油菜盐胁迫响应miRNA及其靶基因鉴定
2022-03-16王越周婷岳彩鹏黄进勇华营鹏
王越,周婷,岳彩鹏,黄进勇,华营鹏
(郑州大学农学院,河南 郑州,450001)
土壤盐碱化是全球重要的环境和农业问题。据估计,盐胁迫可能影响全球一半的可耕地。土壤盐胁迫严重抑制了作物生长发育并显著降低了作物的产量[1],其主要原因是盐胁迫会导致高渗透胁迫、氧化应激、营养失衡、离子毒性、生物膜紊乱,并严重破坏细胞分裂及关键代谢等过程,导致水势下降,从而限制水分的吸收和细胞的扩张[2]。为了在盐胁迫条件下生存下来,植物已经进化出各种复杂的机制来缓解盐胁迫对自身造成的损伤[3,4]。因此,了解盐胁迫的分子机制对于提高作物耐盐性具有重要意义[5]。
MicroRNA(miRNA)是一类由内源基因编码的长度约为18~24 nt 的非编码单链RNA 分子,在转录后和翻译水平负调控基因表达[6]。越来越多的证据表明,miRNA 参与了植物对非生物胁迫的响应[6~10],如温度[11]、干旱[12]和盐[13,14]胁迫。在拟南芥盐胁迫反应中,miR156、miR167、miR168 和miR396 等多个miRNA 存在差异表达[15]。过表达水稻miR319a的匍匐禾草(Agrostis stolonifera)表现出更强的耐盐性[16],其原因是盐胁迫诱导miR319 表达升高,导致至少4个miR319 的靶基因(AsPCF5、AsPCF6、AsPCF8和AsTCP14)和AsNAC60的表达下调,从而提高了植株的盐胁迫抗性。过表达miR396和miR394的棉花和拟南芥表现出高度的盐胁迫敏感性[17~19],其原因是miR396 和miR394 可能作为一种负调控因子,分别以GRF 和LCR 等调控蛋白为靶点。其中GRF 抑制拟南芥的应激反应基因AtDREB2A的表达,增加了植物对渗透胁迫的耐受性,LCR 则以ABA 依赖性方式参与拟南芥盐胁迫抗性反应。盐胁迫激活WRKY并调节植物中胁迫耐受性相关途径越来越受关注[15]。前人研究表明,拟南芥RPD3-like 组蛋白去乙酰化酶HDA9 通过调节WRKY53 的DNA 结合能力和转录活性来降低拟南芥的盐胁迫抗性[20]。近期研究发现苹果中MdSPL13 与MdWRKY100启动子区结合,并正向调节MdWRKY100的表达,通过促进基因转录,如抗氧化剂生物合成,糖代谢和脯氨酸生物合成有关基因的转录,进一步维持ROS 和渗透平衡,从而增强盐胁迫的耐受性[21]。植物在盐胁迫下的生存需要对胁迫条件的快速感知和适应。前人研究[22]表明RACK1A诱导的miR393 生物发生,下调了生长素信号,可能以此调节拟南芥对盐度的适应。最近研究表明[23]通过表达TIR1,增加植物生长素信号传导,可能会触发植物生长素介导的下游途径,以通过渗透压调节和增加Na+外排来增强植物对盐胁迫的抗性。这一发现使我们更好地理解miR393 介导生长素调节的盐胁迫反应。上述研究结果均表明,miRNA 在植物的盐胁迫抗性反应中发挥重要作用。
油菜(AnAnCnCn,2n=4x=38)是世界上重要的油料作物之一,起源于其二倍体祖先Brassica rapa(ArAr, 2n=2x=20)和Brassica oleracea(CoCo, 2n=2x=18)的自发杂交[24]。目前,在拟南芥以及甘蓝型油菜等作物中,已有一些耐盐性相关的基因被克隆与鉴定。如脯氨酸合成的关键基因P5CS1[25],通过缓解渗透胁迫来参与油菜盐胁迫响应;控制油菜离子稳态的Na+/H+逆向转运蛋白基因BnNHX1[26],可在液泡中隔离Na+和K+,并通过维持Na+/K+稳态并增强渗透和抗氧化调节能力,赋予转基因油菜耐盐性。还有诸多转录因子参与调控耐盐性,如WRKY家族[27],NAC家族[28]等。尽管前人已经对油菜的盐胁迫抗性展开了系列研究,但由于油菜基因组庞大复杂,耐盐机制中许多问题仍有待探索。
本研究的目的是鉴定油菜生长发育过程中对响应盐胁迫的miRNA 及其核心靶基因并研究其在盐胁迫中发挥的作用。本研究在200 mmol·L-1NaCl(T)和无盐处理对照(C)条件下培养油菜,分别从地上部(S)和根(R)中取样,提取总RNA 后构建了12个miRNA 文库。通过使用高通量测序技术分析了12个miRNA测序文库,最终鉴定到了稳定差异表达的26个miRNA,并根据与相应的转录组表达谱的比较预测了它们的靶基因。这些鉴定到的差异表达miRNA 及其核心靶基因有助于深入了解植物盐胁迫抗性的分子机制,并为油菜盐胁迫抗性的遗传改良提供了优异的基因资源。
1 材料与方法
1.1 植物材料及试验设置
油菜种子为中双11,由郑州大学黄进勇教授提供,在25℃黑暗条件下催芽约24 h,挑选露白一致的油菜种子均匀摆放在铺有滤纸的育苗盘中,放于温室25℃±3℃培养架上培养,幼苗长到5 cm 左右选取生长一致的健壮幼苗,移栽到盛有适量霍格兰营养液的黑色塑料盆中水培生长,每5 d 更换一次营养液[29]。试验在光照培养室中进行,生长环境设定如下:光照强度150 μmol/(m2·s),室温24℃(白天)/22℃(夜晚),光照周期16 h(光照)/8 h(黑暗),相对湿度60%。
全转录组测序是在种子萌发7 d 后,选取长势一致的油菜幼苗,将其培养在不含NaCl的条件下生长10 d,随后转移到含200 mmol·L-1NaCl 的溶液中培养12 h 直至取样。处理组和对照组油菜幼苗分别取其地上部和根部,每个部位均包含3 个生物学重复。
1.2 mRNA文库构建和测序
mRNA 文库构建、测序详情以及本文所用的转录组数据参考本课题组已经发表的文章[30]。本文中重点分析油菜盐胁迫响应的miRNA及其靶基因。
1.3 sRNA文库的构建和测序
使用TRIzol法提取油菜总RNA,用Nanophotometer®分光光度计(IMPLEN, CA, USA)检测RNA 纯度,使用Qubit®RNA 检测试剂盒在Qubit®2.0 荧光计(Life Technologies, CA, USA)中检测其浓度。为了评估RNA 的完整性,使用Agilent Bioanalyzer 2000 system(Agilent Technologies,CA,USA)的RNA Nano 6000 检测试剂盒测定RNA 的完整性值。随后使用Illumina®的NEBNext®Multiplex 小RNA 文库制备试剂盒(NEB,USA)构建小RNA 文库,将索引代码添加到每个样本的属性序列中,每个样品共使用约3.0 μg RNA。在miRNA 的3'和5'端添加特定的索引代码,然后使用M-MuLV 逆转录酶(RNase H-)作为催化剂合成第一链cDNA。使用LongAmp Taq 2× Master Mix、Illumina 小RNA 引物和index 接头引物进行PCR 扩增。PCR 产物经8%聚丙烯酰胺凝胶电泳纯化,回收140~160 bp 的DNA 片段。使用Agilent Bioanalyzer 2100 system(Agilent Technologies,CA,USA)对文库质量进行评估。随后使用TruSeq SR Kit v3-cBOt-HS(Illumina)在cBot 集群生成系统上对索引代码样本进行聚类。集群生成后,在Illumina Hiseq 2500平台上对库进行测序,生成50 bp的单端reads。
1.4 数据过滤和读取
测序完成后,首先对原始reads 进行过滤,去除reads中的3’接头序列、由于接头自连等原因导致没有插入片段的reads、剪切3’端测序质量较低的碱基(质量值小于20)、含未知碱基N 的reads、长度过短的reads(<18 nt)和长度过长的reads(>32 nt)以及含有poly A 尾的reads。计算数据的Q20、Q30和GC 含量,以便进一步分析。然后,将高质量的sRNA 标记通过Bowtie 无错配的方式对比到油菜参考序列上,分析其在油菜参考基因组(中双11)上的表达和分布。
1.5 miRNA分类,靶基因预测及其差异表达分析
对过滤后的reads 进行分类,分析sRNA 数据的组成,选择筛选18~32 nt的reads 作为后续分析的有效性数据,并进行该区间sRNA 长度分布的统计[31]。参照miRBase 20.0,使用mirdeep2 和srna-tolls-cli获得潜在的miRNA 并绘制二级结构。使用自定义脚本获得miRNA 计数和碱基偏置。整合miREvo 和miRdeep2,通过探索不加注释的小RNA 标签的二级结构、Dicer 裂解位点和最小自由能,预测新的miRNA。psRobot 中通过psRobot_tar 对miRNA 的靶基因进行预测。miRNA 表达水平通过每百万转录量(TPM)来估计。使用DESeq R 软件包(1.8.3)将校正后的p值设为0.05 作为鉴别差异表达miRNA 的阈值。差异表达的miRNA 及其靶基因通过Wallenius non-central hyper-geometric distribution 进行GO分析。利用KEGG(http://www. kegg. jp/)数据库对miRNA 及其靶基因的代谢途径富集进行分析[30,32],并使用KOBAS 来检测KEGG 通路中miRNA 及其靶基因的富集情况。
2 结果与分析
2.1 sRNA序列分析
为了探究油菜盐胁迫条件下miRNA 的调控机制,提取了在200 mmol·L-1盐浓度下生长的油菜总RNA 并进行了小RNA 测序。使用Illumina 测序技术,在对照组地上部(control shoot,CS)、处理组地上部(treatment shoot,TS)、对照组根部(control root,CR)和处理组根部(treatment root, TR)小RNA 文库中分别获得约35 704 556、40 977 538、45 958 866 和41 322 651 条raw reads(表1)。对raw reads 进行过滤后,单个样品的clean reads 比率、Q20、Q30以及GC含量分别在90%、98%以及39%以上。总共获得了164 070 484个clean reads(表2)。

表1 油菜文库小RNA测序结果Table 1 Results of sRNA sequencing of B.napus library
在本研究的转录组中,小RNA 序列长度在18~32 nt 范围内,最丰富的是21~24 nt(图1A)。这些小RNA 由已知的miRNA、预测的miRNA、核糖体RNA(rRNAs)、转移核糖核酸(tRNAs)、核小RNA(sn-RNAs)、核仁小RNA(snoRNAs)、反作用小干扰RNA(TAS)和未知的片段组成(表2)。本研究发现,在总的小RNA 中有6.68%~14.34%是已知的miRNA,而73.91%~80.86%是未知的miRNA。未知的miRNA数量如此之多,表明在油菜中仍有很多的miRNA 有待鉴定。从地上部和根中分别鉴定出2897 和2435个miRNA(图1B),其中,不论在地上部还是根中,已知的miRNA 数量总是少于预测的新的miRNA。最丰富的miRNA 序列长度为24 nt(12.91%),其次是21 nt(8.40%)和23 nt(8.85%)(图1A),这些长度与被子植物中典型的miRNA 分布相似。样品中长度为24 nt的miRNA 首位碱基以“U”为主,这符合miRNA首位碱基偏向“U”的特征(图1C,D)。
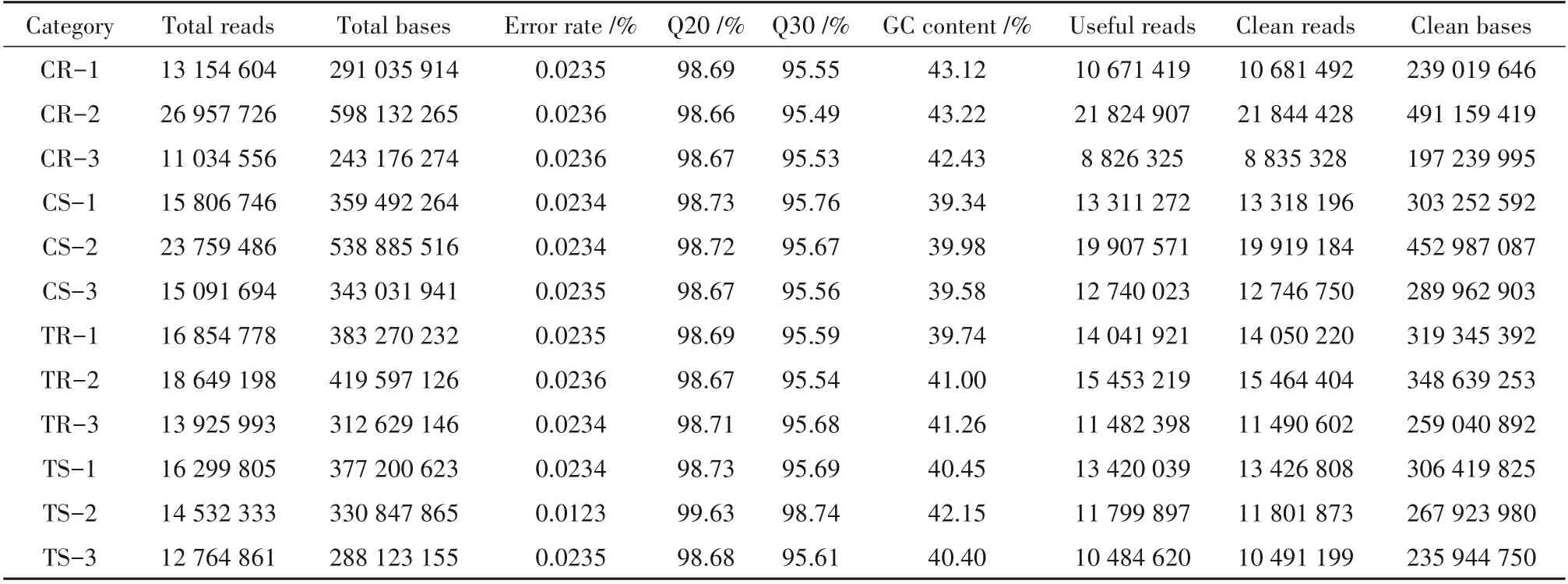
表2 小RNA注释分类Table 2 Annotation classification of small RNA

图1 不同处理条件下油菜地上部和根中miRNA碱基长度分布(A)、数量(B)、首位碱基偏好性(C)、各位点碱基偏好性(D)Fig.1 Base length distribution of miRNAs(A),numbers of miRNAs(B),preference distribution of the first base of different length miRNAs(C),base preference distribution of miRNAs(D)in shoots and roots of B.napus under different treatments
2.2 油菜中保守miRNA及新miRNA的鉴定
为了鉴定油菜地上部和根中的保守miRNA,将小RNA 序列与miRBase 数据库中已知的植物miRNA序列进行比较。通过BLASTN搜索和进一步的序列分析,共发现了26 个miRNA 家族中的91 个miRNA。最大的miRNA 家族miR169 有10 个成员(图2)。从文库中可以看出,这些miRNA 家族的丰度存在明显差异。为了比较它们表达水平,计算了12 个文库中每个miRNA 的每百万读次的平均转录量(TPM)[33,34]。TPM 标准化后,bna-miR395f 的表达水平最低,而bna-miR171f的相对表达水平最高,其次是bna-miR166b 和bna-miR169j。为了识别新的miRNA,使用miREvo 和miRDeep2 程序预测二级结构和Dicer 裂解位点,并测量最小自由能。总共从12 个文库中预测了2116 个新miRNA。新的成熟miRNA的长度为18~24 nt,约60%为24 nt。
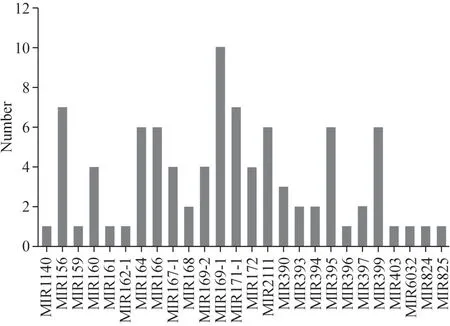
图2 油菜地上部和根中每个miRNA家族的保守miRNA数量Fig.2 Number of conserved miRNAs of each miRNA family in shoots and roots of B.napus
2.3 不同盐处理下油菜miRNA的差异表达分析
为了确定miRNA 的表达与盐胁迫的关系,对miRNA 的表达量进行了分析,筛选出了表达量相对较高的15个miRNA,并分析它们在油菜地上部和根中的表达情况(图3A)。使用fold-change 进行计算,将处理组地上部和根部中miRNA 的表达与对照组地上部和根部中miRNA 表达进行比较,规定|fold change|≥2 且p<0.05 条件下的miRNA 在盐胁迫条件下存在表达差异[35,36]。
与CS文库中的表达相比,TS文库中有9个miRNA 表达下调,6 个miRNA 表达上调(图3B),与CR文库中的表达相比,TR 文库中有25 个miRNA 表达下调,13 个miRNA 表达上调(图3C)。这些差异表明,盐胁迫影响着这些miRNA的表达。
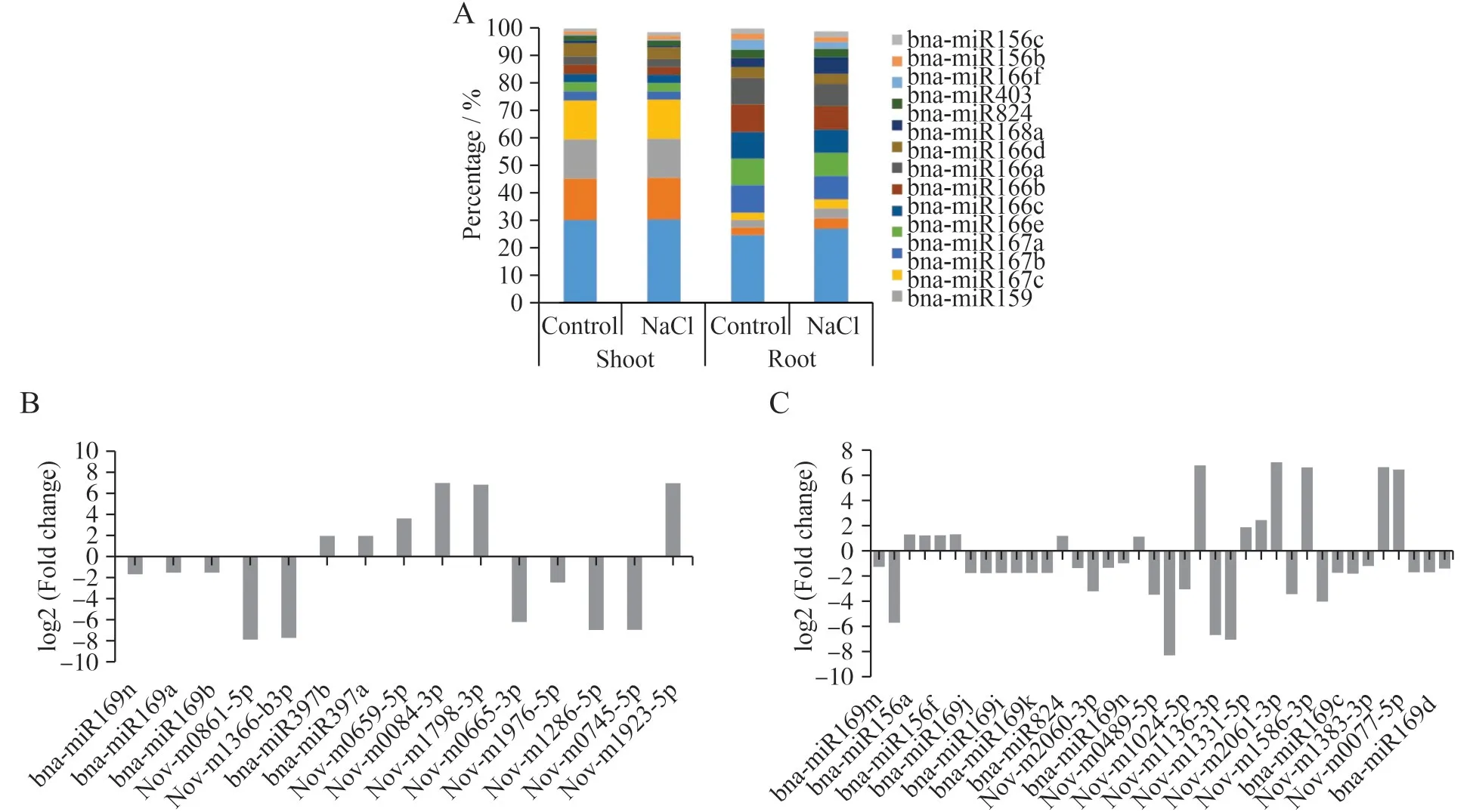
图3 不同处理下差异表达量前15的miRNA在地上部和根中所占的比例(A),地上部处理组和对照组之间差异表达miRNA的表达量(B)和根部处理组和对照组之间差异表达miRNA的表达量(C)Fig.3 Proportion of miRNA in shoots and roots of the top 15 with differential expression under different treatments(A),differential expressions of miRNA between treatment shoot and control shoot(B),and differential expressions of miRNA between control root and treatment root(C)
2.4 差异表达miRNA的靶基因数量分析
为了进一步解释miRNA 在盐胁迫反应中的作用,我们对miRNA 的靶基因进行了分析,并统计了每个差异表达miRNA 的靶基因数目,其中靶基因数目最多的是bna-miR156d,bna-miR156e和bnamiR156f,分别都含有85个受调控的靶基因(图4)。
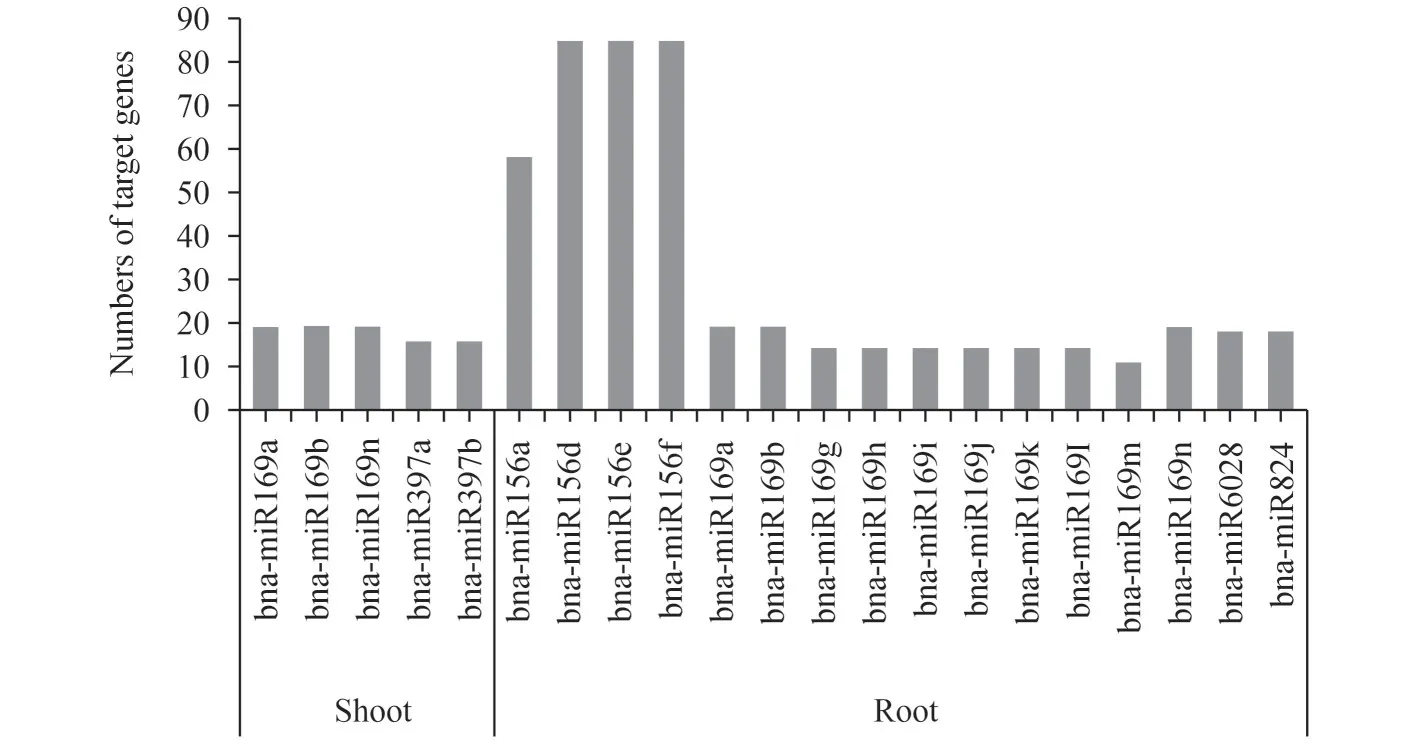
图4 差异表达miRNA的靶基因数量Fig.4 Numbers of target genes of miRNAs
2.5 miRNA靶基因GO注释和KEGG分析
在CS 和TS 小RNA 文库中,鉴定出17 个差异表达的miRNA,共含有93 个靶基因;其中已知miRNA的靶基因有89 个,新miRNA 的靶基因有4 个。在CR 和TR 小RNA 文库中,鉴定出38 个差异表达的miRNA,共含有684个预测靶基因,其中已知miRNA的靶基因有544 个,新miRNA 的靶基因有140 个。大多数的预测靶基因编码的一些与胁迫相关的转录因子,其中包括SPL、LAC、UBC、NFY以及其它蛋白质如AGO2、超氧化物歧化酶等。为了了解miRNA 的生物学功能,所有推测的靶基因都通过Blast2GO 软件进行了GO 功能分类,以进一步探讨miRNA在响应盐胁迫中的作用。
已知miRNA 和新miRNA 的靶基因被分为三个GO类别:细胞组分(CC)、生物学过程(BP)和分子功能(MF)。图5(A)是在CS 和TS 小RNA 文库中鉴定到的miRNA 靶基因GO 富集图,图5(B)是在CR 和TR 小RNA 文库中鉴定到的miRNA 靶基因GO 富集图。CS 和TS 小RNA 文库中共有93 个靶基因参与了1835 个GO 条目(图5A)。其中BP 中高度富集的GO 条目是细胞过程(cellular process);CC 中高度富集的GO 条目是细胞(cell);MF 中高度富集的GO 条目是结合活性(binding)。CR 和TR 两个小RNA 库共有684 个靶基因参与了4941 生物过程(图5B)。其中BP 中高度富集的GO 条目是细胞过程(cellular process);CC 中高度富集的GO 条目是细胞(cell);MF 中高度富集的GO 条目是结合活性(binding)。四个数据库中都发现了刺激反应(response to stimulus),表明这些靶基因可以在应对盐胁迫中发挥作用。已知KEGG 代谢通路被分为7 大类:代谢(Metabolism)、遗传信息处理(genetic information processing)、环境信息处理(environmental in-formation processing)、细胞过程(cellular processes)、生物体系统(organismal systems)、人类疾病(human diseases)、药物开发(drug development)。图6是差异表达miRNA靶基因参与的pathway通路,其中A图为CS和TS小RNA 文库中鉴定到的差异表达miRNA 靶基因参与的pathway 通路,B 图为CR 和TR 小RNA 文库中鉴定到的差异表达miRNA 靶基因参与的pathway 通路。在CS和TS小RNA文库中的生物体系统(organismal systems)中有18 个靶基因参与了免疫防御系统(immune system)。在CR 和TR 两个小RNA 库在CS 和TS 小RNA 文库中的生物体系统(organismal systems)中有21 个靶基因参与了免疫防御系统(immune system)。上述结果表明这些靶基因可以积极参与油菜盐胁迫抗性反应。
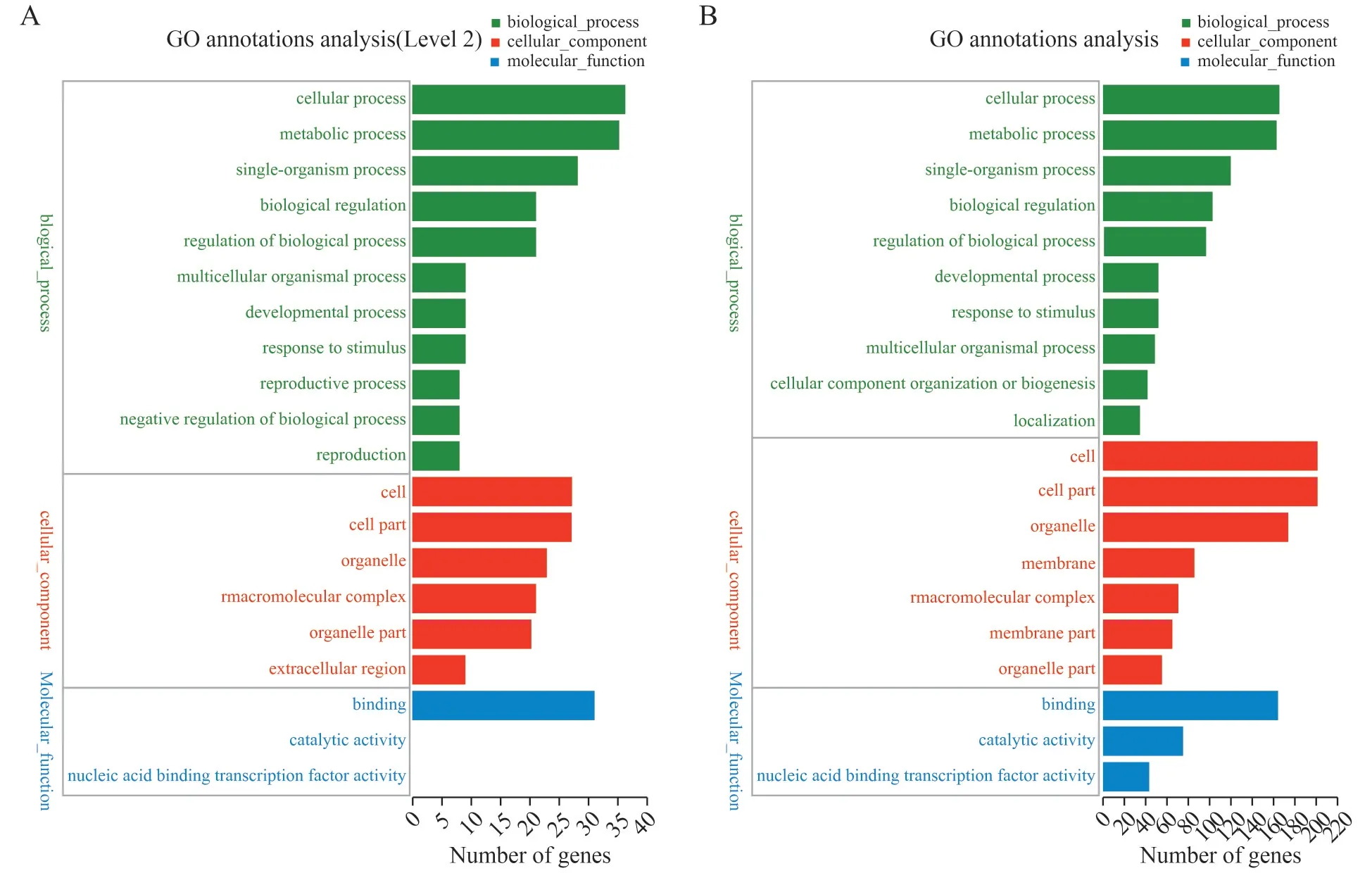
图5 差异表达miRNA靶基因功能分类Fig.5 Functional classification of different expression miRNA targets

图6 差异表达miRNA靶基因参与的pathway通路Fig.6 Pathway involved in differentially expressed miRNAs target genes
2.6 油菜地上部和根中差异表达miRNA的分析
通过分析油菜地上部和根中miRNA 的差异表达谱,图7A 和图7B分别是盐胁迫条件下,油菜地上部中上调表达和下调表达的miRNA;图7C 和图7D是盐胁迫条件下,油菜根中上调和下调表达的miRNA。在地上部中,发现NaCl处理后bna-miR397a和bna-miR397b表达显著上调,bna-miR169a、bnamiR169b和bna-miR169n的表达显著下调。在根中,bna-miR156a、bna-miR156d、bna-miR156e、bnamiR156f以及bna-miR824的表达显著上调,而bnamiR169j、bna-miR169i、bna-miR169h、bna-miR169k、bna-miR169g、bna-miR169a和bna-miR169b的表达并没有显著变化,bna-miR169m、bna-miR169n、bnamiR169c、bna-miR169e、bna-miR169f、bna-miR169d和bna-miR169b表达显著下调。
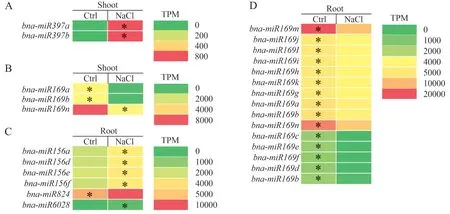
图7 盐胁迫条件下油菜的差异表达miRNAFig.7 Differentially expressed miRNAs in B.napus under salt stress
2.7 靶基因表达分析
前人研究表明有多个miRNA 参与植物盐胁迫反应[23,37~41],分别为miR397-LAC(bna-miR397a、bnamiR397b) 、 miR156-SPL(bna-miR156a、bnamiR156d、bna-miR156f、bna-miR156e)、miR169-NFY(bna-miR169m、bna-miR169g、bna-miR169n)以及miR399-UBC(bna-miR399b)。其中转录因子miR156 的靶基因是SPL[37],在本研究中(图8)发现miR156在根中表达量上调,而在本课题组前期转录组测序结果中发现SPL15表达量下调[30],因此miR156 可能通过调控SPL15来响应盐胁迫,从而缓解盐胁迫给植物带来的损伤。LAC是miR397 的靶基因,其可以通过调控木质素含量来响应外界非生物胁迫,差异表达分析显示miR397在地上部中表达上调,LAC12在本课题组前期转录组测序结果中表达量下调[30],miRNA397 可能通过调控LAC12的表达,促进原生木质部细胞壁增厚,从而响应盐胁迫。转录因子NFY是miR169的靶基因[42],它是非生物胁迫反应的重要调节因子,在本研究中发现miR169在地上部和根中的表达量都下调,而在本课题组前期转录组测序结果中NFYA5的表达量显著上调[30],miR169 可能通过调控NFYA5的表达参与盐胁迫反应。UBC是miR399 的靶基因,差异表达分析显示miR399在根中表达量下调,而在本课题组前期转录组测序结果中UBC29的表达量显著性上调[30],miR399 可能通过调控UBC29的表达从而参与异常蛋白质降解过程,提高作物的耐盐性。

图8 盐胁迫条件下对miRNA397靶基因LAC的影响(A)、miRNA156靶基因SPL的影响(B)、miRNA169靶基因NFY的影响(C)、miRNA399靶基因UBC的影响(D)Fig.8 Impact on miRNA397 target gene LAC(A),miRNA156 target gene SPL(B),miRNA169 target gene NFY(C),and miRNA399 target gene UBC(D)under salt stress
3 讨论
盐渍土使植物暴露在渗透胁迫下,渗透胁迫对植物最重要的后果之一是产生大量活性氧,随后是氧化损伤,例如蛋白质、脂质、色素和DNA 的降解。生长在含盐条件下的植物吸收有害离子,尤其是钠离子和氯离子,其大量积累对细胞有毒,并加剧渗透胁迫。这些离子破坏膜完整性、细胞代谢、酶结构、细胞生长和光合作用[43]。近年来,miRNA 在基因表达调控中的作用越来越清晰,许多miRNA 如miR156、miR169、miR393、miR396、miR397 及其靶基因[44]等在拟南芥[13]、大豆[45]、大麦[46]等多种植物的盐胁迫响应中发挥重要作用。
前人研究表明油菜中有56 个基因编码假定的转录因子在非生物胁迫下会发生改变[47]。在这些基因中,已经显示在盐胁迫下上调超过5 倍的基因来自AP2-EREBP家族(ATERF11,CBF4/DREB1D,CBF1/DREB1B, ATERF4/RAP2.5),碱性-螺旋-环-螺旋(bHLH)家族(AtbHLH17),碱性区亮氨酸拉链(bZIP)家族(AtbZIP55/GBF3),C2H2家族(ZAT10、ZAT12、RHL41、ZAT6和ZAT102/RHL41) ,NAC 家 族(ANAC036、ANAC029/ATNAP、ANAC055/ATNAC3、ANAC047、ANAC072/RD26、ANAC002/ATAF1、ANAC019和ANAC032),WRKY家族(ATWRKY53,ATWRKY40和ATWRKY33),热激家族(ATHSFA1E)以及Homeobox家族(ATHB-7)。此后,由转录因子和其他蛋白质组成的复杂基因调控网络控制着许多基因的表达。但是在本研究中,发现SPL15在盐胁迫条件下表达量下调,而LAC12、NFYA5和UBC29在盐胁迫条件下表达量显著上调。遗憾的是,目前对油菜盐度胁迫下miRNA及其作用靶点的研究还不够全面。
为了鉴定miRNA 及其靶基因在油菜盐胁迫响应中发挥的重要作用,在本研究中,鉴定了来自油菜的数百万个小RNA 序列,以了解盐胁迫条件下miRNA 对植物生长发育的影响。分别在200 mmol·L-1NaCl(T)和无盐处理对照(C)条件下培养油菜品种中双11,取其地上部(S)和根(R)构建了12 个小RNA 文库,并用Illumina 基因组分析仪对其进行测序,从CS、TS、CR 和TR 库中共得到198 831 780 个raw reads。进一步对全基因组的miRNA 进行差异表达分析,一共鉴定到了26 个差异表达的miRNA。这些差异表达的miRNA 导致了下游多个基因的表达发生显著变化,并且植物对盐胁迫的响应伴随着一系列广泛的细胞内过程,包括信号感知、信号转导、转录和蛋白质生物合成等。这说明不同的植物通过多种miRNA 介导的调控策略来响应胁迫[48],例如miR159、miR393、miR396 和miR398 等与盐胁迫有关[49~52]。
在本研究中鉴定出miR156-SPL15、miR397-LAC12、miR169-NFYA5和miR399-UBC29在盐胁迫条件下发挥作用。有研究表明miR156 及其靶基因SPL在植物非生物胁迫的响应中发挥着重要的作用[53],在本研究中,miR156 特异性的在根中上调表达,这一结果与前人的研究结果形成对比,即拟南芥在盐胁迫下miR156 的表达下调[21],并通过调控其靶基因SPL15参与到盐胁迫响应中;苹果在盐胁迫条件下miR156的表达下调,其靶基因SPL13通过激活WRKY100,促进与抗氧化剂生物合成,糖代谢和脯氨酸生物合成有关基因的转录,进一步维持ROS和渗透平衡,从而增强盐胁迫的耐受性[21]。这表明不同植物对盐胁迫的响应存在差异[54]。miR397 及其靶基因LAC可以通过调控木质素含量来参与非生物胁迫(58),在本研究中,鉴定到LAC12是miR397a 的靶基因,该基因可以导致木质素含量升高,进一步改变Na+迁移途径从而提高植物对盐胁迫的抗性。已有研究表明,番茄和玉米根系中高浓度的NaCl均能提高LAC的转录水平,并指出盐胁迫下根系转录水平的提高可能是植物的普遍响应。因此,LAC可能在植物根系适应盐胁迫过程中发挥重要作用[55,56]。越来越多的证据表明NFY是非生物胁迫反应的重要调节因子[16,17,43,57~61],并受到miR169 的调控。在本研究中miR169 家族成员miR169g 和miR169n 在根中的表达量下调,这和前人之前的研究一致,但是过表达miR396c 的水稻和拟南芥对盐胁迫抗性都降低,而过表达miR396a/b 转基因植株的耐盐性提高。这表明同一个miRNA 家族的不同成员在不同的植物中采用不同的方式参与盐胁迫响应[60]。然而,盐胁迫条件下油菜中NFY的作用还没有被详细研究。前人研究表明UBC[42,62]定位在内质网上,是miR399 的靶基因,其表达受高盐度或ABA 诱导并参与异常蛋白质的降解过程[42],且在盐胁迫条件下UBC转录水平升高,通过调控泛素化途径参与植物盐胁迫抗性反应。这表明,植物在盐胁迫反应中可能存在着共同的调控机制。这些结果为进一步研究作物抵抗盐胁迫的分子机制提供了理论依据。
4 结论
植物的耐盐过程是复杂的,由多种因素决定,miRNA 的参与进一步增加了这一过程的复杂性。本研究通过油菜miRNA 测序以及转录组组测序技术发掘盐胁迫响应相关的miRNA及其核心靶基因,丰富了油菜地上部和根部miRNA 的情况。共筛选出26 个与盐胁迫应激反应相关的候选miRNA,并且同一家族的不同miRNA 表达模式有所不同。差异表达的miRNA 的靶基因显著富集的通路不同。这些miRNA 的鉴定及其功能意义的阐明拓宽了我们对盐胁迫反应中转录后基因调控的理解,为探索油菜耐盐机制提供重要的参考。
