甘蓝型油菜分枝角度QTL定位及候选基因分析
2022-03-16赵小珍赵卫国张春余坤江彭门路陈锋张维孙程明李保军王灏王晓东张洁夫
赵小珍,赵卫国,张春,余坤江,彭门路,陈锋,张维,孙程明,李保军,王灏,王晓东*,张洁夫*
(1.南京农业大学作物遗传与种质创新国家重点实验室,江苏 省现代作物生产协同创新中心,江苏 南京,210095;2.江苏省农业科学院经济作物研究所,农业 农村部长江下游棉花与油菜重点实验室,江苏 南京,210014;3.陕西省杂交油菜研究中心,陕西 杨凌,712100)
油菜是我国第一大油料作物,菜籽油占国产食用植物油的47%以上[1]。随着生活水平的提高,人们对油脂的需求不断增强,进一步提高油菜产量变得尤为重要。分枝角度指的是有效分枝与主茎形成的夹角,是影响油菜产量的重要株型性状[2]。研究表明,适度紧凑的株型有利于油菜中下部的通风透光、降低病虫害、提高群体光能利用率、提高叶面积系数和抗倒伏性,从而实现高产[3,4]。
植物分枝角度的形成是其感受器官受到重力刺激后,迫使各器官与重力形成最适角度的过程[5],重力反应的异常会导致分枝角度的异常[6]。水稻(Oryza sativaL.)的分蘖角度[7]及玉米(Zea maysL.)的茎叶夹角[8]等均受到重力反应影响。当前,人们普遍认为重力反应分为4 个步骤,即重力信号的感受、转导、重力信号造成的生长素差异性分布及生长角度的形成[9]。目前就植物重力感受机制得到普遍认可的是淀粉-平衡石假说[10],该假说认为平衡石细胞的淀粉体能够感受重力刺激的改变并作出应激反应[11],这里的平衡石细胞是一种富含淀粉体的高度极化细胞。植物在感受到重力刺激后随之引起体内信号转导,研究表明,Ca2+、InsP3(1,4,5-三磷酸肌醇)、pH 等信号分子独自或协作参与调控了该过程[12]。最后,通过将不同信号分子传递到反应器官来激活下游底物发挥功能,进而完成重力反应。Cholodny-Went 假说[13]认为重力刺激引起生长素不对称性分布,从而引起重力反应部位差异性生长,其在拟南芥重力实验中得到了证实[14]。除了生长素,乙烯[15]、油菜素内酯[16]等激素也参与了重力反应调控。
1938 年水稻lazy1(la1)突变体的研究首次分析了其分枝(分蘖)角度调控机理,该突变体重力反应缺陷导致后期分蘖平卧生长[17];2007年la1突变体的基因LAZY1被成功克隆[18]。除了LAZY1基因,控制水稻分枝(分蘖)角度的基因TAC1(TILLER ANGLE CONTROL 1)[19]、PROG1(PROSTRATE GROWTH 1)[20]、LPA1(LOOSE PLANT ARCHITECTURE 1)[21]及PAY1(PLANT ARCHITECTURE AND YIELD 1)[22]等相继被克隆出来。基因LAZY1在玉米和拟南芥中的同源基因ZmLA1[8]和AtLAZY1[23],基因TAC1在玉米和桃树同源拷贝ZmTAC1[24]和PpeTAC1[25]等,也相继被克隆。截止目前,部分分枝角度、分蘖角度、茎叶夹角候选基因已在水稻、拟南芥和玉米中被克隆和鉴定,但在油菜上研究较少,尚未有基因克隆的报道。
近年来,QTL 定位和GWAS 分析被广泛应用于识别复杂的数量性状位点,并成功应用于甘蓝型油菜分枝角度性状的研究中。张倩[26]利用F2群体定位到1 个分枝角度的QTL,位于LG1 连锁群,解释14.16%的表型变异。Liu 等[27]利用包含143 份株系的自然群体进行分枝角度的全基因组关联分析,在A2、A3、A7、C3、C5 和C7 染色体上定位到25 个显著QTL,并在A3 和C3 关联位点附近找到了分枝角度基因LAZY1的同源拷贝。Sun 等[28]利用从国内油菜主产区及欧洲等其它国家收集来的520份自然群体进行全基因组关联分析,定位到56 个QTL,共解释表型变异的51.1%,并在这些区域预测了多个候选基因,包括LAZY1、SGR2(SHOOT GRAVITROPISM 2)、PIN3(PIN-FORMED 3)、TIR1(TRANSPORT INHIBITOR RESPONSE 1)等。Wang 等[29]利用BSA 的方法结合二代测序技术对双亲分枝角度差异较大的F2群体进行分枝角度精细定位,最终在A6染色体上检测到一个分枝角度主效QTL,解释17.17%的表型变异,并在该QTL 的置信区间内筛选到生长素合成相关基因BnaYUCCA6为候选基因。Li 等[30]利用60K SNP 芯片对472 份油菜核心种质进行全基因组关联分析,在6 个不同环境下进行分枝角度表型鉴定,MLM 和MRMLM 模型各检测出46 个和38 个显著相关的位点,分别解释表型变异的62.2%和66.2%,并根据同源拟南芥的注释,首次发现了定位在油菜中的分枝角度基因TAC1和SGR1等。Shen等[31]利用60K SNP 芯片对含有208 份株系的DH 群体进行基因分型,构建遗传图谱并进行QTL 定位,共定位到17 个QTL。汪文祥等[32]利用由分枝角度差异显著亲本构建的包含163个株系的DH群体,在2 个环境中共定位到17 个分枝角度QTL,并推测基因VAMP714,TAC1,IAR和ARF16等为油菜分枝角度的候选基因。以上研究初步获得了一些油菜分枝角度性状位点信息,但还远远不能揭示该性状的遗传机制。
前期利用油菜60K SNP 芯片,对包含189 个株系的甘蓝型油菜重组自交系(RIL)群体进行基因分型,构建了高密度SNP 遗传图谱[33]。本研究在此基础上,对RIL群体的分枝角度性状进行考察,结合高密度遗传图谱进行QTL 定位与分析,并在稳定表达的QTL 区间内筛选候选基因,为深入解析甘蓝型油菜分枝角度的遗传机理,精细定位和克隆分枝角度相关基因提供参考。
1 材料与方法
1.1 材料
前期利用父本Holly和母本APL01,构建了一个包含189 个株系的RIL 群体并命名为AH。利用油菜60K SNP芯片,构建了高密度的遗传连锁图谱,其与甘蓝型油菜参考基因组Darmor-bzh 具有良好的共线性[33]。该图谱包含2755 bin(含11 458 个SNP)和57个SSR,覆盖甘蓝型油菜全基因组19个连锁群的2027.53 cM,标记间的平均距离为0.72 cM[33]。利用该群体,前期对无花瓣性状[33]、脂肪酸组分[34]、初花期[35]及耐渍性状[36]进行了QTL 定位及分析。本研究中,AH 群体用于分枝角度的QTL 定位及候选基因分析。
1.2 田间试验
将RIL 群体及其亲本于2015 年9 月至2016 年5月种植于江苏南京和陕西杨凌两个试验点,两个环境的实验分别记录为15NJ 和15YL。田间播种均采用随机区组设计,每个小区每个株系种植2行,设置两次重复,行间距40 cm,单株间平均间距20 cm。试验材料的种植、管理和收获等均按照当地大田生产管理方式进行。
1.3 表型考察
参考孙程明[37]对分枝角度测量的方法,本实验在油菜成熟后剪取带有油菜顶部5个分枝的茎段作为单株的分枝角度,并将其装袋编号。在15NJ 和15YL 两试验点,每个株系选取5 株进行考察,每个环境包括两次重复。取样回来的样品将其放置在样品置放板上,利用数码相机拍下能清晰呈现主茎与每一个分枝节点夹角的部分,每株5个分枝共得5张照片,利用计算机Photoshop 软件中的直线工具和角度工具测量照片中主茎和分枝之间的夹角,并将其记录到Microsoft Excel文档中,两个重复的平均值作为株系在一个环境的表型值。利用R 软件[38]计算其广义遗传力。
1.4 QTL定位分析
使用软件Windows QTL Cartographer 2.5[39]的复合区间作图(composite interval mapping,CIM)法,对2 个环境下分枝角度进行QTL 的检测。窗口大小设为10 cM,选用2 cM 的步长(walking speed),同时设置5个标记作为辅助因子。每个性状的显著性水平设为0.01,进行1000 次排列检验(permutation test),以此来判断是否存在QTL,并将其称为identified QTL。对于在2 个环境下都有表达且具有重叠置信区间(confidence intervals,CIs)的QTL 进行整合并称为一致性QTL(consensus QTL);对于在2 个环境中只检测到一次的QTL同样称为一致性QTL。QTL的整合应用软件BioMercator V4.2[40]元分析(metaanalysis)法实现。参考McCouch 等[41]和Wang 等[42]对QTL的命名的介绍,本研究对QTL命名,以检测到的QTLidentified QTL 英文缩写iq 作为开头,依次加上环境缩写(15NJ、15YL)和连锁群编号(A1-A10、C1-C9)。若在同一个连锁群上出现了不止一个QTL,就依据顺序对其进行编号。如QTLiq15NJ. C4-1,表示2015 年南京环境中检测到的C4 染色体上的第一个QTL。对于经元分析整合后的一致性QTL,其命名法则类似上述identified QTL 命名原则,以一致性QTL 英文缩写cq 开头,再依次加上分枝角度性状缩写(branch angle,BA)和连锁群编号。若在同一个连锁群上出现了不止一个一致性QTL,就依据顺序对其进行编号。例如QTLcqBA. C4-2表示C4 染色体上分枝角度的第二个一致性QTL。
1.5 候选基因预测
候选基因的定位已经实现了从区间定位到通过分析基因组数据库信息筛选候选基因的阶段[43]。本研究利用Illumina HiSeq 2500 平台(Illumina,Inc;San Diego,CA,USA),对双亲APL01 和Holly 基因组进行了重测序,测序深度为30×,共获得约249.95 Mb Clean Reads,74.89 Gb Clean Data,Q30平均达到92.00%。
候选基因的预测分为3步:首先,利用重测序的结果,在两亲本的全基因组内鉴定变异位点;其次,利用遗传图谱上SNP 的探针序列,通过Blast(E 值≤1e-10)将QTL 区间两侧的标记映射到参考基因组Darmor-bzh 的物理位置[44],并根据第一步的结果,查找QTL 区间内的SNP 位点和InDel 位点及造成非同义突变的SNP 和发生移码突变的InDel;第三,以非同义突变的SNP和移码突变的InDel为目标,根据拟南芥基因的功能注释筛选分枝角度可能的候选基因。依据对现有分枝角度相关文献的参考,分枝角度候选基因的选择偏向于功能注释中关于向重力性,生长素的生物合成、运输及代谢相关基因。测序工作由诺禾致源生物信息科技公司(北京)完成,数据分析工作由中国农业科学院油料作物研究所油料作物分子改良理论与技术团队完成。
2 结果与分析
2.1 RIL群体及其双亲分枝角度表型分析
对RIL群体及亲本的分枝角度进行了两个环境的考察(图1、表1)。在15NJ 和15YL 2 个试验点,APL01 的分枝角度分别为25.2°和32.8°,Holly 的分枝角度分别为33.5°和41.9°,两亲本的分枝角度在两个环境中均存在极显著差异(P<0.01)(表1)。RIL 群体的分枝角度在两个环境中具有广泛的变异,平均变异幅度各为24.74°~37.76°和31.05°~46.67°。单个环境的最大值大于高值亲本,最小值小于低值亲本,表现出超亲分离现象(图1、表1),暗示增效等位基因在双亲中都有分布。两环境下的分枝角度表现出连续性分布,表现出明显的数量性状特征,适合用于QTL检测(图1)。R软件计算出分枝角度在2 个环境下的广义遗传力为86%,说明油菜的分枝角度主要受遗传因素控制。
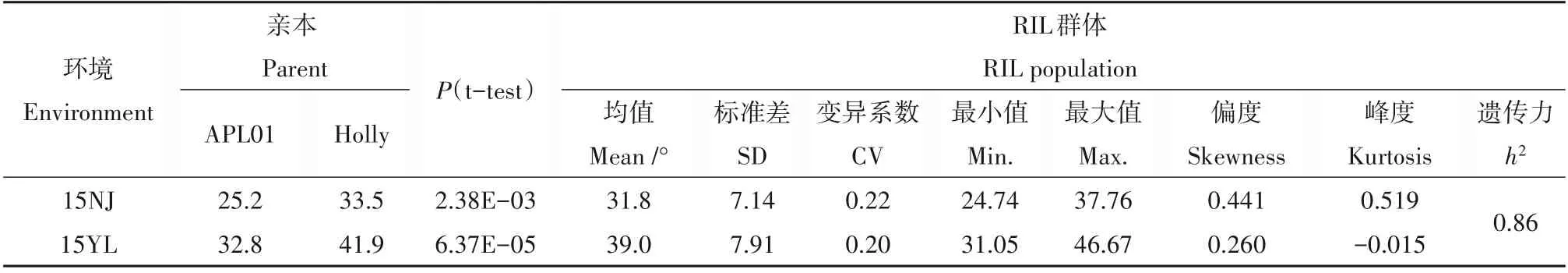
表1 两环境下亲本及RIL群体分枝角度性状的统计分析Table 1 Statistical analysis of branch angle of parents and RIL populations under 2 environments

图1 AH RIL群体在两环境的分枝角度表型频率分布Fig.1 Phenotypic frequency distributions of branch angle in AH RIL population under 2 environments
2.2 分枝角度QTL检测
利用软件Windows QTL Cartographer 2.5 中的复合区间作图法进行QTL 定位,在15NJ 和15YL 两环境中共检测到11 个identified QTL,分布在A9、C3、C4 和C7 染色体上(表2),单个QTL 解释表型变异的4.05%~9.60%,加性效应范围为-1.52~1.96。其中有7 个identified QTL 分布在C4 染色体,每个QTL 对表型变异的贡献率在4.60%~9.60%。利用软件BioMercator V4.2 将其中5 个具有重叠置信区间QTL 整合成2 个一致性QTLcqBA. C4-2和cqBA.C4-3(表2),这两个QTL 均在15NJ 和15YL 被反复检测到。其它6 个QTL 只在1 个环境中检测到(表2),表现出环境特异性,说明环境对于这些QTL 位点影响的程度较大,利用它们对分枝角度进行遗传改良时,环境因素不可忽略。经过元分析后,本研究共获得8个分枝角度的一致性QTL,其中7个一致性QTL 的加性效应为正值,说明其增效基因来自于母本APL01;另一个一致性QTLcqBA. A9,具有-1.52 的负加性效应,表明其增效基因来自于父本Holly(表2)。

表2 甘蓝型油菜分枝角度QTL结果Table 2 QTL results for branch angle of Brassica napus
2.3 与已报道的油菜分枝角度QTL的比较
油菜中分枝角度的文献报道较少,对已经报道的分枝角度性状定位结果进行整理[26~32],发现除了C1染色体外,分枝角度位点在其它染色体上都有分布,其中A3,A7 和C3 染色体分布数目最多,各有10个以上位点,其余染色体各有2~7 个位点。为比较本研究定位的8 个QTL 和前人报道的QTL 是一致,将其连锁/关联的标记/探针序列比对到参考基因组Darmor-bzh 以确定QTL 的物理位置。本研究在A9染色体上定位到一个QTLcqBA.A9,其位于22.15~23.05 Mb 区间(表2),通过与已研究报道的QTL[28,30]比较,置信区间没有重叠(表3);在C3 染色体上也定位到一个QTLcqBA.C3,其位于4.21~4.87 Mb 区间(表2),通过与已研究报道的QTL[27,28,30~32]比较,置信区间没有重叠(表3);在C4 染色体上定位到四个QTLcqBA. C4-1(0.082~1.64 Mb),cqBA. C4-2(10.32~14.3 Mb),cqBA.C4-3(44.39~44.46 Mb)和cqBA. C4-4(44.74~46.33 Mb)(表2)。经比较,除cqBA. C4-1与汪文祥等[32]的区间有重叠外,其它3个QTL 与前人的定位结果不同[28,30,32](表3);在C7 染色体上定位到两个QTLcqBA. C7-1(5.73~5.86 Mb)和cqBA.C7-2(30.72~32.18 Mb)与前人的定位结果均不同[27,28](表3)。因此,本研究中定位的8 个QTL,除cqBA.C4-1外,其它7 个QTL 与前人的定位结果均不同,推测为新的控制油菜分枝夹角的QTL位点。
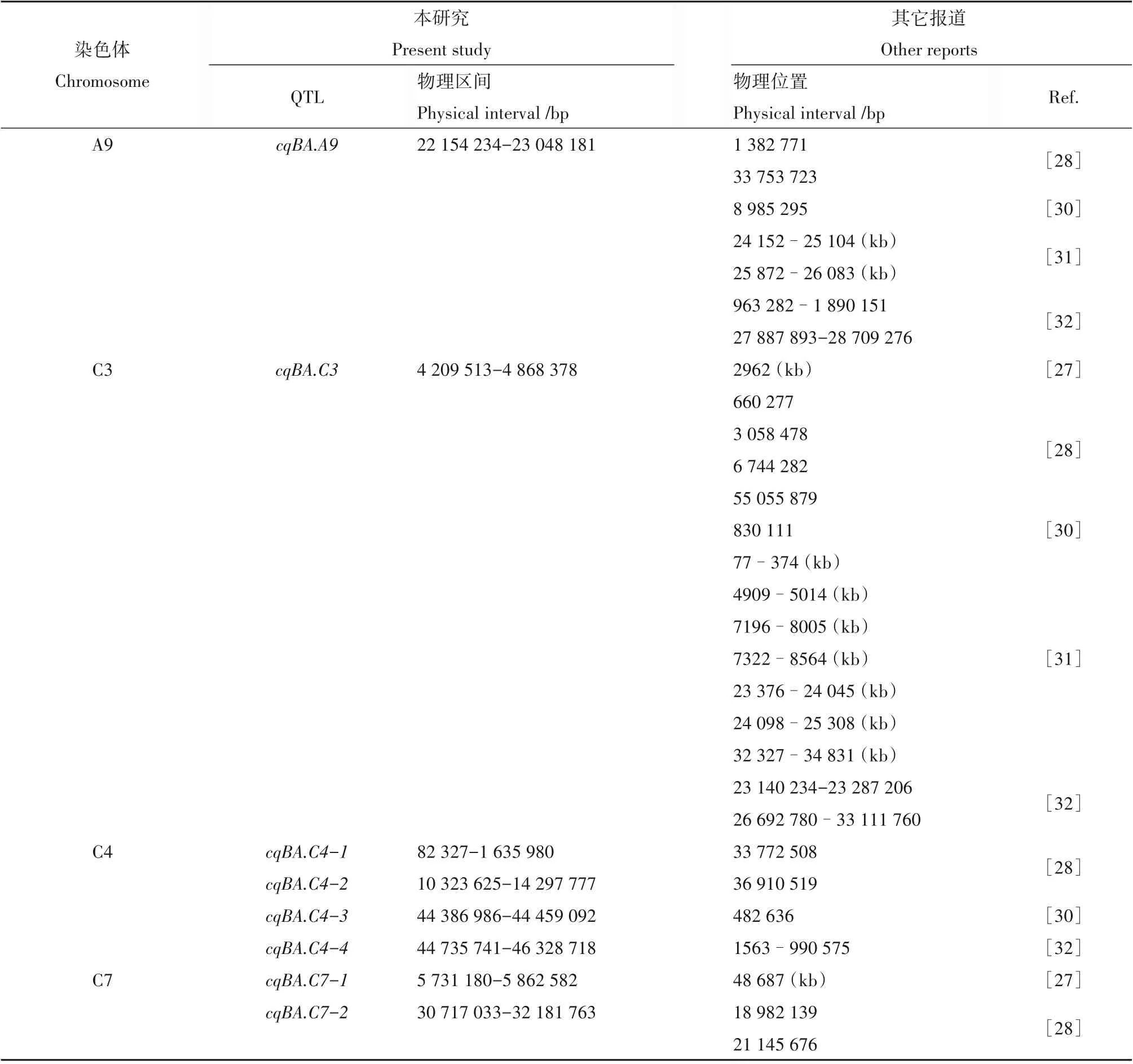
表3 本研究与前人报道的分枝角度QTL比较Table 3 Comparison of the QTL for branch angle from present and previous reports
2.4 分枝角度候选基因的预测
QTLcqBA. C4-2和cqBA. C4-3能在2 个环境中稳定表达(表2),表明这2个QTL区间内可能存在控制甘蓝型油菜分枝角度的基因。这2个QTL均位于C4 连锁群上,对应物理图谱上的位置为10.3~14.3 Mb 和44.4~44.5 Mb。根据Holly 和APL01 的重测序结果,在QTLcqBA.C4-2区间内共发现有7226 个SNP 和829 个InDel,其中7226 个SNP 中包含220 个非同义突变和5 个stopgain 突变,829 个InDel 中有15个发生移码突变,这些突变位点共对应55个基因(附表1,见首页OSID 码);在QTLcqBA. C4-3区间内共发现有416 个SNP 和65 个InDel,其中416 个SNP中包含50个非同义突变,65个InDel中有1个发生移码突变,这些突变位点共对应15 个基因(附表2,见首页OSID 码)。在此基础上,以这70 个基因为目标,借助拟南芥同源基因的功能注释,共筛选到4个分枝角度相关的候选基因,BnaC04g13100D,BnaC04g15900D,BnaC04g16280D和BnaC04g44330D(表4)。

表4 QTL cqBA.C4-2和cqBA.C4-3置信区间获得的候选基因Table 4 Candidate genes in QTL confidence interval of cqBA.C4-2 and cqBA.C4-3
在QTLcqBA.C4-2的置信区间内(10 323 625~14 297 777 bp),共筛选到3 个油菜分枝角度相关基因 ,BnaC04g13100D(AT2G31500,CPK24) 、BnaC04g15900D(AT2G28350,ARF10)和BnaC04g16 280D(AT2G28070,ABCG3)。根据拟南芥基因的功能注释,CPK24(BnaC04g13100D)为钙依赖性蛋白激酶编码基因,CPK基因广泛分布在植物体的不同组织,其产物CPK 蛋白通过解读由各种发育和环境刺激触发的Ca2+信号来参与调节植物的生长[45]。Ca2+作为重要的信号物质,在植物的生长发育中起着关键作用,已有研究表明,Ca2+参与调控植物的重力反应[12],当重力刺激改变时,会导致细胞内Ca2+外流,这些外流Ca2+被CPK 和CaM(钙调素)等感知,并将信号传递到下游效应器,最终导致植物重力响应[46],其中CPK 参与了重力信号的传递[47]。推测钙依赖性蛋白激酶成员基因CPK24(BnaC04g13100D)可能通过参与重力信号的传递来调控分枝角度。Roychoudhry 等[48]研究发现植物非垂直分枝的生长不仅仅由重力性因子决定,提出了反重力性抵消因子(antigravitropic offset, AGO),其与重力性因子共同调控侧枝的生长角度。这种反重力抵消机制发挥作用依赖于生长素转运,拟南芥生长素缺陷突变体,在生长素信号恢复之后,分枝角度也随之减小,生长素通过一个复杂的调控网络参与调控植物分枝角度的形成[49]。本研究在QTLcqBA.C4-2置信区间内筛选到生长素响应因子(auxin response factor,ARF)中的ARF10(BnaC04g15900D),该基因也存在于前人定位的分枝角度区间内[28]。基因ABCG3(BnaC04g16280D)为ABC 转运蛋白(ATP-binding cassette transporter)家族一员,ABC 转运蛋白是植物界普遍存在的一类跨膜运输蛋白,其通过消耗ATP产生的能量参与生物活动。Sun 等[28]和汪文祥等[32]在其置信区间相继筛选出多个ABC 转运家族为分枝角度基因,推测本研究ABC 转运家族基因ABCG3可能为分枝角度候选基因。在QTLcqBA.C4-3的置信区间内(44 386 986~44 459 092 bp)筛选到1 个油菜分枝角度相关基因BnaC04g44330D(AT2G3399 0,IQD9),是IQ67结构域家族的成员,该家族成员只与CaM 特异性结合。CaM 作为Ca2+另一个重要的传感蛋白,同样通过传递钙信号,继而调控植物的生长发育。Bürstenbinder 等[50]认为多个IQD基因为ARF 的潜在靶标,其IQD 蛋白作为钙信号和生长素信号的枢纽来调控植物的形态发生。因此推测IQD9可能参与调控分枝角度。
3 讨论
分枝角度是决定作物株型的重要因素,对于作物合理密植及机械化收获有重要意义。迄今为止,在水稻(分蘖角度)、玉米(茎叶夹角)和拟南芥(分枝角度)中已有较多研究,并对相关基因进行了克隆和鉴定,但在油菜上的研究较少且尚处于定位阶段。甘蓝型油菜中,除C1 染色体外,其余染色体均有分枝角度QTL 位点的分布[26~32],其中A3、A7 和C3上的分布数目最多,各有10个以上位点。本研究在两个环境中,共定位到8 个QTL,分布在A9、C3、C4和C7 染色体上。通过和前人研究中报道的分枝角度QTL 的物理位置比较发现,定位的8 个QTL 除cq-BA. C4-1外,其它7 个QTL 与前人报道的油菜分枝角度QTL 均没有重叠,推测是新的油菜分枝角度QTL位点。
本研究对在2 个环境中均稳定表达的QTLcq-BA.C4-2和cqBA.C4-3进行了候选基因的预测,共筛选到4 个油菜分枝角度相关基因,BnaC04g13100 D(CPK24)、BnaC04g15900D(ARF10)、BnaC04g1628 0D(ABCG3)和BnaC04g44330D(IQD9)。CPK24仅依赖于Ca2+信号来调节植物生长发育[51],而重力刺激会引起Ca2+的变化[52],同时有研究表明CPK 参与了重力信号的传递[47]。此外,有报道[53]称CPK 可以通过磷酸化其载体进而影响植物体内生长素水平,如定位于质膜上的AtCRK5(CPK-related kinase 5)通过磷酸化生长素输出载体AtPIN2,导致其生长素水平降低,重力反应延迟。因此,推测重力因素的改变引起Ca2+信号发生变化继而被CPK24识别,通过影响生长素运输进而控制分枝角度的形成。ARF10为生长素响应因子之一,能够与生长素特异性地结合,以此影响植物的分枝。Sun 等[28]、Shen等[31]和汪文祥等[32]也在其置信区间相继筛选出ARF10和ARF16为分枝角度基因。ABCG3为ABC转运家族蛋白一员,通过消耗ATP 产生的能量来进行跨膜运输,并主要运输植物激素。有研究者表明[54],ABCG36 为一种定位于质膜上的ABC 转运蛋白,其突变体pdr8(ABCG36 缺陷型)在生长素前体IBA(吲哚-3-丁酸)转运上表现出缺陷。此外,ABC超家族PDR 转运蛋白直接参与分枝角度的形成[55]。因此,推测ABCG3可能通过影响生长素运输直接或者间接影响油菜分枝角度。IQD9 属于IQD(IQ67-domain containing protein)基因家族蛋白,只与CaM特异性结合。研究表明,IQD 蛋白作为钙信号和生长素信号的中介来调控植物的形态发生[50],拟南芥和番茄中已证实了IQD家族基因的过表达导致茎叶等形态的改变[56,57]。因此,推测IQD9可能通过影响生长素信号和钙信号来影响油菜分枝角度。
本研究利用油菜基因组序列信息及亲本的重测序结果,从稳定表达的QTLcqBA. C4-2和cqBA.C4-3区段内,筛选出4 个油菜分枝角度的候选基因,为进一步理解分枝角度形成的分子机理提供了有价值的信息。
