肉制品中杂环胺类化合物形成及控制措施的研究进展
2022-03-24王惠汀孙学颖王丹郭进靳烨赵丽华
王惠汀,孙学颖,王丹,郭进,靳烨,赵丽华
(内蒙古农业大学食品科学与工程学院,内蒙古 呼和浩特 010018)
杂环胺类化合物(heterocyclic aromatic amines,HAAs)是富含蛋白质的食物在加工过程中产生的一类致癌、致突变性的物质。20世纪70年代,人们首次在烤牛肉和烤鱼的表面发现了杂环胺的存在,并发现长期摄入含HAAs的食物会增加患结肠癌、乳腺癌、肝癌等癌症的风险[1]。目前,人们已经发现近30种HAAs[2]。研究表明,一些富含蛋白质的肉制品(如腌腊肉制品、油炸肉制品、酱卤肉制品等)经过高温加热后都会检出杂环胺[3]。随着生活水平的提高,人们对于食物的要求不再只是色、香、味,而更侧重于食物的安全品质。
杂环胺作为日常饮食中常见的一类食源性致癌物,广泛存在于高温加热的肉制品中,因此有必要对其进行研究。本文主要对杂环胺进行综述,通过分析其形成途径,系统地阐述了杂环胺形成的影响因素、控制措施及其代谢的研究进展。
1 杂环胺的分类
1.1 根据化学结构分类
HAAs根据化学结构可分为氨基-咪唑-氮杂芳烃(amino-imidazo-azaarenes,AIAs)与氨基-咔啉类(aminocarbolines,ACs)两大类。根据结构差异分类,AIAs包括喹啉类、喹喔啉类、吡啶类和呋喃吡啶类。咔啉类可分为 α-咔琳类、β-咔啉类、γ-咔啉类和 ζ-咔啉类[4]。常见的杂环胺类化合物及其分类见表1。

表1 常见杂环胺类化合物及其分类Table 1 Common heterocyclic amines and their classification
1.2 根据化学性质分类
HAAs根据化学性质可分为极性杂环胺和非极性杂环胺。AIAs均含有咪唑环,与IQ化学性质类似,故又称为IQ型杂环胺,即极性杂环胺。氨基-咔啉类称为非IQ型杂环胺,即非极性杂环胺。
1.3 根据生成温度分类
根据Millar[5]的研究发现,300℃是加热蛋白质食物(如肉类和鱼类)中形成不同类别杂环胺的关键临界温度。根据生成温度可将HAAs分成两大类:一类是氨基咪唑氮杂芳烃(即IQ型杂环胺),是氨基酸、肌酸(肌酰)、糖在100℃~300℃的条件下通过美拉德反应生成,其结构上的氨基可耐受2 mmol/L亚硝酸钠的重氮处理,在体内可转化成N-羟基化合物,又称为“热反应杂环胺”,具有致癌、致突变性;另一类是咔啉类杂环胺(非IQ型杂环胺),是蛋白质或氨基酸在300℃以上的温度下生成,其结构上的氨基不能耐受2 mmol/L亚硝酸钠的重氮处理,处理时氨基脱落转变成C-羟基而失去致癌、致突变性,因此其致癌、致突变性比热反应杂环胺弱,又称为“热解型杂环胺”[6]。
2 杂环胺的形成机制
2.1 热反应型HAAs的形成机制
2.1.1 IQ和IQx型杂环胺的形成
富含蛋白质的食品在加热过程中会发生美拉德反应,很容易生成HAAs。己糖和氨基酸通过美拉德反应中的Strecker降解,生成的吡啶和吡嗪分别是IQ和IQx型杂环胺的前体物质,吡啶或吡嗪衍生物与肌酸加热过程中形成的肌酐通过醇醛缩合反应,生成了IQ或 IQx型 HAAs[7]。
2.1.2 吡啶类杂环胺的形成
目前,吡啶类杂环胺中PhIP的形成机制已经得到了证实。Felton等[8]首次从油炸牛肉中分离出PhIP。目前,PhIP的形成途径已基本清晰,主要分为3步:首先通过Strecker降解将苯丙氨酸分解生成苯乙醛,此步骤中没有肌酐的参与,但体系中存在的活性羰基化合物会促进反应;然后体系中的肌酐与苯乙醛通过醇醛缩合反应脱水后形成缩合物,在此过程中,苯乙醛会降解生成甲醛,肌酐会降解生成氨;最后,脱水产物与氨和甲醛反应形成PhIP[9]。
2.2 热解型HAAs的形成机制
目前,热解型杂环胺的已知形成途径主要有两种,一是由游离氨基酸(色氨酸、谷氨酸、苯丙氨酸等)在300℃以上的高温热解形成;二是由大豆球蛋白和酪蛋白热解形成。两者都是通过热解形成各种脱羧基、氨基产物和活性自由基进而诱导形成杂环胺[10]。然而也有特殊的即色氨酸和葡萄糖在小于100℃的低温下干热形成Harman和Norharman[11]。
3 杂环胺的代谢
通常杂环胺的生成量是以纳克每克为单位进行计量的,但长期摄入会对人体健康造成巨大威胁,因此研究杂环胺在人体内的消化吸收及代谢尤为重要。杂环胺的代谢分为解毒和活化,首先发生的是Ⅰ相代谢,进入体内的杂环胺经过细胞色素P450(cytochrome P450,CYP450)酶催化发生羟基化反应,随后发生Ⅱ相代谢,即葡萄糖醛酸转移酶(glucuronyl transferase,UGTs)和磺基转移酶(sulfotransferase,SULTs)催化反应产生的芳香环羟基化产物和部分环外氨基羟基化产物,和葡萄糖醛酸或磺酸形成解毒物质,最后经粪便或尿液排出体外;另一部分的环外氨基羟基化产物通过氮乙酰转移酶(N-acetyltransferase,NATs)或磺基转移酶发生催化酯化,形成的活化产物会进一步异裂生成亲电子的芳基氮离子中间物,易与大分子物质(如蛋白质、DNA等)进一步反应形成加合物,最终产生致癌活性[12]。以PhIP为例,其代谢途径见图1。
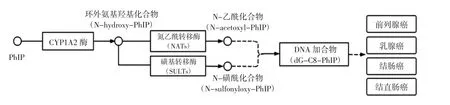
图1 PhIP代谢途径Fig.1 The metabolic pathway of PhIP
PhIP在进入体内后,经细胞色素P450 CYP1A2酶代谢后生成环外氨基羟基化产物,部分环外氨基羟基化产物与葡萄糖醛酸或磺酸形成解毒物质排出体外,另一部分代谢途径如图1所示,环外氨基羟基化产物通过氮乙酰转移酶(NATs)或磺基转移酶(SULTs)发生催化酯化,生成N-乙酰化合物或N-磺酰化合物,与DNA进一步生成DNA加合物即dG-C8-PhIP,最终引起前列腺癌、乳腺癌、结肠癌、结直肠癌等。
4 杂环胺的影响因素
4.1 烹饪方式
在日常饮食中,常用烹饪方式有水煮、煎炸、油炸、酱卤等,加工方式不同对肉制品中杂环胺生成的影响也不同。研究表明,传统的烹饪方式升温是通过介质传递来实现,先是给食物表层加热,然后传热到中心,使食物彻底熟透需要较长的时间和温度[9]。Sinha等[13]研究了煎、烤、炸、煮等加热方式以及熟化程度对PhIP浓度和致癌性的影响,发现水煮牛肉、鸡肉、培根中的PhIP含量均低于其他加热方式。Fan等[14]采用微波水煮和电加热水煮两种加热方式对牛肉糜进行了长达2 h的处理,发现在加热过程中,微波牛肉糜中的PhIP生成量始终显著低于传导加热。杨洪生等[15]研究了不同的加工方式对草鱼鱼糜制品中杂环胺生成影响的研究,结果表明在油炸、烘烤、水煮3种加工方式中,油炸方式下杂环胺生成的种类和数量最多,其次为烘烤方式,而水煮方式最少。因此,在日常饮食中,选择合适的烹饪方式对食物进行加工,可以从源头有效得减少HAAs的生成。
4.2 加工温度和时间
对杂环胺生成影响程度最大的两个因素是温度和时间。Gibis等[16]研究发现,在200℃~220℃下加工的培根中,MeIQx和PhIP的含量高于在150℃~170℃下煎炸的水平。Ahmad等[17]研究发现,9种HAAs的浓度随着温度从150℃升高到350℃逐渐升高。Buła等[18]研究发现220℃与180℃烧烤相比,烤猪肉中4,8-二聚体和7,8-二聚体的含量分别增加2倍和9倍。此外,烹调时间越长,HAAs含量也越高。在大多数情况下,应考虑热处理温度和时间的组合,以确定HAAs的形成。多项研究表明,食品中HAAs的生成量与其加工温度和时间呈显著正相关[19-20]。
4.3 前体物质
目前,肌酸、氨基酸、葡萄糖、含氮碱以及相应的核苷被认为是生成HAAs的前体物质。任何前体物质的缺失都会影响HAAs的生成。Szterk[21]研究了游离氨基酸、核苷、含氮碱、蛋白质和葡萄糖对烤牛肉中HAAs水平的影响,发现HAAs生成的影响与氨基酸总量有很强的相关性。自由氮碱基和相应的核苷形式与HAAs形成的关联性比自由氨基酸的关联性强。此外,这些前体物在不同动物品种的肉制品中的存在和含量也不同。廖国周等[22]研究前体物含量对杂环胺形成的影响,发现前体物的含量在不同物种间和同一物种内都存在着较大的差异,且在同一物理条件下加工的肉制品中形成的HAAs也存在较大的差异。
4.4 其它因素
脂肪作为肉制品中的主要营养成分之一,对于HAAs的生成也起到了一定的作用。首先,脂肪含量较高的肉类传热能力更高效,达到产生HAAs的热温度所需的时间更短[2]。其次,肉类中高脂肪含量与导致HAAs形成的前体物(如氨基酸、肌酸和葡萄糖)的较低相对含量有关。综上所述,如果同时考虑这两个方面,脂肪是否有利于促进HAAs的形成仍然是个未知数。郭海涛等[23]研究了脂肪添加量及原料肉形态对烤羊肉饼中杂环胺形成的影响,结果发现IQ型杂环胺的生成量与脂肪含量具有显著的相关性,即脂肪含量的增加会显著提高极性杂环胺的生成量,而非IQ型杂环胺的含量与脂肪含量无显著相关性,且极性杂环胺的生成量与肉饼质量呈显著负相关,而非极性杂环胺下降趋势不明显;通过对原料肉进行绞碎处理可使极性和非极性杂环胺含量升高,其原因可能是肉在绞碎过程中肌酸等前体物质流出至表面,使更多的前体物质暴露在高温下,形成较多的HAAs。因此,优化脂肪含量,是尽可能减少食物中HAAs产生的一条有效的途径。水作为前体物质迁移的关键介质,防止肉制品中水分的流失有助于减少HAAs的形成。杨调调[24]发现,生香肠经过干燥、烘烤和蒸煮处理后,HAAs含量逐渐增加,而干燥阶段HAAs生成量最多。肉制品中的糖在热加工过程中会发生焦糖化反应和美拉德反应,α-二羰基化合物作为反应的重要中间体具有高度的化学活性,易进一步发生反应[25]。肉制品中的杂环胺通过美拉德等相关反应生成,所以美拉德反应的中间体α-二羰基化合物与肉制品中糖对杂环胺形成的影响密切相关。
5 杂环胺的抑制措施
一些研究者已经证明了HAAs的诱变活性,而进一步的研究也表明HAAs对包括癌症在内的人类疾病的发生和发展有很大的贡献[26-27]。国际癌症研究机构已确认9种HAAs,包括2-氨基-9H-吡啶 [2,3-b]吲哚、2-氨基-3-甲基-9H-吡啶[2,3-b]吲哚、2-氨基-1-甲基-6-苯基咪唑[4,5-b]吡啶和 2-氨基-3,4-二甲基咪唑[4,5-f]喹啉可能具有致癌作用;2-氨基-3-甲基咪唑并[4,5-f]喹啉也可能是致癌物,并建议饮食中减少接触这些有害物质。此外,美国国家毒理学计划(National Toxicology Program,NTP) 研究报告认为,IQ、MeIQ和PhIP等一些HAAs可能是人类致癌物[28]。目前,已被证实杂环胺的摄入量与人类患食道癌、胃癌、结肠癌等癌症的风险呈正相关。近年来,患阿尔兹海默症的人数逐年上升,Syeda等[29]研究表明,人类患阿尔兹海默症可能与PhIP的暴露相关。因此,有必要控制杂环胺的形成并对其机制进行研究。
5.1 控制杂环胺的生成
杂环胺是一类具有致癌性和致突变性的多环芳香族化合物,主要通过与其环外游离氨基与DNA反应生成加合物而产生,常见于经过高温烹饪富含蛋白质的肉制品中[30]。目前,我国尚未建立杂环胺的相关限量标准,但由于其致癌性和致突变性且普遍存在于高温肉制品中,因此其形成规律及控制措施成为了人们的研究热点[31]。杂环胺的生成受到许多因素的影响,如加工方式、温度和时间等。因此可以从以上几个方面来控制杂环胺的产生。
5.1.1 选择合适的加工方式
肉制品在传统的加工方式中如煎炸、油炸、烘烤等都会产生HAAs,因此选择合适的加工方式是控制肉制品中HAAs生成的一条有效措施。郭海涛[32]采用不同的烹饪方式对羊肉饼进行加工,结果发现,羊肉饼中总杂环胺含量由高到低依次为酱卤、烘烤、油炸、煎炸。王震等[33]研究了反复卤煮对鸭胸肉和卤汤中杂环胺及其前体物的影响,结果表明,鸭胸肉在卤煮过程中,随着卤汤使用次数的增多,卤汤和鸭肉杂环胺的前体物质即游离氨基酸、葡萄糖、肌酸等含量显著升高,并且杂环胺生成总量与卤煮次数呈显著正相关。李利洁[34]比较了3种加热过程即传导密封加热、微波回流加热和传导回流加热中PhIP的生成情况,微波回流加热产生的PhIP量较传导回流加热少,但加合物产生量约是传导回流加热的10倍;传导密封加热中密封条件引发的压强增大会显著促进PhIP及加合物的生成。Felton等[35]探究降低油炸牛肉饼过程中HAAs的方法时发现,在对牛肉饼进行加工前先对其进行微波预处理,会降低体系中杂环胺的前体物的含量,从而降低杂环胺的生成。Suleman等[36]比较分析了木炭烧烤、红外烧烤和过热蒸汽烧烤对羊肉饼中杂环胺含量的影响,结果表明,红外烧烤法和过热蒸汽烧烤法可以降低羊肉饼中HAAs的含量,而炭烤是非极性HAAs(如Harman和Norharman)形成的主要原因,且在极性HAAs中DMIP和PhIP的含量也相对较高。多项研究表明,随着肉制品加热时间的延长和加热温度的升高,其HAAs的含量会逐渐增加,影响肉类产生杂环胺的因素强弱顺序为时间>温度>肉的种类[37]。王园等[38]研究油炸条件对鱼肉中杂环胺生成量影响时发现,鱼肉在150℃油炸,检出Norharman和Harman 2种杂环胺,总量为0.23 ng/g;在210℃时检出4种,总量为9.85 ng/g。210℃油炸20 min的样品中检出6种杂环胺,杂环胺总量达27.09 ng/g。由此可知,HAAs含量和种类会随着油炸温度的升高及时间的延长逐渐增加。因此,通过适当降低加热温度、缩短加热时间以及对原料肉进行微波预处理可降低加工肉制品中产生的杂环胺。
5.1.2 添加外源物质
自由基反应是HAAs形成的重要途径之一,因此可通过添加抗氧化物质来阻止HAAs的形成。原因是抗氧化物质可以干扰HAAs形成反应的不同阶段。由于合成的抗氧化剂具有致癌潜力,近年来人们多集中于研究天然的抗氧化剂,因此,富含植物或香辛料等天然抗氧化剂备受人们关注[39]。BAO等[40]研究了不同浓度黑胡椒粉对罗非鱼鱼片中6种芳香杂环胺含量的影响。结果表明,添加黑胡椒和不添加黑胡椒的罗非鱼鱼片中 HAAs含量分别为 6.4 ng/g~24.72 ng/g和10.43 ng/g~33.19 ng/g。1.0% 黑胡椒粉对 PhIP和MeIQx的生成抑制率为100%,由此得知黑胡椒可降低高温加工肉制品中的杂环胺。罗舟[41]研究竹叶提取物和9种黄酮类化合物对化学模型中PhIP和MeIQx的抑制作用,结果发现化学模型中PhIP和MeIQx的形成明显受到竹叶提取物抑制,且呈显著负相关,即杂环胺的形成量随竹叶提取物的添加而减少。这可能是因为竹叶提取物的抗氧化作用清除了反应过程中的自由基,阻碍了反应的进行,从而显著抑制了杂环胺的产生。不同种类黄酮类化合物因具有不同的化学结构而形成了不同的抗氧化能力,从而对两种杂环胺表现出了不同的抑制作用,且在相同添加量的情况下,表现出了不同的抑制效果;而对于同一种黄酮类化合物,随着黄酮类物质添加量的增加,PhIP的抑制率显著升高,说明黄酮对杂环胺的抑制可能与其清除自由基的能力有关。壳聚糖是一种由甲壳素衍生的天然聚合物,具有较强的抗氧化能力。Mirsadeghi等[42]探究了不同类型的壳聚糖对鱼肉中杂环胺的影响,结果发现壳聚糖对杂环胺表现出了一定的抑制作用。水作为杂环胺前体物质迁移的关键介质,在抑制杂环胺方面起着很重要的作用,因此,人们在肉制品加工过程中常加入一些保水性物质(如大豆分离蛋白、马铃薯淀粉等)来抑制杂环胺的生成。Wang等[43]研究发现在煎炸牛肉中加入一定量的大豆浓缩蛋白,会使杂环胺的诱变活性下降,此外,还发现大豆汉堡中的HAAs与炸牛肉汉堡中的HAAs相比具有更低的诱变活性。综上所述,可以通过添加抗氧化物质如香辛料、黄酮类化合物和保水性物质等外源物质来抑制杂环胺的生成。
5.2 控制杂环胺的代谢
虽然有大量的方法可以控制杂环胺的形成,减少从食物中的摄入,但仍然会有部分杂环胺进入体内,因此,对于杂环胺代谢的调控也是必不可少的。研究表明,人体内只有0.5%~6.0%的HAAs没有被代谢转化。有研究表明部分乳酸菌可以吸附未代谢的HAAs,使其失活不能再进行代谢,最后排出体外,多数人认为其吸附机理是由于乳酸菌中细胞壁上的肽聚糖与HAAs发生阳离子交换[44]。Bolognani等[45]研究产乳酸的生物体培养物与膳食致癌物的结合能力,并研究乳酸菌对苯并[a]芘、杂环胺(IQ、MeIQ、MelQx、PhIP、Trp-P-2)的结合能力,结果发现,长双歧杆菌和嗜酸乳杆菌对苯并[a]芘和Trp-P-2的结合最有效,而对MelQx、MelQ、PhIP和IQ的结合能力较差,且乳酸菌对杂环胺的吸附能力与培养过程中的pH值条件密切相关,且pH值对吸附能力的影响在长链链球菌中比嗜酸链球菌更明显。Nowak等[46]通过体外研究鼠李糖乳杆菌0900、鼠李糖乳杆菌0908和干酪乳杆菌0919对IQ和PhIP的影响,结果发现3株乳酸菌对IQ和PhIP具有吸附作用,从而阻止结肠癌等癌症的发生。有研究者对比了常规大鼠、无菌大鼠和有菌群的大鼠的单细胞凝胶电泳,结果表明,肠道微生物的存在显著增强了IQ对结肠和肝细胞DNA损伤的诱导作用。此外,研究还发现,在饲料中添加乳酸菌可减弱IQ对结肠癌的诱导作用。这些研究表明,乳制品中的肠道菌群和乳酸杆菌能干预HAAs给人体带来的健康风险[47]。Stidl等[48]研究了12株乳酸菌对肉制品中较丰富的杂环胺(AαC、PhIP、IQ、MeIQx、DiMeIQx)的解毒能力,结果发现,乳酸菌对每种胺的解毒能力存在较大差异,解毒能力从大到小依次为AαC>DiMeIQx>MeIQx>IQ>PhIP。Thyagaraja等[49]研究了一种非乳发酵食品中的乳酸菌在pH 6.98的环境下对Trp-P-1、Trp-P-2、Glu-P-1等诱变剂的结合潜力。在此pH值条件下,所有的乳酸菌菌株都表现出对Trp-P-1和Trp-P-2的良好结合活性,而与Glu-P-1和IQ的结合活性较差。Sreekumar等[50]研究加氏乳杆菌对杂环胺的结合受体时发现,杂环胺与细胞壁的结合与碳水化合物含量呈高度相关,且经过不同化学(蒸馏水、酸、碱等)和酶(溶菌酶、蛋白酶K、胰蛋白酶)处理后的菌对杂环胺的吸附能力均有不同程度地提升。因此,乳酸菌吸附杂环胺的机制还需要进一步的研究。Orrhage等[51]研究8株人肠道或乳酸菌对高蛋白食品烹饪过程中形成的诱变杂环胺的结合能力。采用高效液相色谱法分析菌株与诱变剂Trp-P-2、PhIP、IQ和MeIQx的结合能力,结果表明试验菌株的结合能力只有微小差异,但诱变化合物的结合效率明显不同。Trp-P-2几乎完全结合,而且这种结合是不可逆的。IQ和MeIQx的结合稍差,且pH值对结合效果有重要影响,结果表明,熟食中常见的诱变化合物在体外可与正常肠道菌群结合。然而,在选择合适的摄食菌株时,还需要考虑其他因素,如菌株的安全性和存活率。一些致病细菌,如梭状芽孢杆菌、类杆菌、真杆菌和大肠杆菌所产生的酶可以使体内HAAs活化产生致癌物质,研究发现乳酸菌可以降低这些酶的活性[42]。研究发现,一些物质也可以使解毒酶(UGTs或SULTs)增加,如啤酒中的啤酒花,其衍生物质黄腐酚是一类丁烯基黄酮类化合物,可能会使二磷酸尿苷(uridine diphosphate,UDP)-葡萄糖醛酸转移酶增多,从而能有效地降低食源性致癌性杂环胺(HAAs)2-氨基-3-甲基咪唑[4,5-f]喹啉(IQ)的遗传毒性和潜在致癌性[52]。
此外,膳食纤维也可以抑制结直肠癌的发生,其作用机制主要有两种,一种是直接作用;另一种是间接作用,即膳食纤维可被结肠细菌酶和发酵产物降解而产生有作用的纤维。Ferguson等[53]使用大鼠研究小麦麦麸对杂环胺的影响,结果表明,麸皮对杂环胺的作用主要通过直接作用即直接与杂环胺结合从而减少其毒性。此外,曹春雨[54]研究辣椒辣素对仓鼠肝微粒体中杂环胺代谢及细胞色素P4501A2活性的影响,结果发现辣椒辣素可在体外明显抑制HCAs的代谢。研究发现,茶叶及其主要成分具有抗氧化、影响致癌物代谢酶或者直接与活性致癌物结合等功效,因此茶叶及其主要活性成分茶多酚具有预防癌症的作用。大鼠谷胱甘肽巯基转移酶(glutathione S-transferase,GST)可催化N-乙酰氧基-PhIP与谷胱甘肽反应,从而抑制PhIP-DNA加合物的形成。然而林东昕等[55]研究绿茶对大鼠体内HAA-DNA加合物生成的抑制情况,结果发现,绿茶确实可以显著抑制大鼠体内PhIP形成加合物,但和CYP及GST等代谢酶无关。分析可能是在体内的茶叶成分与PhIP的活性代谢产物反应,从而阻止了PhIP进一步活化形成致癌加合物。因此,茶叶及其主要成分茶多酚对杂环胺的抑制机理仍需进一步探究。综上所述,可以添加乳酸菌来吸附进入体内而未代谢的杂环胺,使其失活不能再继续进行代谢,最后随尿液或粪便排出体外,同时添加一些可使解毒酶增加的物质阻止杂环胺的遗传毒性。
6 总结与展望
本文从肉制品中杂环胺的分类、形成机制、影响因素以及控制措施方面进行阐述,以期为肉制品加工中杂环胺的监控作出贡献。烹饪方式、加工时间与温度、前体物质、水分和脂肪含量等均影响HAAs的形成。根据杂环胺形成影响因素,可以找到一些控制方法,例如选择合适的烹饪方式、适当降低加热温度和时间、添加一些保水性物质防止水分迁移从而阻止前体物质流失等方法均可以降低加工肉制品中HAAs的形成;还可以通过添加乳酸菌等阻止杂环胺的代谢。杂环胺种类繁多,形成机制较复杂,目前,仍然有许多杂环胺的形成机制尚不明确,且其代谢机理也需做进一步的探讨。
