昆明犬IGF2基因克隆、生物信息学分析及其时空表达研究
2022-03-17韦云芳李飞翔黄庆国李居东万九生陈方良
韦云芳,李飞翔,星 云,黄庆国,李居东,万九生,陈方良
(1.公安部昆明警犬基地,昆明 650204;2.警犬技术公安部重点实验室,昆明 650204;3.云南农业大学动物科学技术学院,昆明 650201)
作为国内独一无二的国产警用工作犬品种,昆明犬于2007年被审定为犬类国家级畜禽新品种,编入《中国畜禽遗传资源名录》和《世界粮食与农业动物遗传资源状况》。昆明犬属于中国七大警用工作犬品种之一,已广泛配发到国内33个省级行政区的警犬使用部门及一线实战单位,在治安防范、消防搜救、护卫搜捕、安保维稳及反恐处突等重要任务中发挥其独特作用[1]。目前昆明犬已出口到朝鲜、新加坡、巴基斯坦、越南、柬埔寨等10多个国家,在世界工作犬领域占有一席之地。作为中国宝贵的犬业种质资源,研究参与调控昆明犬生长发育的基因,深入阐明昆明犬生长发育规律,进而应用分子育种技术进一步提高其生长发育性能具有重要的理论和实际应用价值。
胰岛素样生长因子(insuline-like growth factors,IGFs)因其化学结构与胰岛素原类似而得名,是一类多肽因子,对动物生长发育、繁殖、营养代谢及组织细胞的增殖、分化、凋亡均有重要调控作用[2-3]。IGF2作为IGFs家族中重要成员之一,又被称为生长调节素A(somatomedin A),广泛参与组织细胞的增殖分化、机体肌肉发育和体脂沉积及肿瘤细胞增殖等众多生理过程[4-5]。分析IGF2基因结构功能并阐明其表达规律,在畜牧生产中具有重要意义。目前,IGF2作为农业动物重要经济性状的候选基因,在多种农业和经济类动物中已有较多研究。众多研究发现,IGF2基因遗传变异与猪[6-8]、牛[9-11]、禽类[12-13]及经济型鱼类[14]和鳖类[15]等动物的生长发育存在显著相关关系,并鉴定出了一些显著影响目标性状的分子标记,这些研究为在昆明犬上开展该基因的研究奠定了理论基础。
在工作犬领域,体型大小是工作犬筛选中最重要的性状指标之一。了解昆明犬体型等生长发育性状的遗传因素,可为研究人员提供一个改进动物体型和生长发育性状的途径[16]。警犬繁育中,2.5月龄的仔犬是考察和挑选的关键期,其体高、体长及体重的增长速度十分迅速,体型大小出现明显差异,对调控其此时期生长发育相关的基因进行研究具有十分重要的意义。基于IGF2基因在生长发育调控中发挥的重要作用,本研究首先克隆昆明犬IGF2基因CDS编码区,并利用生物信息学方法分析其序列结构、进化关系及编码蛋白的功能,同时在mRNA水平上探究IGF2基因在2.5月龄犬各组织和昆明犬不同生长时期肝脏中的表达水平,以期为今后深入研究IGF2基因调控昆明犬生长发育过程的作用机制提供参考。
1 材料与方法
1.1 试验动物及样品采集
供试的昆明犬均源自公安部昆明警犬基地,随机选取仔犬期(2.5月龄)、幼犬期(6月龄)、青年期(1岁)、成年期(1.7和2.5岁)正常饲养的昆明犬各3头,麻醉后采集心脏、肝脏、脾脏、肺脏、肾脏及大腿内侧肌肉组织样品,置于液氮中保存备用。
1.2 主要试剂及仪器
TRIzol RNA分离试剂购自Invitrogen公司;反转录和实时荧光定量PCR试剂盒均购自Vazyme公司;2×PCR Buffer、dNTP Mixture、Taq酶等PCR相关试剂均购自生工生物工程(上海)股份有限公司;胶回收纯化试剂盒、pMD18-T载体、大肠杆菌DH5α感受态细胞均购自宝生物工程(大连)有限公司。冷冻离心机(5430R)购自Eppendorf公司;电泳仪(DYY-6B)购自北京六一仪器厂;超微量紫外分光光度计(NanoDrop 2000)购自Thermo Scientific公司;实时荧光定量PCR仪(Prism 7300)购自ABI公司;梯度PCR仪(T100)及凝胶成像系统(GelDoc XR+)均购自Bio-Rad公司。
1.3 引物设计及合成
根据NCBI上公布的家犬(Canislupusfamiliaris)IGF2基因mRNA序列(登录号:XM_005631053.3)设计CDS序列扩增引物和实时荧光定量引物,以GAPDH(登录号:NM_001003142.2)为内参基因,引物序列见表1。引物均由生工生物工程(上海)股份有限公司合成。

表1 引物信息
1.4 昆明犬IGF2基因CDS区扩增与克隆测序
按照TRIzol法提取2.5月龄昆明犬肝脏组织样品总RNA,通过超微量分光光度计检测RNA的质量,根据反转录试剂盒操作说明反转录合成cDNA。以昆明犬肝脏组织cDNA为模板进行PCR扩增。PCR扩增体系25 μL:PCR Buffer(2×)12.5 μL,dNTP混合物(10 mmol/L)0.2 μL,cDNA(100 ng/μL)1 μL,上、下游引物(10 μmol/L)各0.5 μL,Taq酶(5 U/μL)0.2 μL,补ddH2O至25 μL。PCR扩增条件:95 ℃预变性3 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸90 s,共35个循环;72 ℃延伸7 min;4 ℃保存。PCR产物经1.0%琼脂糖凝胶电泳检测后按胶回收试剂盒说明书进行胶回收,并按pMD18-T克隆载体试剂盒说明书进行连接反应。将连接产物转化至大肠杆菌DH5α感受态细胞中,挑取白色阳性单克隆菌落至LB(Amp+)液体培养基中扩大培养(37 ℃),最后进行菌液PCR鉴定,选择PCR鉴定正确的阳性单克隆送至生工生物工程(上海)股份有限公司进行测序。
1.5 昆明犬IGF2基因生物信息学分析
利用DNAStar软件将克隆序列与NCBI数据库中家犬(Canislupusfamiliaris,登录号:XM_005631053.3)、赤狐(Vulpesvulpes,登录号:XM_026004185.1)、猫(Feliscatus,登录号:XM_023240193.1)、猪(Susscrofa,登录号:NM_213883.2)、马(Equuscaballus,登录号:NM_001114539.2)、牛(Bostaurus,登录号:NM_174087.3)、羊(Ovisaries,登录号:NM_001009311.1)及鸡(Gallusgallus,登录号:NM_001030342.2)的IGF2基因核苷酸序列进行相似性比对,并构建系统进化树。IGF2蛋白的保守结构域、理化性质及结构特征分析等参照韦云芳等[16]方法进行。
1.6 昆明犬IGF2基因组织表达检测
按照TRIzol法提取2.5月龄昆明犬心脏、脾脏、肺脏、肾脏及大腿内侧肌肉组织及6月龄和1、1.7、2.5岁肝脏组织样品总RNA,并反转录合成cDNA。通过实时荧光定量PCR方法检测IGF2基因在2.5月龄昆明犬组织中及各年龄段昆明犬肝脏中表达量。PCR反应体系20 μL:cDNA 2 μL,2×Real-time PCR Master Mix 10 μL,上、下游引物(5 μmol/L)各0.4 μL,50×Rox Reference Dye 0.4 μL,补ddH2O至20 μL。PCR反应程序:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火60 s,共40个循环;同时以熔解曲线分析扩增产物的特异性,程序为95 ℃ 15 s;60 ℃ 60 s,95 ℃ 15 s。每个样品检测3次,取其平均值。以GAPDH为内参基因,对昆明犬不同组织和不同年龄段肝脏中的表达水平进行定量结果分析。
1.7 数据统计分析
采用2-ΔΔCt法计算IGF2基因的相对表达量,利用SPSS 22.0进行单因素方差分析(One-Way ANOVA),结果用平均值±标准差表示,用GraphPad Prism 5.0软件作图。P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 昆明犬IGF2基因克隆及测序
以昆明犬肝脏组织cDNA为模板,PCR扩增获得IGF2基因CDS编码区,扩增产物经1.0%琼脂糖凝胶电泳检测出现单一目的条带,片段大小约717 bp,与预期片段大小相符(图1)。测序后分析得到昆明犬IGF2基因片段长度为717 bp,编码238个氨基酸,与PCR结果一致。
2.2 昆明犬IGF2基因相似性比对和系统进化树构建
将昆明犬IGF2基因序列与其他物种序列进行相似性比对,结果显示,昆明犬与家犬、赤狐、猫、猪、马、牛、羊及鸡的相似性分别为100%、100%、94.1%、93.2%、89.6%、88.7%、88.3%及66.3%(图2)。系统进化树显示,昆明犬与家犬属于同一分支,与同为犬科动物的赤狐进化关系最近,与鸡的亲缘关系最远,构建的系统进化树直观地展示了不同物种之间IGF2基因进化的差异,与序列相似性比对结果一致(图3)。

M,DL10000 DNA Marker;1~2,昆明犬IGF2基因PCR扩增产物M,DL10000 DNA Marker;1-2,PCR amplification products of IGF2 gene in Kunming dogs图1 昆明犬IGF2基因PCR扩增结果Fig.1 PCR amplification results of IGF2 gene in Kunming dogs
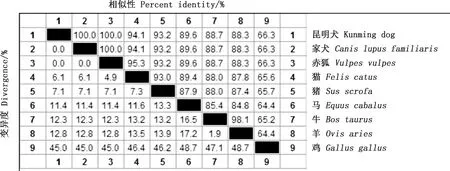
图2 不同物种IGF2基因核苷酸序列相似性比对Fig.2 Similarity alignment of nucleotide sequences of IGF2 gene in different species
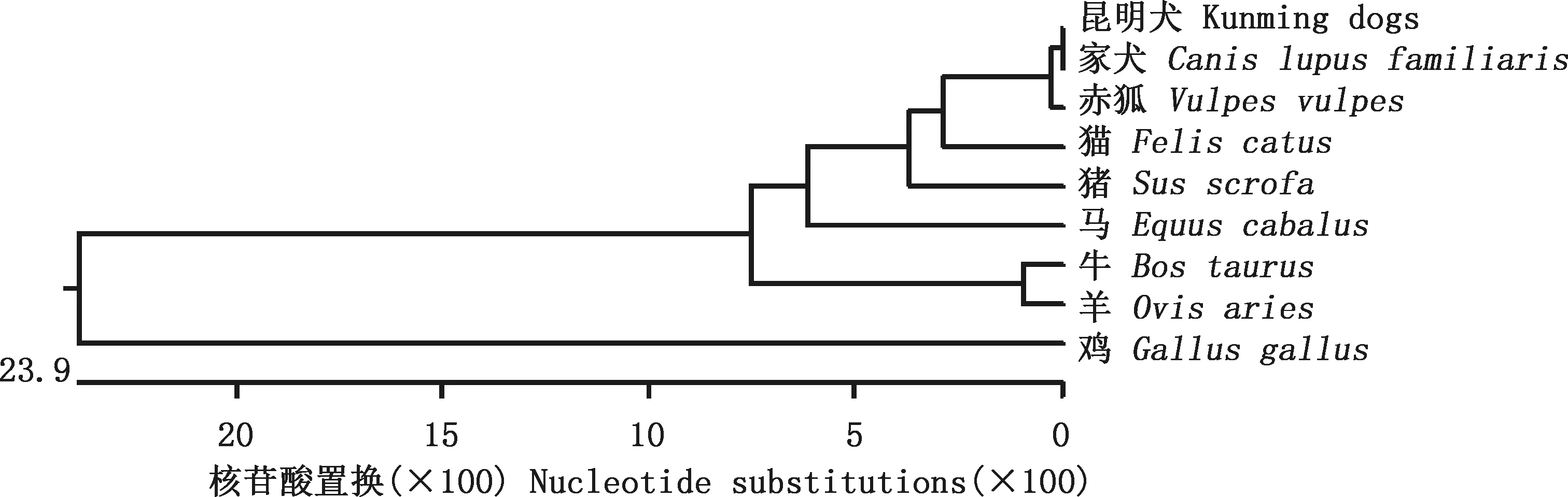
图3 不同物种IGF2基因核苷酸序列系统进化树Fig.3 Phylogentic tree of nucleotide sequence of IGF2 gene in different species
2.3 昆明犬IGF2蛋白结构和功能分析
2.3.1 理化性质及亲/疏水性分析 保守结构域分析表明,IGF2属于ILGF-like superfamily和IGF2-C superfamily超家族成员之一(图4)。ExPASy-ProtParam在线工具分析显示,IGF2蛋白分子质量为26.46 ku,等电点为9.61,属碱性蛋白,半衰期为30 h;不稳定系数为63.63,属不稳定蛋白;脂肪系数78.70,总平均亲水性为-0.241,属亲水性蛋白。IGF2蛋白共由22种氨基酸组成(表2),其中占比最高的是精氨酸(Arg,10.9%),最少的是天冬酰胺(Asn)和色氨酸(Trp),均为0.8%;带负电荷的残基总数(Asp+Glu)有22个,带正电荷的残基总数(Arg+Lys)有33个,推测IGF2蛋白可能带正电,肽链N-端为甲硫氨酸(Met)。昆明犬IGF2蛋白亲/疏水性分析结果表明,在第70位氨基酸处存在疏水最大值3.011,在第210位氨基酸处存在疏水最小值-2.344(图5),氨基酸序列亲水性残基数大于疏水性残基,整体表现为亲水性,属亲水性蛋白,这与平均亲水性指数相吻合。
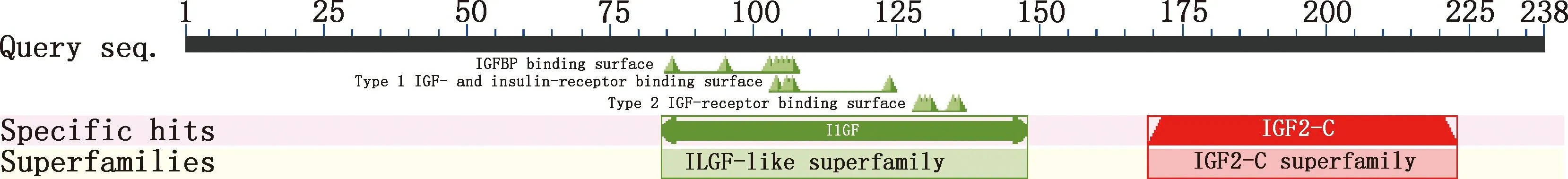
图4 昆明犬IGF2蛋白保守结构域预测Fig.4 Conserved domain prediction of IGF2 protein in Kunming dogs

表2 昆明犬IGF2蛋白氨基酸组成
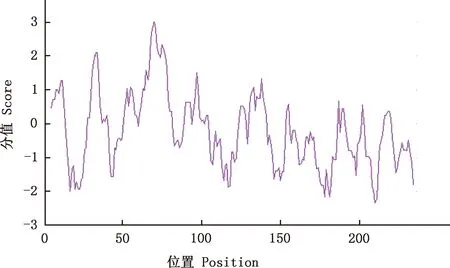
图5 昆明犬IGF2蛋白的疏水性分析Fig.5 Hydrophobicity analysis of IGF2 protein in Kunming dogs
2.3.2 信号肽、跨膜结构及亚细胞定位 通过SignalP 4.0在线软件分析表明,昆明犬IGF2蛋白不存在信号肽剪切位点(图6),属非分泌型蛋白。通过TMHMM 2.0在线软件分析跨膜结构发现,昆明犬IGF2蛋白不存在跨膜域,属非跨膜蛋白。使用PSORT Ⅱ在线软件预测昆明犬IGF2蛋白亚细胞定位发现,IGF2蛋白广泛分布于细胞核(47.8%)、线粒体(17.4%)、细胞质(13%)、质膜(8.7%)、内质网(4.3%)、细胞外(4.3%)及囊泡分泌系统(4.3%)。
图6 昆明犬IGF2蛋白信号肽预测Fig.6 Signal peptide prediction of IGF2 protein in Kunming dogs
2.3.3 蛋白磷酸化位点 运用NetPhos 3.1在线软件分析蛋白磷酸化位点显示,昆明犬IGF2蛋白存在23个磷酸化位点,其中丝氨酸(S)14个、苏氨酸(T)9个,不存在酪氨酸(Y)磷酸化位点,上述23个位点可能成为蛋白激酶磷酸化位点(图7)。
2.3.4 二级结构、三级结构预测 通过SOPMA预测昆明犬IGF2蛋白二级结构,发现IGF2蛋白结构以α-螺旋(32.77%)和无规则卷曲(47.48%)为主,延伸链和β-转角分别占11.76%和7.98%(图8);使用SWISS-MODEL在线软件预测该蛋白的三级结构发现,昆明犬IGF2蛋白的三级结构主要为α-螺旋和无规则卷曲(图9),与二级结构预测结果相符。
2.4 昆明犬IGF2基因组织表达谱
由图10可知,IGF2基因在昆明犬心脏、肝脏、脾脏、肺脏、肾脏和肌肉中均有表达,在肝脏和脾脏中的表达极显著高于其他组织(P<0.01);在肺脏、肾脏和肌肉中的表达量显著高于心脏中的表达量(P<0.05)。
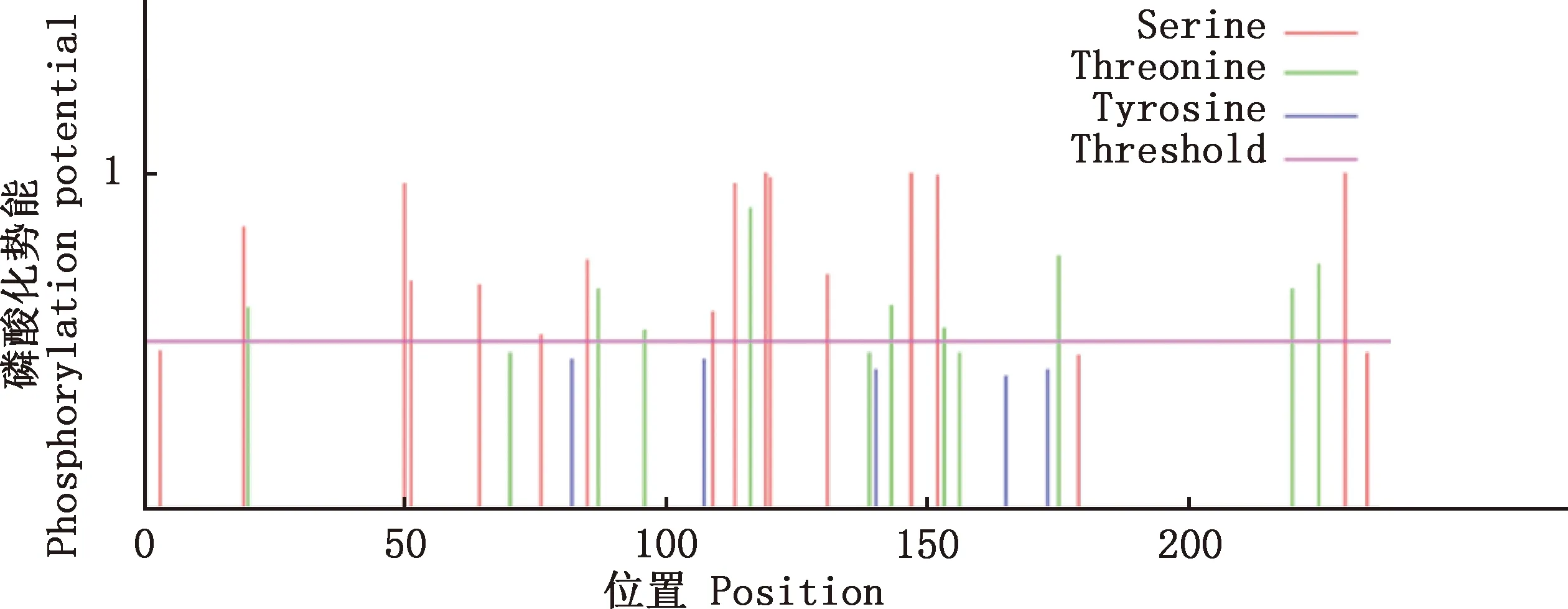
图7 昆明犬IGF2蛋白磷酸化位点预测Fig.7 Phosphorylation site prediction of IGF2 protein in Kunming dogs

线条由长到短分别代表α-螺旋、延伸链、β-转角和无规则卷曲Lines from long to short represent alpha helix,extended chain,beta turn and random coil,respectively图8 昆明犬IGF2蛋白二级结构预测Fig.8 Secondary structure prediction of IGF2 protein in Kunming dogs
图9 昆明犬IGF2蛋白三级结构预测Fig.9 Tertiary structure prediction of IGF2 protein in Kunming dogs
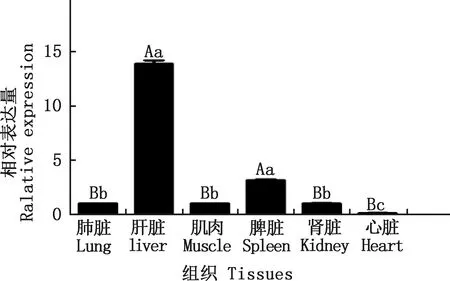
肩标不同小写字母表示差异显著(P<0.05);肩标不同大写字母表示差异极显著(P<0.01);肩标相同字母表示差异不显著(P>0.05)。下同Values with different small letter superscripts mean significant difference (P<0.05); And with different capital letter superscripts mean extremely significant difference (P<0.01); While with the same letter superscripts mean no significant difference (P>0.05). The same as below图10 IGF2基因在昆明犬不同组织中的表达Fig.10 Expression of IGF2 gene in different tissues of Kunming dogs
2.5 IGF2基因在昆明犬不同生长阶段肝脏中的表达
对昆明犬仔犬期(2.5月龄)、幼犬期(6月龄)、青年期(1岁)及成年期(1.7和2.5岁)不同生长阶段肝脏中IGF2基因的表达量进行分析,结果显示,IGF2基因在2.5月龄昆明犬肝脏脏组织中表达量极显著高于其他生长阶段(P<0.01),在幼犬期(6月龄)及以上生长阶段中表达较弱,但大致能看出随着年龄的增加呈现出微弱降低趋势(图11)。
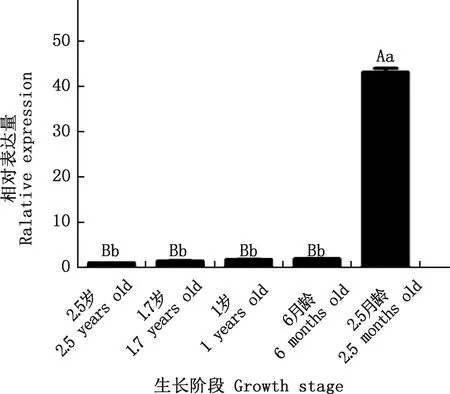
图11 IGF2基因在不同生长阶段昆明犬肝脏中的表达Fig.11 The expression of IGF2 gene in liver at different growth stages of Kunming dogs
3 讨 论
IGF2是最早被证实的印迹基因之一,在遗传水平上受亲本印迹的调控,主要由父系来源染色体表达[17]。作为一种广泛表达的多功能单链多肽,成年时期主要靠肝脏分泌,而胚胎时期则是在子宫内通过胎盘分泌,是胚胎生长和发育过程中的重要调节因子[18-19]。研究表明,在人胚胎时期IGF2基因过表达可能导致初生婴儿罹患伯-韦综合征(Beckwith-Wiedemann syndrome),表现为组织器官的不对称发育和过度生长,即单侧肥大和生长紊乱;而IGF2基因表达不足或缺失则可能导致罗素银综合征(Silver-Russell syndrome),表现为发育迟缓和身体矮小畸形[20-21]。小鼠中靶向敲除IGF2基因则会导致胎儿生长发育缺陷[17]。IGF2基因不仅在哺乳动物胚胎生长和发育中起着非常重要的作用,而且还参与动物的多种生理和病理调控过程[22-25]。在猪和其他哺乳动物上,IGF2基因启动子上转录抑制物结合位点的单核苷酸多态性改变会影响启动子的活力,进而改变骨骼肌中IGF2基因的表达水平,从而调控肌肉的生长发育[26-27]。基于IGF2基因在调控动物生长发育等方面的重要作用,本研究在犬上针对该基因进行克隆、序列分析及组织表达分析。
目前,关于哺乳动物IGF2的研究更多的是集中于IGF2在动物血液中的浓度水平和IGF2基因多态性与生产性能的关系及其与肿瘤发生的关系。关于哺乳动物IGF2基因序列分析和结构变异的研究较少。本研究首次从昆明犬中成功克隆得到IGF2基因CDS区,序列长度为717 bp,共编码238个氨基酸。将昆明犬IGF2基因核苷酸序列与多个物种进行相似性比对,发现其核苷酸序列与同为犬科动物的赤狐的相似性为100%,初步说明IGF2基因进化在犬科动物中是相当保守的。另外,多物种IGF2基因核苷酸系统进化树与各物种在分类学的地位基本吻合,且与物种进化的结果相一致。本试验运用多个生物信息学软件对昆明犬IGF2蛋白进行预测,发现IGF2蛋白的不稳定指数为63.63,亲水性为-0.241,无信号肽,无跨膜结构,属不稳定的亲水性非分泌蛋白。在辽宁绒山羊的研究中发现IGF2蛋白具信号肽结构,为不稳定的亲水性分泌蛋白[28],推测该蛋白可能存在物种间表达调控模式上的差异。昆明犬IGF2蛋白主要定位于细胞核和线粒体中,有23个磷酸化位点,是进行蛋白磷酸化修饰的潜在作用位点。二级结构中无规则卷曲占比最高(达47.48%),间接说明IGF2蛋白稳定性较差,这与其不稳定指数远远高于40(63.63)的结果相符。
尽管IGF2基因在序列结构上高度保守,但在不同物种中具有不同的表达模式[29]。IGF2基因在动物各组织中广泛表达,是控制动物生长和脂肪沉积的重要基因之一[30-31]。本研究中,IGF2在不同组织中出现一定的差异表达,其在肝脏中的表达最高,进一步说明肝脏是IGF2合成的主要器官[32]。除肝脏以外,该基因在脾脏的表达量显著高于其他组织,由于脾脏是动物体内最大的淋巴免疫器官,所以推测IGF2可能参与到昆明犬的免疫调节途径中,但其具体机制还有待于进一步研究。目前,对于IGF2在免疫中所发挥的作用,也是研究者们较为关注的问题。另外,各组织中IGF2基因的表达量可能会随着机体的发育阶段而呈现出差异,而2.5月龄犬生长发育旺盛,是仔犬向幼犬过渡的关键期,此时期IGF2基因在各组织中的表达情况尤其值得关注。本研究发现IGF2基因在2.5月龄仔犬肝脏中的表达量极显著高于后期各个成长阶段,这一结果从侧面印证了IGF2在动物机体早期生长发育中发挥着极为重要的作用[3,33]。Goodyer等[34]的研究结果也显示,胎儿肝脏细胞产生的IGF2是IGF1的10倍。IGF2基因在不同年龄的猪、牛等家畜背最长肌中的表达量也表现出随年龄增加而下降的现象[35-36]。此外,IGFs的高亲和力结合伴侣胰岛素样生长因子4(IGFBP4)和IGFBP5在牦牛不同生长阶段肝脏中的表达量与其年龄增长呈正相关,表现为随年龄增加表达量也增加,并于5岁时达到峰值[37],由此推测昆明犬不同生长发育时期肝脏中可能也存在上述IGFs结合伴侣随年龄增加而出现表达量上升的现象,从而可能导致IGF2在不同生长时期肝中表达量与年龄增长呈负相关,表明IGFs与其结合伴侣以协同方式共同调控动物组织的生长发育。后续研究可进一步探索在犬类早期胚胎和幼体发育过程中IGF2的具体生物学作用和调控机制。
4 结 论
本研究克隆了昆明犬IGF2基因,其编码区全长717 bp,编码238个氨基酸,昆明犬IGF2相似性与赤狐最高(100%),证明其在生物进化过程中非常保守。IGF2蛋白属于带正电、不稳定、亲水性蛋白,主要分布在细胞核和线粒体中;二级结构以α-螺旋和无规则卷曲为主。IGF2基因在2.5月龄昆明犬心脏、肝脏、脾脏、肺脏、肾脏和肌肉中均有表达,其中肝脏组织表达量最高,该基因在肝脏中的表达随着昆明犬年龄的增加呈现降低的趋势。研究结果加深了对犬类IGFs分子生物学方面的认识,可为昆明犬生长调控及分子标记辅助育种的研究提供一定理论依据和支持。
