基于同质园实验的两种典型荒漠植物叶片功能性状变异研究
2022-03-28单立山解婷婷种培芳师亚婷
马 丽, 单立山, 解婷婷, 种培芳, 杨 洁, 师亚婷
(甘肃农业大学林学院, 甘肃 兰州 730070)
植物功能性状是指植物在个体水平上所表现出的植物形态、生理等特征,能够有效表达植物在长期进化过程中对外界环境变化的响应[1-2]。叶片作为植物地上部分的主要器官,是植物进行光合作用和物质生产的主要场所[3],其功能性状如叶片含水量、比叶面积、叶片碳氮磷含量和解剖结构等最能体现植物对环境因子的适应性进化[4-6]。目前,植物功能性状研究已成为植物功能生态学研究中的前沿和热点问题之一[7-8]。虽有大量研究报道了植物叶功能性状变异及其与环境因子的关系[9-11],但并不能说明植物叶功能性状是受环境影响还是遗传因素的影响,而同质园实验能够在一定程度上控制环境因素所造成的影响,进而说明不同种源植物叶功能性状的变异是由于原种源地环境差异造成的植物遗传性基因变异。因此,基于同质园实验探究植物叶功能性状的变异及其与环境因子的关系具有重要意义,有助于我们更好地理解不同种源地植物对环境梯度变化的响应和适应机制。
植物叶片通过改变自身的形态和化学元素来适应环境变化[12-13]。目前,关于植物叶功能性状受环境因子影响的研究主要集中在全球等大尺度层面[14],但这种研究会忽略一些小尺度层面的环境因子如微地形、生境等,对这些环境因子的研究也许能够揭示大尺度下所无法解释的环境过滤效应[15]。植物为适应环境会形成以该环境为主的特定地理种源,遗传变异就是植物为适应该环境变化的适应结果,而由于短期环境因子如土壤质量等引起的植物叶功能性状的变异是植物适应短期环境变化的结果[16]。此外,在以往试验中沿自然环境梯度采样时,很难分辨植物表现型、基因型和环境之间的关系,如黄河上游的宁夏平原和黄河下游的黄河三角洲地区的芦苇(Phragmitesaustralis)植物叶功能性状的差异同时受到环境和遗传两个因素的共同影响[17];全缘叶绿绒蒿(Meconopsisintegrifolia)随着海拔升高,株型变矮,叶片组织结构紧密度增大[18]。然而,同质园实验将不同生境的物种栽培在环境因素相对一致的同一园地内,进行统一栽培管理,使其生境保持一致;该方法能够减少立地条件和气候变化造成的影响,在局部适应性研究中发挥重要作用[19]。近年来,虽然有关同质园的生态学研究已有诸多的报道,但主要集中在森林植物叶功能性状变异和对环境的适应性研究等方面[20-21],而关于荒漠植物在其叶功能性状变异及影响机制方面的研究则较少。
白刺(Nitrariatangutorum)和红砂(Reaumuriasoongarica)是半干旱、干旱地区常见的盐生、旱生荒漠植物,具有抗旱、抗寒、耐高温和耐盐碱等特性以及防风固沙、水土保持等作用[22-23]。白刺和红砂广泛分布于我国西北荒漠地区,并在当地形成了以它们为建群种或优势种的荒漠群落[23]。以往对白刺和红砂的研究多集中在自然环境梯度下叶片形态解剖、根系的形态和功能、对极端环境的适应性及群落功能等方面[24-27],但是采用同质园实验方法对不同种源两种典型荒漠植物叶功能性状的研究相对较少。因此,本研究分析了不同地理种源两种典型荒漠植物叶片的肉质化程度(Degree of fleshiness,DOF)、含水量(Water content,WC)、组织密度(Tissue density,TD)、比叶面积(Specific leaf area,SLA)、碳(Carbon,C)含量、氮(Nitrogen,N)含量、磷(Phosphorus,P)含量、碳氮比(Carbon and nitrogen ratio,C∶N)、碳磷比(Carbon and phosphorus,C∶P)和氮磷比(Nitrogen and phosphorus ratio,N∶P)指标,旨在探讨不同地理种源两种典型荒漠植物叶功能性状在种源间是否存在差异以及其与种源环境因子的关系,以期为两种典型荒漠植物选择优良种源、适宜的种植环境及两种典型荒漠植物对环境适应策略等提供理论依据。
1 材料与方法
1.1 研究地概况
同质园位于甘肃省武威市凉州区羊下坝镇甘肃武威市良种繁育中心(38°24′ N,103°9′ E),地处河西走廊北部,属于典型的温带大陆性荒漠气候,干旱少雨,土壤性质为典型砂壤土,pH值为8.2,有机碳含量为14.30 g·kg-1,全氮含量为0.68 g·kg-1,全磷含量为1.42 g·kg-1,速效钾含量为152.6 mg·kg-1,0~20 cm土壤容重为1.57 g·cm-3,平均土壤含水量0.4%,海拔为1 378 m,年均降雨量为113.2 mm,年均蒸发量为2 604.3 mm,年均气温为6.9℃,7月平均温为24.9℃,1月平均气温为—9.9℃,日最高气温为35.9℃,日最低气温为—25.1℃[28-29]。
1.2 试验材料的培养及选取
2011年在白刺和红砂的成熟季节,沿自然环境降水梯度在我国甘肃酒泉(JQ)、张掖(ZY)、武威(WW)和兰州(LZ)天然分布区分别选取每个种源中生长良好、无病虫害的多个优势单株,采集种子,并进行标记装袋;2012年1月在武威市凉州区羊下坝镇甘肃武威市良种繁育中心苗圃温室进行穴盘育苗,基质成分为沙土10%~20%,砂壤土60%~80%,腐熟羊粪10%~20%,播种后进行日常的苗木管理。2013年4月,土壤解冻后,进行移栽,建立白刺和红砂种源试验地。移栽时采用随机区组试验设计,株行距0.5 m×1.5 m。移栽后随即灌水,保证苗木的成活率。待苗木成活后,适时除草,按日常管理方法对苗木进行管理[29]。
2020年在苗木生长旺季,以同质园内定植的4个种源地的白刺和红砂作为研究对象(4个种源地的地理位置与环境因子见表1[30]),分别选取每个种源中生长健壮、长势良好、无病虫害的10个成年植株,剪取每个植株直立茎中部的叶片放入打湿的信封袋中,并且装入封口塑料袋内封口,立即带回实验室用于叶片功能指标测定。
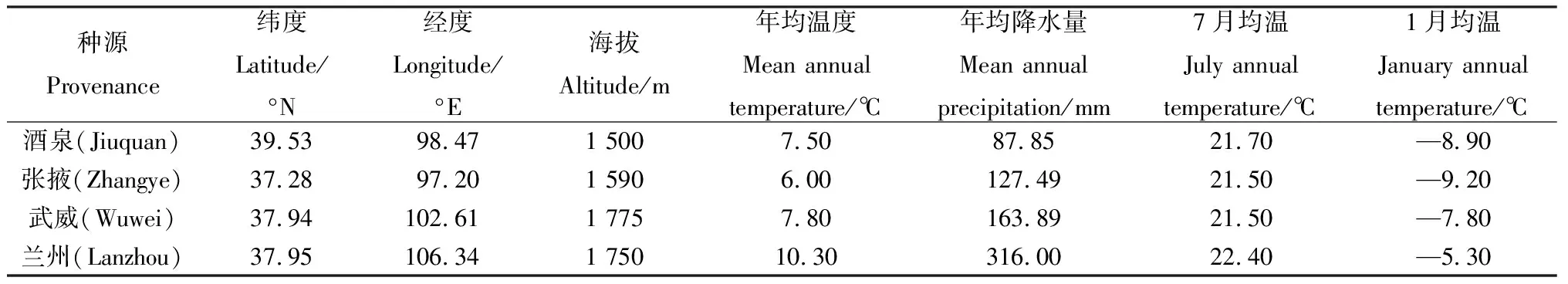
表1 白刺和红砂4个种源地的地理位置与环境因子Table 1 Geographical location and environmental factors of the four provenances of N. tangutorum and R. soongarica
1.3 测定方法
1.3.1叶片形态性状的测定 将采集叶片上的杂质清理掉,称量叶鲜重,再将叶片用根系分析软件(Win-RhIZO 2008a)测量叶片表面积、体积等形态特征。测量完成之后,将叶片置于75℃烘箱烘至恒重,称量干重并记录。每种源各有3次重复。白刺和红砂叶片的肉质化程度、含水量、比叶面积和组织密度的计算公式如下:

(1)
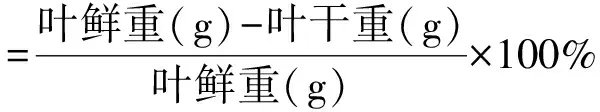
(2)

(3)

(4)
1.3.2叶片碳(C)、氮(N)和磷(P)含量的测定 将以上测定完的植物样品用碾槽碾碎,装进密封袋用于进行叶片C,N,P含量的测定。C含量测定采用重铬酸钾氧化加热法;N含量测定采用凯氏定氮法;P含量测定采用钒钼黄比色法测定[31]。
1.4 气象数据来源
本研究选择了对植物叶功能性状具有影响的环境因子。气象数据以酒泉、张掖、武威和兰州4个种源地的数据为主(http://www.weather.com.cn/)。通过对数据的整理,计算出每个种源附近各站点的经度(Longitude,Lng)、纬度(Latitude,Lat)、海拔(Altitude,ALT)、年均温度(Mean annual temperature,MAT)、年均降水量(Mean annual precipitation,MAP)、1月均温(January annual temperature,JaT)、7月均温(July annual temperature,JuT)等数据,作为本次研究的气象数据。
1.5 数据处理
本试验所有数据统计分析和相关作图均是使用Excel 2016,SPSS 24.0(America),Origin 2018(America)和Canoco5.0(America)软件完成。采用单因素方差分析法分析同一物种在不同种源间叶片各性状差异显著性,并用Duncan检验法和最小显著差异法(LSD)进行多重比较;用Pearson相关系数检验白刺和红砂各叶功能性状之间的相关性。采用冗余分析(RDA)法和逐步线性回归法分析白刺和红砂叶功能性状对不同种源地环境因子的响应关系;在RDA分析之前,先对叶功能性状进行去趋势对应分析(DCA),确定白刺和红砂是采用以线性模型排序的冗余分析(RDA)还是以单峰模型排序的典型对应分析(CCA)。若最大排序轴长小于3,则适合线性模型排序分析的RDA方法;若最大排序轴大于4,则适合单峰模型排序的CCA方法;若最大排序轴长在3~4之间,则两种方法都可以。本研究中最大排序轴长为0.1,小于3,故采用RDA方法分析,且将种源地环境因子作为解释变量(空心箭头),将叶功能性状作为响应变量(实心箭头)。文中数据均为平均值±标准误差。
2 结果与分析
2.1 白刺和红砂叶片形态性状的种源间差异
由图1可知,白刺叶片肉质化程度在JQ种源与其他地理种源差异显著(P<0.05),且在JQ种源最小(3.99),与ZY种源、WW种源和LZ种源相比,JQ种源白刺叶片肉质化程度减小8.62%,14.36%和10.95%;含水量在JQ种源与其他地理种源差异显著(P<0.05),且在JQ种源最小(74.88),与ZY种源、WW种源和LZ种源相比,JQ种源白刺叶片含水量减小2.56%,4.47%和3.22%;组织密度在种源间差异显著(P<0.05),且在ZY种源最大(0.08 g·cm-3),在WW种源最小(0.06 g·cm-3),与WW种源相比,ZY种源白刺叶片组织密度显著增加了30.79%。
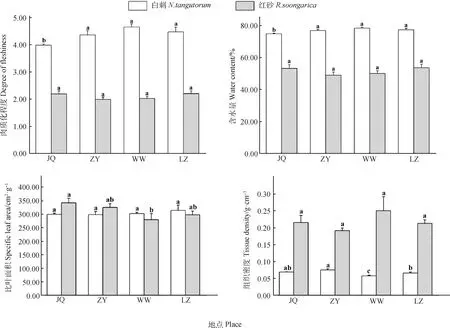
图1 不同地理种源白刺和红砂叶片形态结构Fig.1 Leaf morphological structure of N. tangutorum and R. soongarica from different geographic provenances注:JQ-酒泉、ZY-张掖、WW-武威、LZ-兰州;不同小写字母表示同一物种在不同种源间差异显著(P<0.05)。下同Note:JQ-Jiuquan,ZY-Zhangye,WW-Wuwei,LZ-Lanzhou;different lowercase letters indicate significant difference between different provenances under the same species at the 0.05 level. The same as below
红砂叶片的比叶面积在JQ和WW种源间差异显著(P<0.05),与WW种源相比,JQ种源的红砂叶片的比叶面积增加了22.24%;肉质化程度、含水量和组织密度在各个种源均无显著差异。
2.2 白刺和红砂叶片C,N,P含量及其计量比的种源间差异
由图2可知,白刺叶片N含量在种源间差异显著(P<0.05),在JQ种源最大,为3.79 g·kg-1,在LZ种源最小,为3.31 g·kg-1;叶片N∶P在JQ种源与其他种源间均差异显著(P<0.05),且在JQ种源比值最大,为10.37,与ZY种源、WW种源和LZ种源相比,JQ种源叶片N∶P增加了47.95%,24.72%和63.72%。

图2 不同地理种源白刺和红砂叶片C,N,P含量及其计量比Fig.2 Contents of C,N,and P in leaves of N. tangutorum and R. soongarica and their stoichiometric ratio from different geographic provenances
红砂叶片P含量在JQ和WW种源间差异显著(P<0.05),在JQ种源最大,为0.37 g·kg-1,在WW种源最小,为0.28 g·kg-1,与WW种源相比,JQ种源红砂叶片P含量增加了30.40%。
二者的C含量、C∶N和C∶P在各个种源均无显著差异。
2.3 白刺和红砂叶片功能性状间的相关性
通过Pearson相关性分析得出(表2),白刺叶片的组织密度与肉质化程度、含水量和比叶面积呈极显著负相关关系(P<0.01);N含量与比叶面积呈显著负相关关系(P<0.05),C∶P与C含量呈极显著正相关关系(P<0.01);N∶P与N含量呈极显著正相关关系(P<0.01),与C∶N呈显著负相关关系(P<0.05)。通过Pearson相关性分析可以看出(表3),红砂叶片的组织密度与比叶面积呈极显著负相关关系(P<0.01);C∶N与C含量呈极显著正相关关系(P<0.01),与N含量呈显著负相关关系(P<0.05);C∶P与C含量呈极显著正相关关系(P<0.01),与P含量呈显著负相关关系(P<0.05),与C∶N呈极显著正相关关系(P<0.01);N∶P与N含量呈显著正相关关系(P<0.05)。

表2 不同地理种源白刺叶片功能性状间的相关性Table 2 Correlation among leaf functional traits of N. tangutorum from different geographic provenances

表3 不同地理种源红砂叶片功能性状间的相关性Table 3 Correlation among leaf functional traits of R. soongarica from different geographic provenances
2.4 不同地理种源白刺和红砂叶片功能性状与种源地环境因子的关系
2.4.1叶片功能性状与种源地环境因子的冗余分析 通过RDA分析(表4),第1,2轴的白刺叶功能性状与环境的相关系数分别为0.9163,0.9213,特征值分别为0.3265,0.1574,共解释了48.40%的叶功能性状和92.37%的叶功能性状与种源地环境的关系,表明前两个排序轴能够较好地反映白刺叶功能性状与种源地环境因子的关系。蒙特卡罗检验第1排序轴和所有排序轴均达到极显著水平(P<0.01),表明结果具有统计学意义。由图3可知,海拔与肉质化程度、含水量呈正相关关系,与组织密度呈负相关关系;纬度与N∶P呈正相关关系,与P含量呈负相关关系;年均降水量与C含量、N含量、C∶P呈负相关关系。RDA分析(表5)表明,第1,2轴的特征值分别为0.804 6,0.006 9,共解释了81.16%的红砂叶功能性状和99.69%的叶功能性状与种源地环境的关系,表明前两个排序轴能够较好地反映红砂叶功能性状与种源地环境因子的关系。蒙特卡罗检验第1排序轴和所有排序轴均达到极显著水平(P<0.01),表明结果具有统计学意义。由图4可知,海拔、年降水量和年均温度与第1排序轴呈正相关关系,且相关性依次为海拔>年降水量>年均温度;海拔与红砂比叶面积、P含量呈负相关关系。
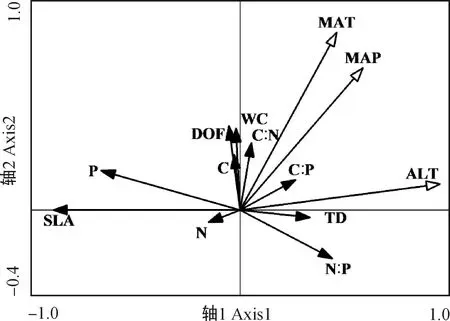
图4 红砂叶功能性状与种源地环境因子的RDA排序Fig.4 Redundancy analysis ordination of leaf functional traits of R. soongarica and geographic environmental factors

表4 白刺叶性状与环境因子的RDA排序Table 4 Redundancy analysis ordination of leaf traits and environmental factors of N.tangutorum
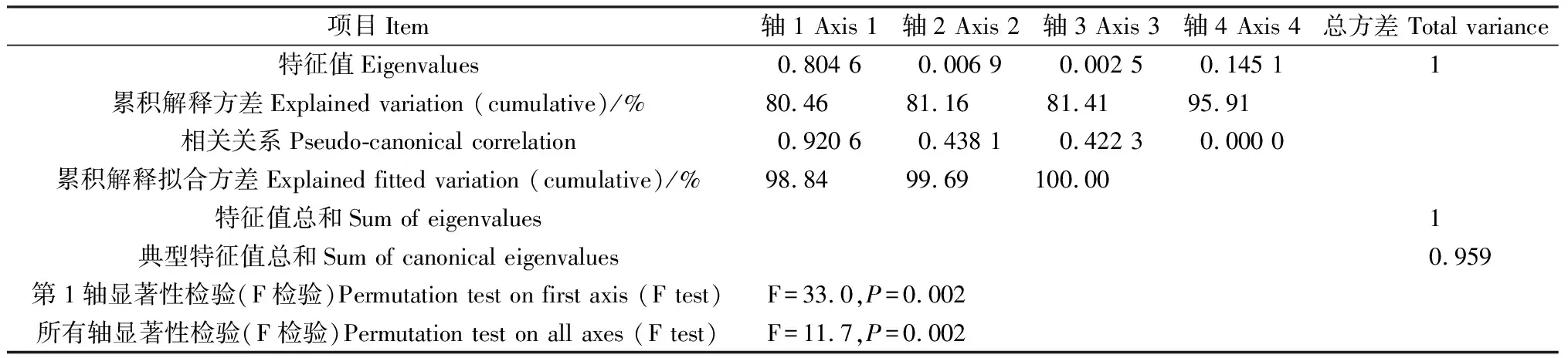
表5 红砂叶性状与环境因子的RDA排序Table 5 Redundancy analysis ordination of leaf traits and environmental factors of R. soongarica

图3 白刺叶功能性状与种源地环境因子的RDA排序Fig.3 Redundancy analysis ordination of leaf functional traits of N. tangutorum and geographic environmental factors注:经度-Lng,纬度-Lat,海拔-ALT,年均温度-MAT,年均降水量-MAP,下同Note:Longitude-Lng,Latitude-Lat,Altitude-ALT,Annual average temperature-MAT,Annual average precipitation-MAP,the same as below
2.4.2叶功能性状与种源地环境因子的逐步线性回归分析 由表6可知,白刺叶片肉质化程度、含水量与海拔呈极显著正相关关系(P<0.01);叶组织密度与海拔、纬度均呈极显著负相关关系(P<0.01),且海拔的标准化回归系数的绝对值(|—0.697|)大于纬度的标准化回归系数的绝对值(|—0.441|),表明影响叶组织密度的主要环境因子是海拔,其次是纬度;叶片N含量与年均降水量呈显著负相关关系(P<0.05),与1月均温呈显著正相关关系(P<0.05),且年均降水量的标准化回归系数的绝对值(|—2.704|)大于1月均温的标准化回归系数(2.126),表明影响叶片N含量的主要环境因子是年均降水量,其次是1月均温;叶片P含量与纬度呈显著负相关关系(P<0.05),叶片N∶P与纬度呈显著正相关关系(P<0.05),与7月均温呈显著负相关关系(P<0.05),且纬度的标准化回归系数(0.708)大于7月均温的标准化回归系数的绝对值(|-0.441|),这表明影响叶片N∶P的主要环境因子是纬度,其次是7月均温。因此,白刺叶片肉质化程度、含水量、组织密度主要受海拔的影响,叶片N含量主要受年均降水量的影响,叶片P含量、N∶P主要受纬度的影响。而红砂叶片的比叶面积和P含量主要受海拔影响。
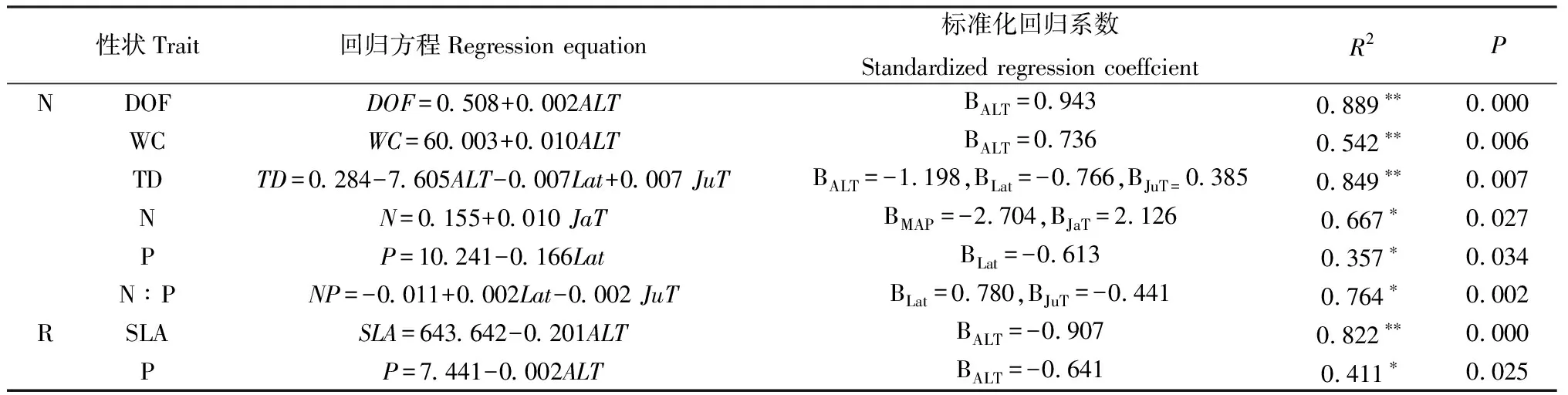
表6 白刺和红砂叶功能性状与环境因子的逐步回归分析Table 6 Stepwise regression analysis of leaf functional traits and environmental factors of N. tangutorum and R. soongarica
3 讨论
3.1 不同地理种源白刺和红砂叶片形态性状的变异特征
植物在长期适应种源环境下,必然会产生生理生态特性的分化,形成不同的地理种源,导致同一叶功能性状在不同种源间差异显著[32]。大量研究表明,植物长期受种源地环境影响,导致其叶功能性状产生遗传分化[33-35]。本研究结果也显示,不同地理种源白刺在同质园环境中叶片肉质化程度、含水量、组织密度表现出显著性差异(图1),表明白刺受种源地环境的影响,其叶功能性状差异可能与遗传分化有关。有研究表明,在野外环境和同质园环境中,植物由于受种源环境的影响,产生了遗传稳定性[17]。杨彪生等[36]研究发现,野外环境下白刺叶片肉化程度、含水量和组织密度均呈现显著性差异,这一野外研究结果与本研究中同质园实验的研究结果一致,说明白刺叶功能性状通过对种源环境的适应可能形成了遗传稳定性。杨彪生等[36]对野外环境中红砂叶片肉质化程度、含水量和组织密度的研究发现这些指标均呈显著性差异。然而,本研究中同质园内不同地理种源红砂叶片的肉质化程度、含水量、组织密度均无显著性差异(图1),说明红砂在同质园环境中不受种源环境的影响。
比叶面积可以反映植物对资源的获取能力,且也能够反映出植物对不同环境的适应性特征,是研究植物功能性状的重要指标之一[37-38]。有研究认为,植物在资源丰富的环境中生长比叶面积较大,在资源匮乏的环境中生长往往比叶面积较小[39]。在本研究中,同质园中红砂的比叶面积(311.26 cm2·g-1)高于野外环境中红砂的比叶面积(277.32 cm2·g-1),这可能是同质园环境有较充足的水分、光照等条件,与野外环境相比,同质园中比叶面积利用光合的能力较强,能够快速获取所需资源。
3.2 不同地理种源白刺和红砂叶片C,N,P含量化学计量及其计量比的变异特征
叶片C,N,P含量对异质环境的适应是表型分化和遗传分化共同的结果,并在长期的进化过程中形成不同的生理生态和养分利用特征,若在同质园环境中,这种生理生态的分化依然存在,则认为植物为了适应环境产生了遗传分化[17]。有研究表明,同质园试验植物叶片的C,N,P含量及其计量比在种源间表现出显著性差异,说明植物叶片中化学计量特征在对种源地环境条件的长期进化适应过程中产生了遗传分化[40-41];本研究发现白刺叶片N含量,N∶P在不同地理种源间表现出的结果与上述结果相同(图2),说明白刺受种源环境的影响可能使其叶片化学元素产生了遗传分化。不同种源翅荚木(Zeniainsignis)叶片C含量、N含量及C∶N存在差异显著,说明其主要叶片养分性状可能主要受长期的种源地环境影响产生了遗传分化[32],这与本文研究中红砂叶片C含量、N含量及C∶N无显著差异的结论相悖(图2),这可能是因为翅荚木研究地属于中亚热带海洋性季风气候,雨水充沛,气候温暖,而本文研究地属于干旱性大陆气候,干旱少雨,两个研究地的立地条件和气候环境截然不同,所以对其叶功能性状的影响程度也不同。
C,N,P含量化学计量比对植物营养元素的平衡起限制性作用,并已经广泛运用于植物养分循环和限制的研究[42-43]。N∶P常被看作植物在生长过程中获取环境养分的限制性因素[1,44]。有研究表明,植物生长发育受N和P限制的N∶P比值为14和16,即当N∶P<14时,植物生长发育主要受N限制;当N∶P>16时,植物生长发育主要受P限制;当14 植物叶片在长期适应地理环境的过程中,通过内部不同功能间的协调,形成一系列适应环境的叶功能性状间的不同组合,并非叶片性状孤立发挥作用[47-48]。通常认为,植物叶片的比叶面积与组织密度呈负相关的关系,即比叶面积增大时,组织密度会变小。比叶面积和组织密度作为植物资源分配利用的重要指标,均能够反映植物对环境的适应策略(水分利用策略)[49-51]。本研究发现,白刺和红砂的比叶面积与组织密度呈极显著负相关关系(表2、表3),出现这种现象可能与植物地处干旱少雨的环境密切有关,即当植物处于干旱环境时,植物为了适应干旱环境,从周围环境中不断汲取养分,使得比叶面积增大,但此过程中植物对光照需求较大,而对叶片中储存的养分和水分的需求较小,故养分资源的利用效率相对较低,因此植物叶片相对较薄,叶组织密度相对较小。因此,白刺和红砂叶片通过比叶面积和组织密度的平衡形成对不同水分环境的适应策略[47,52]。有研究表明,作为植物生长中主要的营养元素,N和P具有一定的相互促进作用,N∶P可以反映N和P两种营养元素供给状况的相对有效性[53]。白刺和红砂叶片N含量与N∶P呈显著正相关的关系(表2、表3),这是由于植物对养分的利用受到了环境和自身需求的影响,植物通过调节自身叶片营养元素及化学计量比来适应环境中养分的供给情况[41]。叶片C∶N和C∶P表示植物吸收N,P元素所能同化碳的能力,是植物养分利用效率的重要指标[54]。本研究中,红砂叶片C∶N与C∶P呈极显著正相关关系(表3),说明红砂叶片具有较高的碳同化能力。 广泛分布的植物对其环境适应的范围较广,可以通过表型可塑性、局域适应等来面对环境变化,其选择的策略将决定植物应对气候变化的能力[55],一般来说,叶性状若与种源地环境因子显著相关,则可认为发生局域适应[56]。结合RDA分析和逐步线性回归法分析,白刺叶片N含量与年均降水量有相关性(图3,表6),这可能是因为本研究4个种源地干旱少雨,除LZ种源外,其余种源年均降水量均不足200 mm,而且水分是植物生长发育过程中所必需的因子,与其他因子相比,植物功能性状对水分的响应更为显著[33],因此说明白刺叶片N含量主要受水分的影响。一般来说,海拔对植物功能性状的影响机理主要是随着海拔的升高,植物生长发育逐渐受到温度和水分的限制,因而植物对环境的适应性会发生显著变化[57]。本研究结果显示,白刺肉质化程度和含水量与海拔呈极显著正相关关系,组织密度与海拔呈极显著负相关关系(图3,表6),并且海拔对植物比叶面积、组织密度、干物质含量等影响显著的结论已得到证实[57-58],因此白刺肉质化程度、含水量和组织密度随海拔的变化受温度和水分的限制。海拔对红砂叶片比叶面积和P含量的影响呈显著负相关关系(图4,表6),并且随着海拔的上升,比叶面积和P含量趋向于降低(图1,图2),这可能是由于本研究所采样的4个种源地均属于高海拔地区,使得植物生长所需要的温度和水分受到了影响,植物为了适应高海拔的恶劣环境,其物质代谢发生改变,导致植物获取资源的能力和对环境的适应能力发生了变化[59]。因不同种源地植物叶功能性状的变异受到多个地理环境因子的共同影响,本文只选取了纬度、经度、海拔、年均降水量、年均温度等环境因子对叶片功能性状进行研究,没有涉及到各种源地物种所在地的土壤养分、坡向、相对湿度、积温等环境因子。因此,在今后的研究中,还需在这方面作进一步研究。 通过研究同质园中各地理种源白刺和红砂叶功能性状的种间差异,发现白刺在适应环境的过程中可能产生了遗传分化,且受环境影响可能形成了遗传稳定性,而红砂在同质园中不受种源环境的影响;两种植物叶片生长发育主要受氮素限制;受环境的影响,两种植物叶片内部功能性状的平衡变化形成了对不同环境的适应策略;分析白刺和红砂叶功能性状与种源环境因子的关系,发现海拔是影响两种植物的主导环境因子,故两种植物生长受温度和水分的限制。3.3 白刺和红砂叶功能性状间的关系
3.4 种源地环境对白刺和红砂叶功能性状的影响
4 结论
