复合益生菌对荷斯坦犊牛生长性能、腹泻率和血清指标的影响
2022-03-19彭轶楠杜津昊穆永松王治业
季 彬, 彭轶楠, 叶 泽, 杜津昊, 穆永松, 王治业
(1.甘肃省科学院生物研究所,甘肃兰州 730030;2. 甘肃省微生物资源开发利用重点实验室,甘肃兰州 730030;3. 甘肃华瑞农业股份有限公司,甘肃张掖 734502)
我国全面进入“饲料禁抗、养殖减抗、产品无抗”的新时代,开发应用无残留、无耐药性、高效、安全环保的新型饲料添加剂成为畜牧养殖行业的重点(任津莹等,2020;印遇龙等,2020)。益生菌制剂又称活菌制剂、有益微生物、微生态制剂等,是将有益微生物从天然环境中分离筛选出来, 经过培养、 传代繁殖、 加工工艺而制成的一类活菌制剂,具有改善肠道环境、抗菌、降血脂、抗肿瘤、免疫调节等生理作用,且具有无残留、无耐药性、绿色环保等优点(刘政等,2020;阿热爱·巴合提等,2020;姚蒙蒙等,2019)。与使用抗生素治疗动物疾病相反, 使用益生菌制剂作为饲料添加剂能够增强动物机体免疫力,提高饲料利用率,改善养殖生态环境(田志梅等,2019;黄玉婷等,2018)。近年来随着人们消费结构的优化,我国牛肉供不应求,荷斯坦公犊牛养殖数量不断增加(李秋凤等,2017),但由于自身消化系统尚未发育完善, 导致荷斯坦公犊牛生长缓慢且极易发生腹泻等疫病, 严重影响其育肥效果。鉴于此,本研究探讨了不同剂量复合益生菌对荷斯坦公犊牛生长性能、 腹泻率和血清生化指标、抗氧化指标、免疫指标的影响,为益生菌在荷斯坦公犊牛养殖生产中的推广应用提供重要的参考价值。
1 材料与方法
1.1 试验材料 复合益生菌由枯草芽孢杆菌、乳酸菌、 曲霉发酵物、 复合酶制剂等组成, 活菌总数≥1.6×109cfu/g,购自山东省碧蓝生物科技有限公司;40 头体况良好荷斯坦公犊牛[日龄为(30±2)d,体重为(49.23±1.15)kg],由甘肃华瑞农业股份有限公司奶牛养殖基地提供。
1.2 检测试剂盒 总蛋白(TP)、白蛋白(ALB)、球蛋白(GLB)、葡萄糖(GLU)、尿素氮(BUN)、碱性磷酸酶(AKP)、谷丙转氨酶(ALT)、谷草转氨酶(AST)、总抗氧化能力(T-AOC)、超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)、丙二醛(MDA)、免疫球蛋白A(IgA)、免疫球蛋白M(IgM)、免疫球蛋白G(IgG)、白介素2(IL-2)、白介素4(IL-4)、干扰素γ(IFN-γ)检测试剂盒均购自南京建成生物工程研究所。
1.3 试验动物分组与饲养管理 将40 头体况良好荷斯坦公犊牛随机分成4 组(对照组、 低剂量组、中剂量组、高剂量组),每组10 头牛,试验犊牛均单栏饲养,饲喂酸化乳和开食饲料,酸化乳每日饲喂3 次,每次饲喂2 L,饲料自由采食。 对照组、低剂量组、 中剂量组、 高剂量组分别在基础饲料(基础日粮组成及营养水平见表1) 中添加0、0.05%、0.1%、0.2%的复合益生菌,试验期为60 d,试验期间犊牛自由饮水, 保持水温恒定, 定时清粪、清洗消毒,保持圈舍干净卫生。 在试验开始和结束时分别称量体重, 记录试验期间每头犊牛的耗料量,在试验结束时对每头犊牛采集血液5.0 mL,分离血清。
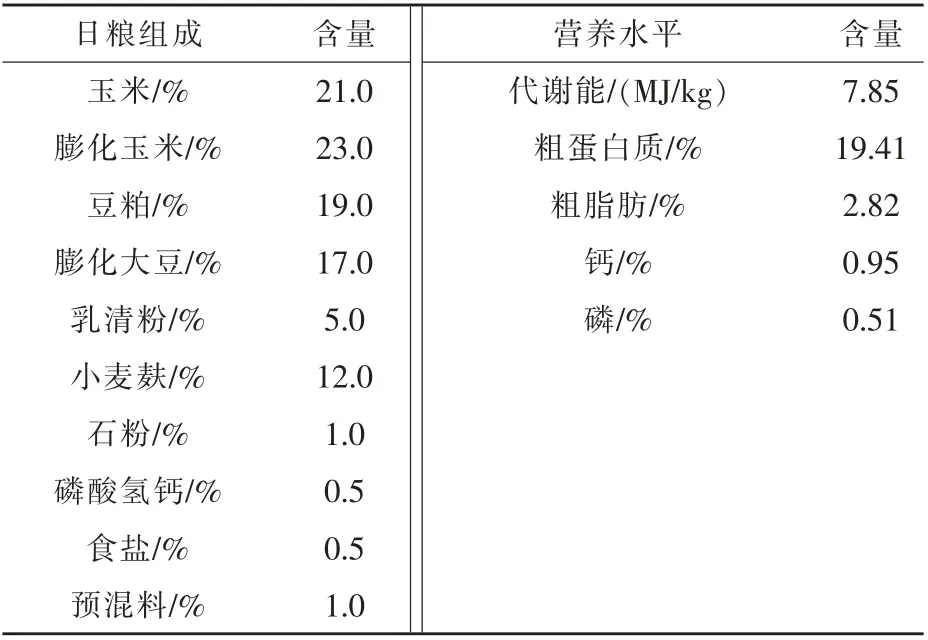
表1 基础日粮组成及营养水平
1.4 测定指标与方法
1.4.1 生长性能测定 分别称量每头犊牛初重和末重,计算日均增重[日均增重=(末重-初重)/试验天数]。 每日记录犊牛的耗料量,计算日均采食量(日均采食量/kg=耗料总量/试验天数)。 根据日均采食量和日均增重计算料重比 (料重比=日均采食量/日均增重)。
1.4.2 腹泻率测定 每日早晚观察犊牛腹泻情况,计算腹泻率(腹泻率/%=腹泻天数/试验天数)。
1.4.3 血清生化指标测定 分别利用TP、ALB、GLB、GLU、BUN、AKP、ALT、AST 检测试剂盒依次测 定 血 清 样 品 中 的TP、ALB、GLB、GLU、BUN、AKP、ALT、AST 含量。
1.4.4 血清抗氧化指标测定 分别利用T-AOC、SOD、GSH-Px、CAT、MDA 检测试剂盒依次测定血清样品中的T-AOC、SOD、GSH-Px、CAT、MDA 含量。
1.4.5 血清免疫指标测定 分别利用IgA、IgM、IgG、IL-2、IL-4、IFN-γ 检测试剂盒依次测定血清样品中的IgA、IgM、IgG、IL-2、IL-4、IFN-γ 含量。
1.5 数据统计分析 试验数据采用SPSS 13.0 软件进行方差比较分析,P<0.05 表示差异显著。
2 结果与分析
2.1 生长性能的测定结果 由表2 可知,中剂量组的末重、 日均增重较对照组分别提高2.79%、5.97%(P<0.05),料重比较对照组降低1.74%(P<0.05)。 高剂量组的末重、日均增重、日均采食量较对照组分别提高3.96%、8.43%、5.45%(P<0.05),料重比较对照组降低2.61%(P<0.05)。高剂量组的日均增重较中剂量组提高2.32%(P<0.05)。说明随着犊牛日粮中复合益生菌添加剂量的增加,其生长性能逐步提高。
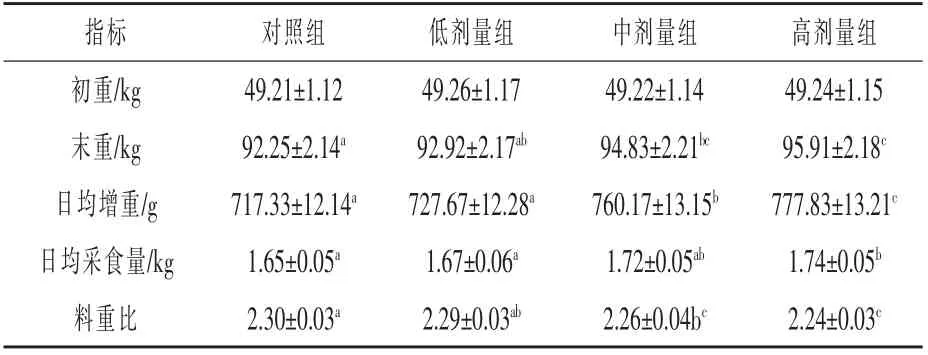
表2 生长性能的测定结果
2.2 腹泻率的测定结果 由表3 可知,低剂量组、中剂量组、 高剂量组的腹泻率较对照组分别降低22.27%、48.94%、71.07%(P<0.05)。 高剂量组的腹泻率较低剂量组、 中剂量组分别降低62.78%、43.35%(P<0.05)。 说明随着犊牛日粮中复合益生菌添加剂量的增加,其腹泻率逐步降低。

表3 腹泻率的测定结果%
2.3 血清生化指标的测定结果 如表4 所示,中剂量组、 高剂量组的TP 含量较对照组分别提高4.79%、6.19%(P<0.05),ALB 含量较对照组分别提高8.14%、10.45%(P<0.05),GLU 含量较对照组 分 别 提 高10.50%、11.78%(P<0.05),BUN 含量较对照组分别降低4.39%、6.48%(P<0.05)。说明随着犊牛日粮中复合益生菌添加剂量的增加,其血清生化指标逐步得到改善。
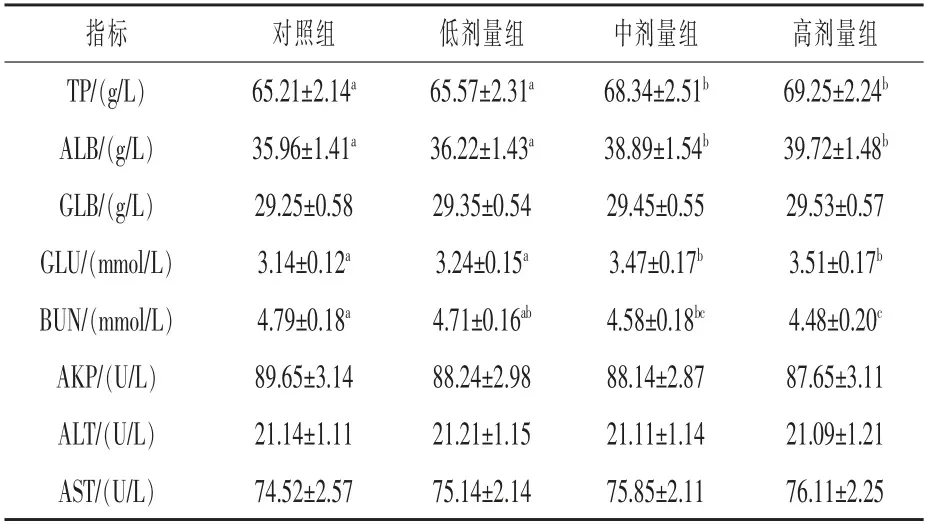
表4 血清生化指标的测定结果
2.4 血清抗氧化指标的测定结果 由表5 可知,中剂量组、 高剂量组的T-AOC 含量较对照组分别提高7.32%、9.27%(P<0.05),SOD 含量较对照组分别提高5.16%、8.19%(P<0.05),GSH-Px 含量较对照组分别提高3.11%、3.58%(P<0.05),MDA 含量较对照组分别降低8.19%、9.08%(P<0.05)。 高剂量组的SOD 含量较中剂量组提高2.87%(P<0.05)。 说明随着犊牛日粮中复合益生菌添加剂量的增加, 其血清抗氧化指标逐步得到改善。

表5 血清抗氧化指标的测定结果
2.5 血清免疫指标的测定结果 由表6 可知,低剂量组的IgG 含量较对照组提高3.31%(P<0.05)。 中剂量组、高剂量组的IgA 含量较对照组分别提高7.95%、10.22%(P<0.05),IgG 含量较对照组分别提高11.91%、15.84%(P<0.05),IL-2含量较对照组分别提高4.92%、6.73%(P<0.05)。高剂量组的IgG 含量较中剂量组提高3.51%(P<0.05)。 说明随着犊牛日粮中复合益生菌添加剂量的增加,其血清免疫指标逐步得到改善。
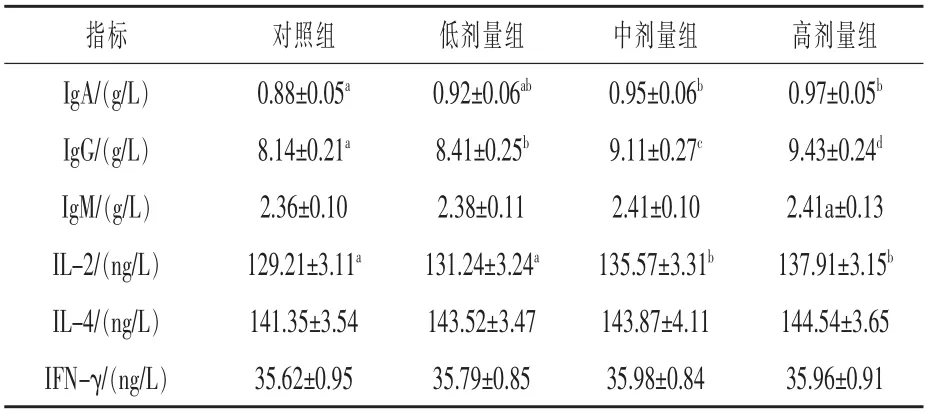
表6 血清免疫指标的测定结果
3 讨论
3.1 复合益生菌对荷斯坦犊牛生长性能的影响益生菌能够提高反刍动物的生长性能, 但由于反刍动物品种、日龄、饲养方式以及益生菌组成、活菌数量等的不同, 临床中益生菌的最适添加剂量存在较大差异。杨明等(2020)比较分析了青海犊牦牛(12 月龄)中添加0.5、1.0 g/kg 的复合益生菌(主要成分为芽孢杆菌、双歧杆菌,活菌数为2.0×109cfu/g)的作用效果,确定1.0 g/kg 为临床最适添加剂量。 鲍玉林等(2017)比较分析了健康肉羊 (体重为25 kg) 中添加2.5%、5.0%、10.0%复合益生菌(主要成分为乳酸杆菌、双歧杆菌、枯草芽孢杆菌、粪尿球菌、酵母菌,活菌数为3.6×1010cfu/g)的作用效果,确定5.0%为临床最适添加剂量。 本研究比较分析了荷斯坦公犊牛 (日龄为30 d, 体重为49.23 kg) 中添加0.05%、0.1%、0.2%复合益生菌(主要成分为枯草芽孢杆菌、乳酸菌、曲霉发酵物、复合酶制剂,活菌数为1.6×109cfu/g)的作用效果,确定0.2%为临床最适添加剂量。 杨明等(2020)、鲍玉林等(2017)以及本研究确定的临床最适添加剂量均存在较大差异,故在选择使用益生菌制剂时,应根据自身养殖的反刍动物品种、日龄、饲养方式以及益生菌组成、 活菌数量等确定适合自身的最适添加剂量,切勿盲目添加。
3.2 复合益生菌对荷斯坦犊牛腹泻率的影响犊牛由于自身消化系统尚未发育完全, 肠道微生物菌群不稳定, 饲养环境的微弱变化极易引起犊牛腹泻, 腹泻一直是犊牛养殖过程中的常见病和多发病, 发生腹泻的犊牛消化功能迅速降低,对营养物质的消化吸收利用率降低,导致生长发育缓慢(肖凡等,2019)。 益生菌进入反刍动物机体后, 通过与病原菌群黏附位点的时空竞争、 营养竞争作用, 限制肠道中有害菌的定植、生存及繁殖,对有益菌生长产生扶持,通过控制有益菌群与有害菌群的比例来调节动物机体内微生物菌落平衡,调节犊牛肠道健康,提高消化功能,降低腹泻率的发生(鲁陈等,2018)。在本研究中, 荷斯坦公犊牛日粮中添加0.05%、0.1%和0.2%复合益生菌均能够显著降低犊牛的腹泻率,且随着复合益生菌添加剂量的增加,犊牛腹泻率逐渐下降, 表明益生菌能够提高犊牛的消化功能,降低腹泻率。
3.3 复合益生菌对荷斯坦犊牛血清生化指标的影响 血液是动物机体内环境与外环境之间联系的纽带, 其在动物体内各组织器官之间的相互沟通联系过程中发挥重要作用, 各种养分和代谢产物均需要通过血液进行交换, 动物机体的各种生理变化均能在血液中体现出来, 血清中TP 含量能够反映出动物机体对蛋白质的消化和吸收利用程度,TP 含量较高则表明动物体内的蛋白质的合成作用增强; 血清中ALB 在维持血浆渗透压和提供动物机体蛋白质中发挥直接作用,ALB 含量较高则表明动物机体新陈代谢加强; 血清中GLU 含量是反映动物机体能量平衡状况的重要指标,GLU 含量较高则表明动物机体的能量代谢作用增强; 血清中BUN 是蛋白质代谢后的终产物, 其是反映动物机体蛋白质代谢和日粮中氨基酸平衡状况的重要指标,GLU 含量较低则表明动物机体的蛋白质沉积作用增强(郭凯等,2019)。 在本研究中,犊牛日粮中添加0.1%和0.2%复合益生菌能够显著提高犊牛血清中的TP、ALB、GLU 含量, 降低犊牛血清中的BCN 含量,该结果与杨明等(2020)在肉羊上的研究结果一致,其研究表明,肉羊日粮中添加复合益生菌能够显著提高肉羊血清中TP和ALB 含量,降低BCN 含量。 本研究通过检测血清中TP、ALB、GLU、BCN 含量表明益生菌能够提高蛋白质的合成, 增加犊牛对蛋白质的消化和吸收。
3.4 复合益生菌对荷斯坦犊牛血清抗氧化指标的影响 动物机体的抗氧化能力对自身健康至关重要,当机体受到自身或外来因素刺激时,机体氧化还原状态失去平衡,引发氧化应激反应,产生大量自由基和脂质过氧化产物, 当自由基生成过剩或清除不足时, 将会导致机体发生氧化性损伤和功能性障碍。 MDA 是不饱和脂肪酸在氧自由基攻击下产生的脂质过氧化终产物,血清MDA 含量变化可以直接反映机体内脂质过氧化的程度,间接反映机体的氧化损伤程度,MDA 含量减少则表明动物机体抗氧化能力加强;SOD、GSH-Px 均是动物体内清除自由基酶促系统中的重要抗氧化酶, 能够特异性清除氧自由基,对发挥抗氧化能力具有重要作用;T-AOC是综合反映机体抗氧化能力强弱的指标 (金宇航等,2021)。 李宁等(2020)研究表明,犊牛日粮中每日添加2 g 活性干酵母能够显著提高犊牛的SOD 活性。彭涛等(2020)研究表明,舍饲山羊饲粮中添加0.4%复合益生菌能够显著提高山羊血清的T-AOC 和SOD 活性, 显著降低MDA 含量。 本研究结果表明, 犊牛日粮中添加0.1%和0.2%复合益生菌能够显著提高犊牛的T-AOC、SOD 和GSH-Px 活性, 显著减低MDA 含量,与李宁等(2020)、彭涛等(2020)的研究结果具有一致性, 表明益生菌确实能够提高犊牛的抗氧化能力。
3.5 复合益生菌对荷斯坦犊牛血清免疫指标的影响 免疫球蛋白(Ig)是一类由浆细胞产生的具有抗体活性的球蛋白, 在抗体介导的多种免疫防御机制中起重要作用,其中IgA 由浆细胞分泌产生, 在黏膜免疫中发挥重要作用;IgG 通过吞噬、凝集、沉淀抗原发挥抑菌、抗病毒、中和毒素等免疫活性(殷术等,2019)。 细胞因子是由免疫细胞产生的介导机体免疫应答和炎症反应一类高活性小分子多肽, 其中IL-2 是淋巴细胞活化过程中由细胞膜分泌的一种糖蛋白, 不但能够刺激T 细胞生长、增殖、分化,也是重要的免疫增强因子。 鲍玉林等(2017)研究表明,犊牦牛饲料中添加0.5 g/kg 和1.0 g/kg 益生菌均能显著提高青海犊牦牛血清的IgG、IL-2 含量,且1.0 g/kg 添加剂量的作用效果显著优于0.5 g/kg 的添加剂量。 在本研究中,犊牛日粮中添加0.05%复合益生菌均能够显著提高犊牛血清的IgG 含量,添加0.1%和0.2%复合益生菌均能够显著提高犊牛血清的IgA、IgG、IL-2 含量, 表明随着复合益生菌添加剂量的增加, 其对犊牛血清免疫指标的改善重要性逐渐增强。 本研究结果与鲍玉林等(2017)的研究结果具有一致性,表明益生菌确实能够提高犊牛的免疫力。
4 结论
在本试验条件下, 犊牛日粮中添加0.1%和0.2%复合益生菌均能够提高犊牛的生长性能,降低犊牛腹泻率,改善犊牛血清生化指标、抗氧化指标和免疫指标, 添加剂量为0.2%时效果最为显著。
