑制MCAO模型脑损伤
2.2 miR-19a-3p与TALNEC2结合为了阐明TALNEC2调控急性脑梗死的潜在机制,研究用starBase检测了TALNEC2的潜在结合miRNA,并显示TALNEC2和miR-19a-3p存在结合位点(图2A)。随后,通过荧光素酶活性分析探讨了这种相互作用。结果表明,miR-19a-3p的上调抑制了转染TALNEC2-WT的BMEC细胞中的荧光素酶活性,而TALNEC2-MUT组没有活性变化(图2B)。
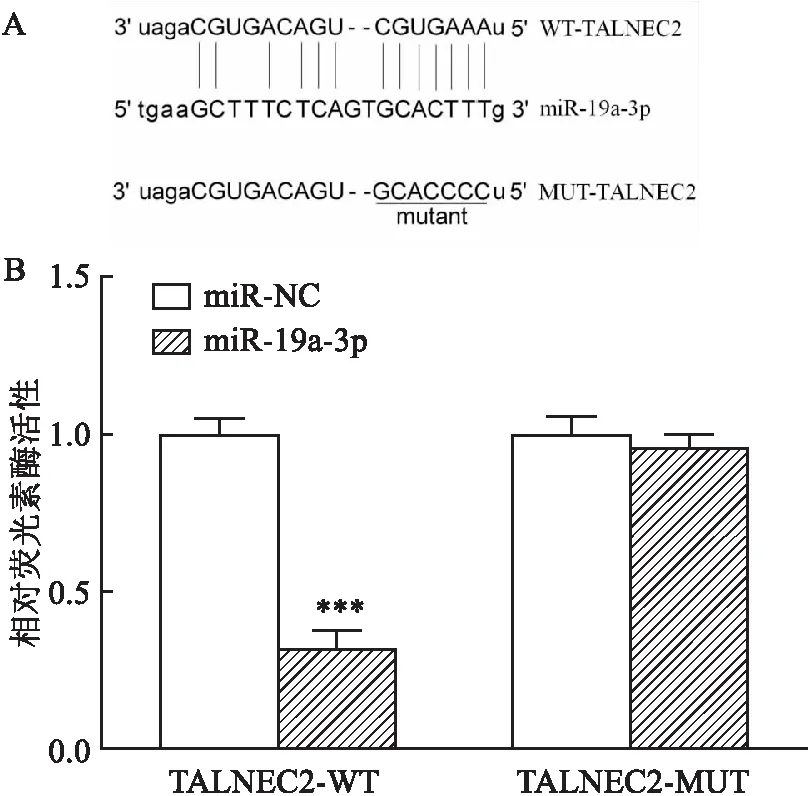
图2 miR-19a-3p与TALNEC2结合
2.3 TALNEC2通过调节miR-19a-3p介导OGD诱导的细胞凋亡和炎症损伤OGD处理后,BMEC细胞中TALNEC2的表达异常增强,而miR-19a-3p水平呈时间依赖性下降(F=23.591,P<0.001)(图3A~C)。为探讨TALNEC2介导的急性脑梗死是否与miR-19a-3p相关,将BMEC细胞分别转染miR-NC、miR-19a-3p、miR-19a-3p和pcDNA或TALNEC2,然后用OGD处理。OGD治疗后BMEC细胞凋亡和炎性细胞因子产生增多,提示体外成功建立了急性脑梗死模型(图3D)。miR-19a-3p的上调抑制OGD诱导的BMEC细胞凋亡,TALNEC2的加入减弱了这种作用。此外,TALNEC2的加入减弱了miR-19a-3p对OGD处理的BMEC细胞中IL-1β、IL-6和TNF-α分泌水平的抑制作用。
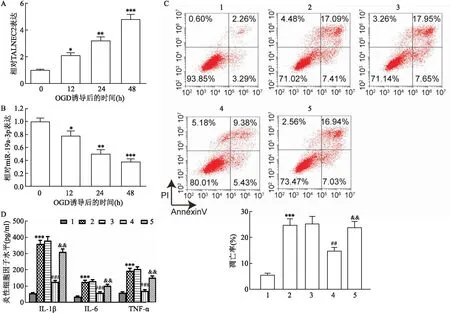
图3 TALNEC2通过调节miR-19a-3p介导OGD诱导的细胞凋亡和炎症损伤
2.4 miR-19a-3p通过靶向JNK抑制OGD处理的细胞凋亡和炎症反应为探讨miR-19a-3p参与急性脑梗死的机制,用starBase预测miR-19a-3p的潜在靶点。图4A显示了miR-19a-3p和JNK的假定结合位点。进行了荧光素酶活性测定以证实这一预测,在转染JNK-WT的细胞中,miR-19a-3p的过度表达导致BMEC细胞中荧光素酶活性降低;但在JNK-MUT组中它未影响活性(图4B)。体内研究证实了上述结果,在MCAO后1、3、7和14 d,小鼠脑组织中miR-19a-3p的表达明显减弱(F=27.140,P<0.001),而JNK的表达异常增强(F=41.503,P<0.001)(图4C、D)。经OGD处理后,BMEC细胞中JNK mRNA的表达呈时间依赖性异常增加(F=31.764,P<0.001)(图5A)。此外,经OGD处理后,BMEC细胞中Bcl-2蛋白表达呈时间依赖性异常降低(F=46.514,P<0.001),而Bax蛋白表达呈时间依赖性异常增加(F=61.802,P<0.001)(图5B)。为探讨JNK是否参与miR-19a-3p介导的急性脑梗死损伤,将BMEC细胞分别转染miR-NC、miR-19a-3p、miR-19a-3p和pcDNA或JNK,然后用OGD处理,结果JNK上调减弱了miR-19a-3p对OGD处理的BMEC细胞的保护作用,并抵消了miR-19a-3p对OGD处理的BMEC细胞中IL-1β、IL-6和TNF-α水平的抑制作用(图5C、D)。
图4 miR-19a-3p的潜在靶点

图5 miR-19a-3p通过靶向JNK抑制OGD处理的细胞凋亡和炎症反应
2.5 TALNEC2通过竞争性地结合miR-19a-3p促进JNK的表达为了进一步阐明TALNEC2处理的潜在ceRNA网络,通过转染JNK-WT分析BMEC细胞中的荧光素酶活性。结果表明,TALNEC2的引入逆转了miR-19a-3p对细胞中荧光素酶活性的抑制(图6A)。此外,Western blot分析显示,TALNEC2的过度表达导致JNK蛋白水平增加,而miR-19a-3p的加入减弱了JNK蛋白水平(图6B)。
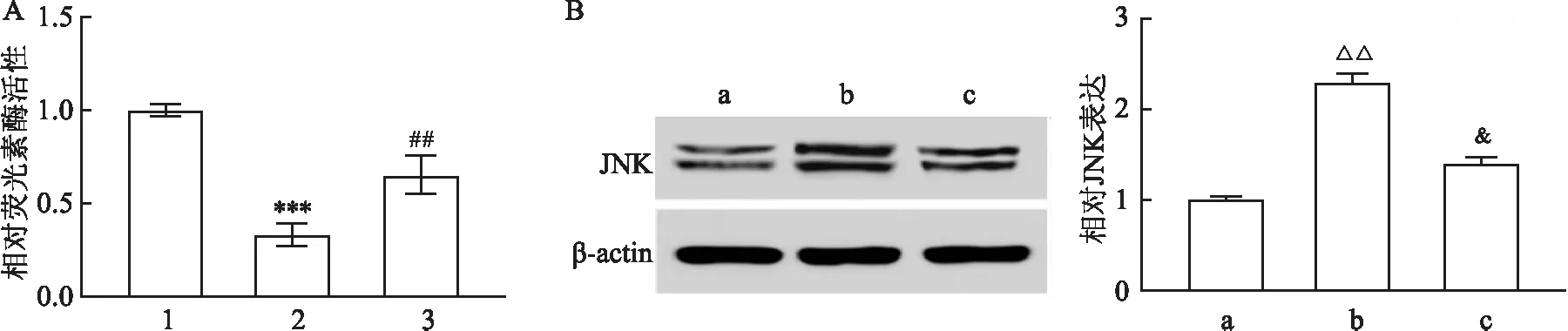
图6 JNK由TALNEC2和miR-19a-3p调节
3 讨论
lncRNA的异常表达与多种脑部疾病有关,如神经退行性疾病和脑血管疾病[8]。研究[9]发现,lncRNA转移相关的肺腺癌转录本1具有抗凋亡作用,并影响缺血性卒中后的脑损伤。lncRNA INK4位点反义非编码RNA在体外保护细胞免受OGD治疗急性脑梗死模型诱导的损伤[10]。因此,阐明lncRNA在急性脑梗死中的作用是非常有价值的,可以为其治疗提供新的治疗靶点。本研究在建立急性脑梗死动物模型和细胞模型中,发现TALNEC2在脑组织和BMEC细胞中的表达上调,并证明了TALNEC2作为miR-19a-3p的ceRNA来调节JNK,在体内和体外促进急性脑梗死损伤。
研究[5]表明,TALNEC2在胶质母细胞瘤疾病中上调,与胶质瘤干细胞的细胞增殖和自我更新呈负相关。此外,TALNEC2能够独立有效地加重H9c2细胞的心肌缺血损伤[6]。本研究中,在MCAO处理的小鼠和OGD处理的细胞中,TALNEC2均升高,促进了急性脑梗死的细胞凋亡。此外,本研究还揭示了TALNEC2的抑制作用可减少脑梗死,提高神经功能评分,提示TALNEC2的敲低在急性脑梗死中起到了神经保护作用。先前的研究[11-12]证实了TALNEC2具有加重缺氧损伤的功能,并且与细胞中的促炎介质相关。因此,本研究假设TALNEC2也促进了急性脑梗死炎症反应的产生,结果发现TALNEC2对促炎细胞因子(IL-1β、IL-6和TNF-α)的表达具有积极的调节作用,表明TALNEC2参与了急性脑梗死的炎症损伤。然而,TALNEC2在急性脑梗死中的潜在作用机制需要更多的研究。这项工作显示了lncRNA-miRNA调节网络在急性脑梗死后脑组织中的重要性。例如,有报道[13]称TALNEC2通过结合miR-650来调节凋亡肽酶激活因子1,从而加重脑缺血/再灌注损伤。为了明确TALNEC2在急性脑梗死中是否作为miRNA海绵,研究对潜在的miRNA进行了探索,结果显示miR-19a-3p含有假定的结合位点。随后,通过荧光素酶活性验证了TALNEC2和miR-19a-3p之间的相互作用。
研究[14-15]表明,miR-19a-3p通过调节缺血脑的炎症和存活而成为神经保护靶点。本研究揭示了miR-19a-3p在MCAO模型脑组织和OGD处理的BMEC细胞中下调,并且其上调减轻了OGD诱导的细胞凋亡和炎症损伤。此外,TALNEC2的加入逆转了miR-19a-3p介导的对急性脑梗死损伤的抑制作用,表明TALNEC2通过结合miR-19a-3p来调节急性脑梗死。已知功能性miRNA可调控相关靶基因的表达。以往的研究[16]表明,miR-19a-3p在不同的条件下负调控JNK的表达。因此,本研究认为miR-19a-3p/JNK的调控网络可能在急性脑梗死过程中起关键作用。通过荧光素酶活性证实了miR-19a-3p和JNK之间的相互作用。研究[17]表明,JNK逆转了miR-150-5p介导的对急性脑梗死神经元死亡的抑制作用。此外,JNK通过激活NF-κB诱导神经炎症,并促进神经元死亡[18]。本研究发现,在MCAO和OGD模型中,JNK表达升高,与先前的研究一致[19]。JNK的恢复减弱了miR-19a-3p对急性脑梗死损伤的保护作用,表明miR-19a-3p通过靶向JNK介导急性脑梗死的进展。此外,TALNEC2上调促进了JNK表达,而miR-19a-3p则削弱了这一作用。表明TALNEC2可能是miR-19a-3p抑制BMEC细胞中JNK表达的一种ceRNA。
综上所述,在体内外脑缺血模型中,TALNEC2的表达均增加。TALNEC2的敲低通过调节miR-19a-3p/JNK减轻MCAO或OGD诱导的急性脑缺血损伤,为急性脑梗死的治疗提供了一种新的潜在策略。
