花叶鹅掌柴不同颜色叶片光合特性
2022-03-25吴霞毛行简凡莉莉陈凌艳何天友荣俊冬郑郁善
吴霞 毛行简 凡莉莉 陈凌艳 何天友 荣俊冬 郑郁善





摘 要:以多年生花葉鹅掌柴(cv. )成年植株为试验材料,选择同一植株上的3种颜色叶片,分别为全绿叶(whole green leaf, WGL)、花叶(variegated leaf, VL)和黄叶(whole yellow leaf, WYL),测定其光合色素含量、光响应曲线、气体交换参数和叶绿素荧光参数,研究花叶鹅掌柴3种不同颜色叶片的光合特性,为合理化种植和植物培育提供理论指导和参考依据。结果表明:3种颜色叶片的叶绿素a()、叶绿素b()、总叶绿素()和类胡萝卜素()含量差异极显著(<0.01),均表现为全绿叶>花叶>黄叶,且3种颜色叶片的均为0.556;全绿叶的净光合速率()随光强增大而上升的幅度大于花叶,黄叶的净光合速率()始终为负值;全绿叶和花叶的最大净光合速率()、光饱和点()和暗呼吸速率()存在显著差异(<0.05),二者的光补偿点()差异不显著(>0.05),均表现为全绿叶>花叶;花叶的初始斜率(α)高于全绿叶,且存在显著差异(<0.05);气孔导度()、蒸腾速率()和水分利用率()表现为全绿叶>花叶>黄叶,胞间二氧化碳浓度()则相反,表现为黄叶>花叶>全绿叶;初始荧光()和实际量子产量(Φ)值与叶绿素含量呈正相关,表现为全绿叶>花叶>黄叶,3种叶片值小于稳定值0.832,和值随着叶绿素含量的上升呈先上升后下降,表现为花叶>全绿叶>黄叶。花叶鹅掌柴3种颜色叶片均具有较强的耐阴能力,其中全绿叶和花叶还能适应一定强光环境,其光合作用的产物是供黄叶生长的重要来源;叶绿素含量较高的全绿叶片具有更高的光合作用效率;花叶对低光强利用能力和对强光下的自我保护能力较强,具有较高的栽培与培育的价值。
关键词:花叶鹅掌柴;花叶植物;光合特性;叶绿素荧光特性中图分类号:S687 文献标识码:A
Photosynthetic Characteristics of Different Color Leaves of ‘Variegata’
WU Xia, MAO Xingjian, FAN Lili, CHEN Lingyan, HE Tianyou, RONG Jundong, ZHENG Yushan
1. College of Landscape Architecture, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China; 2. College of Forestry, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China
Three color leaves of the adult plant of cv. on the same plant including whole green (WGL), variegated leaf (VL) and whole yellow leaf (WYL) were used as the test material, to determine its photosynthetic pigment content, light response curve, gas exchange parameters and chlorophyll fluorescence parameters and to study the photosynthetic characteristics of three different color leaves of cv. so as to provide theoretical guidance for rational planting and plant cultivation and reference basis. The results showed that the contents of chlorophyll a (), chlorophyll b (), total chlorophyll () and carotenoid () of the three colors of leaves were extremely different (<0.01), and they were WGL > VL > WYL, and the of the three colors of leaves were 0.556; the net photosynthetic rate () of whole green leaf increases with the increase of light intensity greater than that of Variegated leaf and whole yellow leaf () that was always a negative value; the maximum net photosynthetic rate (), light saturation point () and dark respiration rate (Rd) of whole green leaf and variegated leaf are significantly different (<0.05), and the light compensation points of the two () the difference was not significant (>0.05), which showed that whole green leaf> variegated leaf; the initial slope (α) of variegated leaf was higher than that of whole green leaf, and there was a significant difference (<0.05); stomatal conductance (), transpiration rate () and water use efficiency () are shown as whole green leaf > variegated leaf > whole yellow leaf, while the intercellular carbon dioxide concentration () was the opposite, and showing whole yellow leaf > variegated leaf > whole green leaf; initial fluorescence () and Actual quantum yield () value was positively correlated with chlorophyll content, which was represented by whole green leaf > variegated leaf > whole yellow leaf. The values of the three leaves were less than the stable value 0.832. The and values increased firstly and then later with the increase of chlorophyll content. The performance is variegated leaf > whole green leaf > whole yellow leaf. Whole green leaf with higher chlorophyll content have higher photosynthesis efficiency, and the growth and development of whole yellow leaf without chlorophyll will be restricted and die; variegated leaf have stronger adaptability and regulation ability than whole green leaf, and can be used at lower chlorophyll concentrations. Under the circumstances, improving photosynthetic efficiency through self-regulation has higher cultivation and cultivation value.
cv. ; variegated plants; photosynthetic characteristics; chlorophyll fluorescence characteristics
10.3969/j.issn.1000-2561.2022.03.014
花葉鹅掌柴(cv. )是一种花叶类观赏灌木,为五加科(Araliaceae)鹅掌柴属()鹅掌柴()的一个变种,多年生常绿灌木,叶革质,较厚,富有光泽,叶片上间有黄白色斑纹,具有很高的观赏和应用价值,具有一定的耐阴能力,在园林植物造景时,常被栽植在乔木下、建筑旁或是植物群下层等阴协地,这对于提高单位面积绿地绿化的生态效益和植物景观的复层结构设计、丰富植物景观具有重要意义。但是目前对其研究主要集中于栽培和被寄生方面,具体光合特性研究还未见报道,栽植仍旧凭借旧的经验。
叶片叶绿素生物合成受阻会是导致花叶和黄叶产生的重要原因,色素含量及种类会影响植物叶片颜色,叶绿素含量越多,叶片绿色越深,另外植物叶片是植物进行光合作用,生成有机物的重要场所,叶片叶绿素含量越多,叶片光合能力越强,黄叶片由于叶绿素合成受阻,导致净光合速率为负值。光合作用是植物生长发育的生理基础,光合作用与植物的生理生态特性存在着紧密联系。对银丝竹、花叶假连翘、竹柏的光合特性研究表明,全绿叶利用弱光能力较强,花叶利用强光能力较强,且在较强光下能刺激花叶的产生,并保持花叶特性。
鉴于此,选择以花叶鹅掌柴作为研究对象,对其全绿叶、花叶和黄叶3种叶色表型叶片的光合特性关系进行研究,整体把握花叶鹅掌柴3种叶片的光合特点,了解其对光照的需求特性,为科学种植和植物配置提供理论参考和配置建议,充分发挥其观赏价值和生态价值,丰富园林植物景观。
材料与方法
材料
花叶鹅掌柴为福建省福州市福建农林大学中华园内种植5年以上的成年植株,作为荫生地被观赏植物,栽植于一棵成年印度橡胶树下,具有较好遮阴效果四周无遮挡,已经充分适应栽培地环境。福州的气候属于海洋性亚热带季风气候,温暖湿润,年相对湿度约77%,年平均降水量为900~2100 mm,年平均日照辐射量为419~ 5 016 MJ/m。待测叶选择同一健康植株第二轮成熟叶片,根据叶色组成,将待测叶分为3种颜色叶片(图1):全绿叶(whole green leaf, WGL)、花叶(variegated leaves, VL)和黄叶(whole yellow leaf, WYL)。
方法
1.2.1 光合色素含量测定 运用丙酮混合法提取光合色素,将丙酮混合液的比例改为丙酮∶无水乙醇∶纯水=4.5∶4.5∶1。采集同一花叶鹅掌柴植株上的3种不同颜色新鲜叶片若干,重复3次,求平均值,根据公式(1)~(3)分别计算出3种光合色素含量:叶绿素a()、叶绿素b()及类胡萝卜素()。
=(12.72×‒2.59×)×/(1000×)
(1)
=22.88×‒4.67××/(1000×) (2)
=(×/‒2.05× ‒114.8×)/245
(3)
1.2.2 光合光响应测定 选择连续两天晴天,在2021年3月17—18日上午9:00—11:30,下午14:00—17:30进行测定,温度为15~20℃。选择同一花叶鹅掌柴植株中上部成熟且健康的3种叶色叶片,LI-6 400-02 XT BLED便携式光合作用测定系统(LI-COR 6400,USA)红蓝光源提供光照强度设定16个不同:0、20、40、60、80、100、200、400、600、800、1000、1200、1400、1600、1800、2000 μmol/(m·s),二氧化碳(CO)流速为400 μmol/mol,记录净光合速率[μmol/(m·s)]、气孔导度[μmol/(m·s)]、胞间二氧化碳浓度[μmol/(m·s)]和蒸腾速率[mmol/(m·s)]数据,计算植物水分利用瞬时效率[mmol/(m·s)],公式为=/。使用直角双曲线修正模型,计算初始斜率α、最大净光合速率[μmol/(m·s)]、光饱和点[μmol/(m·s)]、光补偿点[μmol/(m·s)]和暗呼吸速率[μmol/(m·s)]和校准参数R。
1.2.3 葉绿素荧光参数测定 使用便携式脉冲调制叶绿素荧光仪OPTI-Sciences OS5P(OS5P,USA),选择同一健康花叶鹅掌柴植株的中上部,
第二轮成熟叶9片,全绿叶、花叶和黄叶各3片,做好标记。先将叶片进行30 min暗适应,再打开测量初始荧光(),饱和脉冲光进行照射,测量最大荧光()和的最大光化学效率()。光下叶片光合稳态时的实际光化学效率Φ的测定需要在自然条件下打开一个饱和脉冲。荧光淬灭参数的测定时,设置Kinetic模式,记录光化学猝灭系数()和非光化学猝灭系数(),重复测定3次。
数据处理
用Microsoft Excel 2007和SPSS 20软件对试验数据进行处理和分析,运用直角双曲线修正模型进行光响应曲线拟合和特征参数计算,使用Microsoft Excel 2007软件制作图表。
果与分析
不同颜色叶片光合色素分析
3种颜色叶片的光合色素含量差异显著,全绿叶片的、、和含量显著高于花叶和黄叶(<0.05),分别是花叶的1.590倍、1.587倍、1.592倍、1.522倍,是黄叶的14.667倍、14.875倍、1.592倍、8.750倍。花叶的、、和含量显著高于黄叶,分别是黄叶的9.222倍、9.375倍、9.320倍、5.750倍。3种颜色叶片的)含量均为0.556(表1)。
不同颜色叶片光响应曲线及光合特征参数分析
在0~100 μmol/(m·s)全绿叶和花叶的净光合速率迅速上升,由负值升为正值;200~ 1000 μmol/(m·s)时,全绿叶和花叶的净光合速率持续增加至光饱和点(),增加速度逐渐减小,全绿叶的净光合速率始终大于花叶;1000~ 2000 μmol/(m·s),全绿叶和花叶的净光合速率由
光饱和点()又逐渐小幅度下降,其中花叶下降的速度大于全绿叶;黄叶片在光照强度()为0~2000 μmol/(m·s)时始终为负值,呼吸速率大于光合速率(图2)。
通过对花叶鹅掌柴3种颜色叶片的光合响应曲线的拟合结果表明,全绿叶和花叶的拟合效果较好,校准系数分别为0.999和0.997,黄叶的拟合效果较差,校准系数仅为0.225;全绿叶片的最大净光合速率、光饱和点、光补偿点和暗呼吸速率分别为14.717、1392.318、13.685、1.269 μmol/(m·s)均高于花叶,且、和与花叶存在显著差异(<0.05),与花叶差异不显著(>0.05)。花叶的初始斜率α显著高于全绿叶(<0.05);黄叶未能测出光响应曲线,因此不予分析(表2)。
不同颜色叶片气体交换参数分析
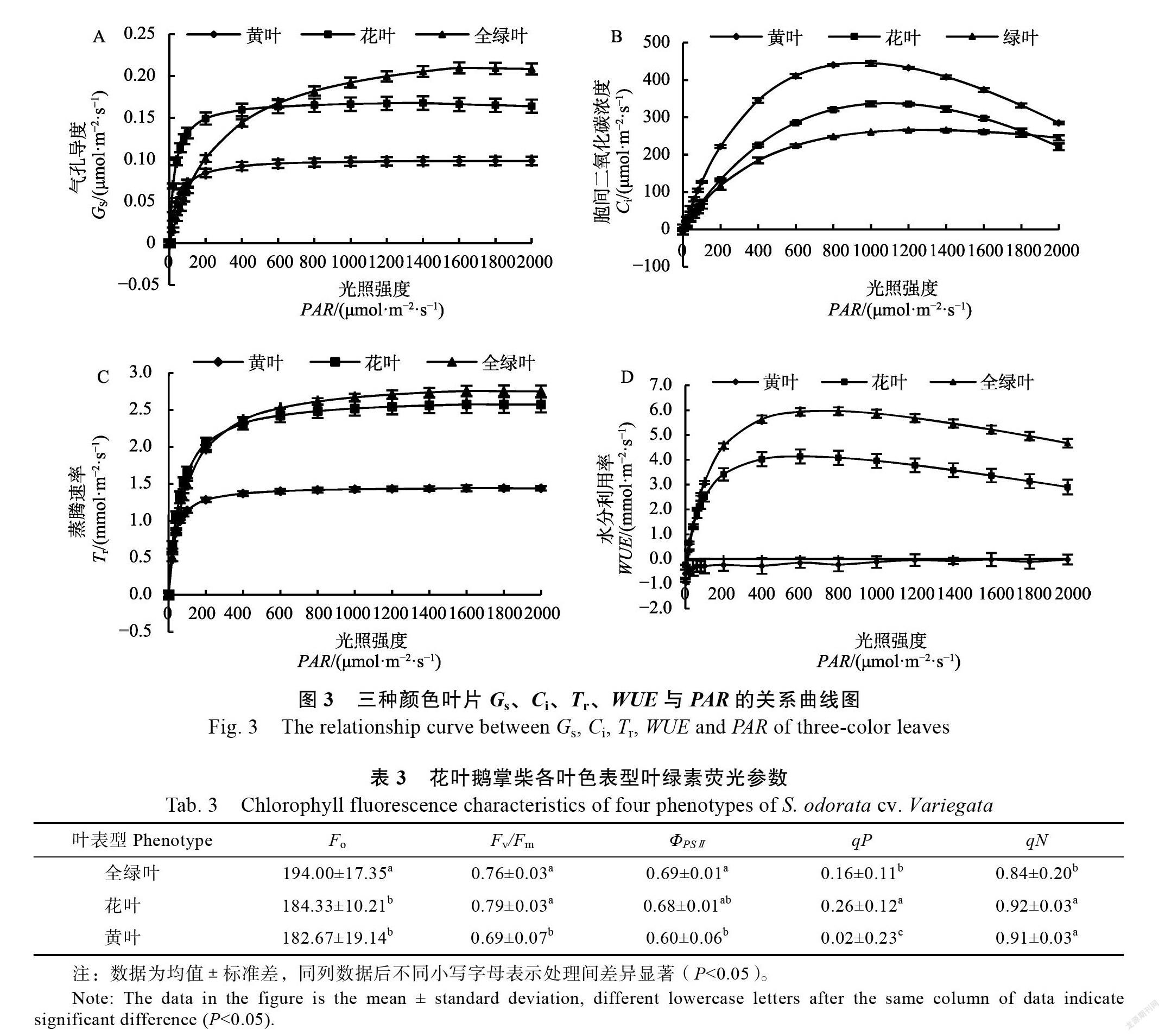
3种叶片的变化趋势一致,随着的增大而增大,至最大值后开始缓慢下降,0~ 200 μmol/(m·s)时,3种颜色叶片的值表现为花叶>黄叶>全绿叶,200~600 μmol/(m·s)时,表现为花叶>全绿叶>黄叶,600~2000 μmol/(m·s)时,表现为全绿叶>花叶>黄叶(图3A)。随着的增大呈现先上升后下降的趋势,光强为1000 μmol/(m·s)時,黄叶和花叶达到最大值,后缓慢下降,全绿叶在光强为1500 μmol/(m·s)时达到最大值,1500~2000 μmol/(m·s)时保持稳定值,并在1900时与花叶的曲线相交(图3B);<300 μmol/(m·s)时,3种颜色叶片的表现为黄叶>全绿叶>黄叶,在=300 μmol/(m·s)时,全绿叶和花叶的曲线相交,在> 300 μmol/(m·s)时,3种颜色叶片的则表现为全绿叶>花叶>黄叶(图3C)。3种颜色叶片的差异显著,表现为全绿叶>花叶,其变化趋势与3种叶片的相一致,黄叶的始终为负值(图3D)。
花叶鹅掌柴不同颜色叶片的叶绿素荧光参数
3种颜色叶片的值大小表现为全绿叶>花叶>黄叶,但全绿叶的显著高于花叶和黄叶;全绿叶和花叶的差异不显著,但黄叶显著低于全绿叶和花叶;从3种颜色叶片的Φ值来看,全绿叶的实际光合效率最大,但花叶与全绿叶的差异不显著,全绿叶与黄叶存在显著差异。3种颜色叶片的均表现为花叶>全绿叶>黄叶,且存在显著差异;花叶和黄叶的差异不显著,但全绿叶的显著低于花叶和黄叶(表3)。
叶绿素与光合生理参数相关性分析
叶绿素是植物进行光合作用时主要的光合色素,其含量的高低直接影响了植物的光合作用效率。花叶鹅掌柴3种不同颜色叶片与其、、及呈显著正相关;与、r及呈显著正相关,相关性不显著;与和呈显著正相关;C与、、、及皆呈负相关,相关系数较低,分别为‒0.552、‒0.655、‒0.508、‒0.303、‒0.532,且显著性不高(表4)。
叶绿素与叶绿素荧光参数相关性分析
与的相关系数很高,相关系数为0.980,与和Φ的相关系数较低,分别为0.601和0.867;与和呈负相关,相关系数分别为‒0.944和‒0.947;与、和呈正相关,相关系数分别为0.984、0.919、0.988,与Φ呈负相关,相关系数为‒0.655;与和呈正相关,相关系数分别为0.975和1.0,与Φ呈负相关,相关系数为‒0.508;Φ与其他均为负相关,与呈正相关,相关系数为0.867(表5)。
讨论
阴协地是一种由自然或非自然因素,如建筑、乔木等,对低矮植物遮挡阳光所致的生长环境,通常阴生环境的光照强度与正常光照环境的光照强度差异较大。在遮荫环境生长的植物,为提高对光能的吸收能力,通常会提高的含量,导致(/)比值减小。因此(/)比值可以判断植物的耐阴能力,一般以3.2为分界线,小于3.2时为耐阴植物,比值越小,植物耐阴性越强,反之则为阳生植物。花叶鹅掌柴3种颜色叶片的(/)均为0.556,小于3.2,因此其具有强耐阴能力。叶绿素通过吸收和传递光能用于植物进行光合作用,其含量越高,植物对光能的吸收能力越强,光合作用速率越大,光合作用速率高低体现着植物生长发育状态,是评价植物生长状态的重要参数。花叶鹅掌柴的全绿叶的叶绿素含量最大,随着的增加而上升的速度较花叶和黄叶快,黄叶由于叶绿素含量太低,叶片吸收和传递光能效率最小,光合作用速率在为0~2000 μmol/(m·s)时始终为负值,因此黄叶生长发育所需的养分只能通过植物自身对同一植株上其他叶片光合作用产物的调控来供给,这与毛行简等的研究结果相同。另外和含量影响着植物叶色,叶绿素积累,叶片绿色加深,叶绿素减少,叶片绿色变浅或失绿。花叶鹅掌柴3种颜色叶片的和含量表现为全绿叶>花叶>黄叶,因此可以判断花叶鹅掌柴白叶和花叶的产生是因为叶绿素含量的减少而导致。
光响应参数表征植物的光合特性,主要包括光合效率、光能利用率及光抑制水平等,不同类型叶片对光照环境的反应不同,对光照强度适应能力较小的叶片在光照增强时易受到光抑制,和代表了植物的需光特性和需光量,是植物叶片光合作用时对光照条件的两种要求,是评估植物对光照强度适应性的特征参数,表征的是植物对弱光的适应能力,表征的是植物对强光的适应能力,有研究表明< 46 μmol/(m·s),<460 μmol/(m·s)时为耐阴植物,花叶鹅掌柴的全绿叶和花叶的为13.685和9.711 μmol/(m·s),均小于46 μmol/(m·s),全绿叶和花叶的为1 392.318和1 021.249 μmol/(m·s),均大于460 μmol/(m·s),且花叶的和显著低于全绿叶,这说明花叶鹅掌柴全绿叶和花叶的光强适应范围较广,都具有强耐阴性,这与全绿叶和花叶的(/)比值反应现象一致。另外花叶鹅掌柴的全绿叶较高,花叶的较低,这说明了花叶对弱光的适应能力更强。初始斜率α表现了植物在低光照强度下的净光合速率的增加速率和对光能的利用率,表征植物在时的净光合速率。花叶的α高于全绿叶,低于全綠叶,表示花叶对弱光的利用率高于全绿叶,且能在较低的时达到最大的净光合速率,这与陈凌艳等的研究结果相同,但其绿底白纹叶片初始斜率为0.08,仅略高于全绿叶(0.07),这可能是不同植物叶片对低光强利用能力具有一定的差异。综上所述,花叶和全绿叶对阴生环境和强光环境都具有一定的适应能力,花叶对弱光的利用能力更强,全绿叶对强光的利用能力更强,这与银丝竹、花叶假连翘的研究结论相反,这可能是因为花叶鹅掌柴属于耐阴植物,而银丝竹和花叶假连翘属于中性或阳生植物,也可能与试验材料所生长的环境有关,有待进一步研究。
气孔是外界气体进入植物叶片,参与光合作用的重要通道,是植物气孔开放程度的特征参数,影响着植物叶片的水分蒸腾、光合作用、呼吸作用等一系列过程。花叶鹅掌柴3种颜色叶片的随着光强的增大先迅速上升后缓慢下降,其中在光强较低时,上升的幅度最大,这说明花叶鹅掌柴在低光强下光合作用会消耗大量的胞间二氧化碳,为保持光合作用的持续进行,叶片会迅速打开气孔,促进叶片与大气之间的气体交换,加快了水分蒸腾速率,同时也提高了水分利用率。
叶绿素荧光参数主要表达的是植物在光合作用中的能量传递和转化,揭示植物进行光合作用时对光的利用、散失和分派等方面的变化 ,特征参数主要包括原初光能转化效率、初始荧光、的实际量子产量Φ、光化学淬灭值、非光化学猝灭值。其中表征的是对光能的利用能力,反映了植物的潜在最大光合能力,该值下降是光抑制最明显的特征,非逆境情况下,比值稳定在0.832±0.0004;是处于完全开放时的反应中心的荧光强度,反映了天线色素受激发后的电子密度,与叶绿素浓度呈正相关;Φ是光照状态下的实际量子产量,反映了植物目前的实际光合效率;表征的是用于光化学电子传递的天线色素利用的光能分量,反映了植物光合活性的高低;表征的是天线色素将吸收过量的光能以热能形式消耗掉的光能部分,体现着植物的光保护能力。从3种颜色叶片的值来看,均在0.832以下,说明此时的花叶鹅掌柴生长环境的光照强度高于其最适的光照强度,花叶鹅掌柴始终是处于光抑制生长状态。3种颜色叶片的变化趋势同总叶绿素含量一致,说明在完全开放时全绿叶的初始荧光强度大于花叶,这与陈凌艳等和赖金莉等结果一致;但花叶具有较强自我调节能力,在叶绿素浓度较低的情况下,花叶(较高值)会提高捕获的光能中转化为光化学电子传递的效率,从而最大限度提高花叶的光合作用,而且在较高光照强度下,花叶(较高值)也会将多余的光能以热能的形式消耗掉,保护自身免遭光灼伤。
综上所述,花叶鹅掌柴不仅耐阴能力较强,同时也能适应一定的强光,在园林栽培过程中应用较为灵活,既可在阳光直射的地方与乔木搭配,作为地被观赏植物,也可在建筑旁边的阴生环境栽植。花叶鹅掌柴的花叶不仅在低光强下具有较高的光能利用能力,而且在较高光强环境下,具有较强的自我保护和调节能力,拥有较强的适应能力,因此在培育时可以提高花叶的数量;而全绿叶片本身叶绿素含量较高,在适合的光照环境下,对光能的利用效率最好,是植物生长所需有机物的重要来源;黄叶能够提高植物的观赏特性,但其无法生产足够的有机物,需要依赖全绿叶和花叶的提供才能正常生长,因此需控制每株上的黄叶数量,确保在保持植物观赏性的前提下,植株也能健康生长。
参考文献
- 张力群. 花叶鹅掌柴无土栽培技术探讨[J]. 中国科技信息, 2012(21): 67.ZHANG L Q. The discussion on soilless culture of cv. [J]. China Science and Technology Information, 2012(21): 67. (in Chinese)
- 许 军, 黄晓红, 杨振德, 周 光, 黄超群. 花叶鹅掌柴对日本菟丝子寄生的生理响应[J]. 中国农学通报, 2010, 26(23): 192-195.XU J, HUANG X H, YANG Z D, ZHOU G, HUANG C Q. Physiological response ofcv. to cuscuta japonica parasitization[J]. Chinese Agricultural Science Bulletin, 2010, 26(23): 192-195. (in Chinese)
- 陈凌艳, 何丽婷, 赖金莉, 何舒婷, 吴玉香, 郑郁善. 银丝竹不同叶色叶绿素合成及叶结构差异[J]. 森林与环境学报, 2017, 37 (4): 385-391.CHEN L Y, HE L T, LAI J L, HE S T, WU Y X, ZHENG Y S, The variation of chlorophyll biosynthesis and the structure in different color leaves of ‘Silverstripe’[J]. Journal of Forest and Environment, 2017, 37 (4): 385-391. (in Chinese)
- YANG Y X, CHEN X X, XU B, LI Y X, MA Y H, WANG G D. Phenotype and transcriptome analysis reveals chloroplast development and pigment biosynthesis together influenced the leaf color formation in mutants of ‘Sonate’[J]. Frontiers in Plant Science, 2015, 6:139.
- 王啸晨, 张 磊, 岳祥华, 高志民. 菲黄竹叶片色素含量、结构与颜色之间关系初探[J]. 世界竹藤通讯, 2012, 10(6): 5-9.WANG X C, ZHANG L, YUE X H, GAO Z M, A preliminary study of relationship of leaf pigments content and structure with leaf color of [J]. World Bamboo and Rattan, 2012, 10(6): 5-9. (in Chinese)
- TANG Y H, FANG Z W, LIU M, ZHAO D Q, TAO J. Color characteristics, pigment accumulation and biosynthetic analyses of leaf color variation in herbaceous peony ( Pall.)[J]. 3 Biotech, 2020, 10: 76.
- 陈凌艳, 谢德金, 荣俊冬, 赖金莉, 林雪玲, 郑郁善. 光合色素含量差异对花叶唐竹不同叶色表型光合特性的影响[J]. 林业科学, 2019, 55(12): 21-31.CHEN L Y, XIE D J, RONG J D, LAI J L, LIN X L, ZHENG Y S. Effects of photosynthetic pigment content on photosynthetic characteristics of different leaf color phenotypes of f. [J]. Scientia Silvae Sinicae, 2019, 55(12): 21-31. (in Chinese)
- 赖金莉, 李欣欣, 陈凌艳, 荣俊冬, 郑郁善. 银丝竹3种颜色叶片光合特性研究[J]. 生态环境学报, 2018, 27 (2): 255-261.LAI J L, LI X X, CHEN L Y, RONG J D, ZHENG Y S. Photosynthetic characteristics of three different colors of leaves of cv. [J]. Ecology and Environmental Sciences, 2018, 27(2): 255-261. (in Chinese)
- 毛行简, 郑风英, 任 可, 陈凌艳, 何天友, 郑郁善. 花叶假连翘3种叶色表型叶片的光合特性[J]. 森林与环境学报, 2021, 41(3): 225-233MAO X J, ZHENG F Y, REN K, CHEN L Y, HE T Y, ZHENG Y S. Photosynthetic characteristics of three leaf color phenotypes of ‘Variegata’[J]. Journal of Forest and Environment, 2021, 41(3): 225-233. (in Chinese)
- 闾邱杰, 曹景怡. 竹柏2种颜色叶片的光合特性研究[J]. 热带亚热带植物学报, 2020, 28(2): 177-184.LV Q J, CAO J Y. Photosynthetic characteristics of two color leaves of [J]. Journal of Tropical and Subtropical Botany, 2020, 28(2): 177-184. (in Chinese)
- 张聪颖, 方炎明, 姬红利, 马成涛. 遮荫处理对红叶石楠和洒金桃叶珊瑚光合特性的影响[J]. 应用生态学报, 2011, 22(7): 1743-1749.ZHANG C Y, FANG Y M, JI H L, MA C T. Effects of shading on photosynthesis characteristics of × and var. [J]. Chinese Journal of Applied Ecology, 2011, 22(7): 1743-1749. (in Chinese)
- 张 婷. 6种常春藤品种的光合生理特性比较[J]. 西北林学院学报, 2019, 34(6): 62-67.ZHANG T. Comparison of leaf photosynthetic and physiological characteristics of 6 cultivars[J]. Journal of Northwest Forestry University, 2019, 34(6): 62-67. (in Chinese)
- 王海珍, 韩 路, 徐雅丽, 牛建龙. 胡杨异形叶光合作用对光强与CO浓度的响应[J]. 植物生态学报, 2014, 38(10): 1099-1109.WANG H Z, HAN L, XU Y L, NIU J L. Photosynthetic responses of the heteromorphic leaves in to light intensity and CO concentration[J]. Chinese Journal of Plant Ecology, 2014, 38(10): 1099-1109. (in Chinese)
- 罗光宇, 陈 超, 李月灵, 金则新. 光照强度对濒危植物长序榆光合特性的影响[J]. 生态学杂志, 2021, 40(4): 980-988.LUO G Y, CHEN C, LI Y L JIN Z X. Effects of light intensity on the photosynthetic characteristics of [J]. Chinese Journal of Ecology, 2021, 40(4): 980-988. (in Chinese)
- 段 娜, 贾玉奎, 郝玉光, 徐 军, 高君亮, 陈海玲. 干旱胁迫对欧李叶绿素荧光特性的影响[J]. 西北林学院学报, 2018, 33(6): 10-14.DUAN N, JIA Y K, HAO Y G, XUN J, GAO J L, CHEN H L. Effects of drought stress on chlorophyll fluorescence characteristics of [J]. Journal of Northwest Forestry University, 2018, 33(6): 10-14. (in Chinese)
- BJRKMAN O, DEMMIG B. Photon yield of O evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins[J]. Planta, 1987, 170(4): 489-504.
