81个矮牵牛自交系遗传多样性的分析
2022-03-10张西西张春和
张西西,李 芳,许 超,徐 菁,华 莹,张春和
(北京市园林绿化科学研究院/绿化植物育种北京市重点实验室,北京 100102)
矮牵牛[Petuniahybrida(J. D. Hooker) Vilmorin]别名碧冬茄、矮喇叭,番薯花,毽子花等,茄科碧冬茄属多年生草本植物,原产于南美洲阿根廷,现在世界各地广泛栽培。在温度适宜的情况下,矮牵牛一年四季均可开花,是节日花坛用花的常用材料[1-2]。对种质资源进行深入研究,是培育矮牵牛新品种的前提与基础。传统的植物形态标记等研究手段较少,此外,由于表型容易受到环境影响,性状表达容易不稳定[3]。目前在全世界范围内普遍采用分子标记技术研究植物遗传多样性和亲缘关系,相关的研究报道不少。迄今利用分子标记技术研究植物矮牵牛遗传多样性和亲缘关系的研究报道很多,但以矮牵牛为对象的相关研究报道还比较少。叶为[4]利用相关序列扩增多态性(sequence-related amplified polymorphism,SRAP)和简单重复序列间区标记(inter simple sequence repeat,ISSR)对18个矮牵牛自交系进行了亲缘关系的研究;吴志祥等[5]利用ISSR对142个矮牵牛品种进行遗传多样性分析;徐进等[6]利用SRAP对58个矮牵牛品种进行了遗传关系分析。利用扩增片段长度多态性标记(amplified fragment length polymorphism,AFLP)分析矮牵牛自交系的遗传关系尚未见报道。
AFLP分子标记技术是常用的一种分子标记方法,具有多态性强、稳定性高、重复性好等特点[7-8],是一种理想、有效的遗传标记方法,广泛用于植物亲缘关系等相关研究[9-12],同时,也在经济作物中被广泛应用[13-15]同时应用在蔬菜遗传关系的研究中[16-17]。在园林观赏植物研究中,AFLP广泛用于海棠等木本观赏植物[18-22]。由于AFLP技术相对SRAP等分子标记来讲,试验步骤较为复杂,近年来,采用SRAP等分子标记研究观赏花卉的遗传多态性较为常见[23-25],而用AFLP研究花卉的报道数量较少[26-28],利用AFLP研究矮牵牛自交系遗传多态性还未见报道。
该研究以北京市园林绿化科学研究院收集的81个矮牵牛自交系为材料,采用AFLP对其进行遗传多样性的分析,旨在明确自交系之间的亲缘关系,为新品种选育和品种保护提供理论依据和技术支撑。
1 材料和方法
1.1 试验材料
2013年在北京市园林科学研究院内进行81个矮牵牛自交系(表1)的种植。
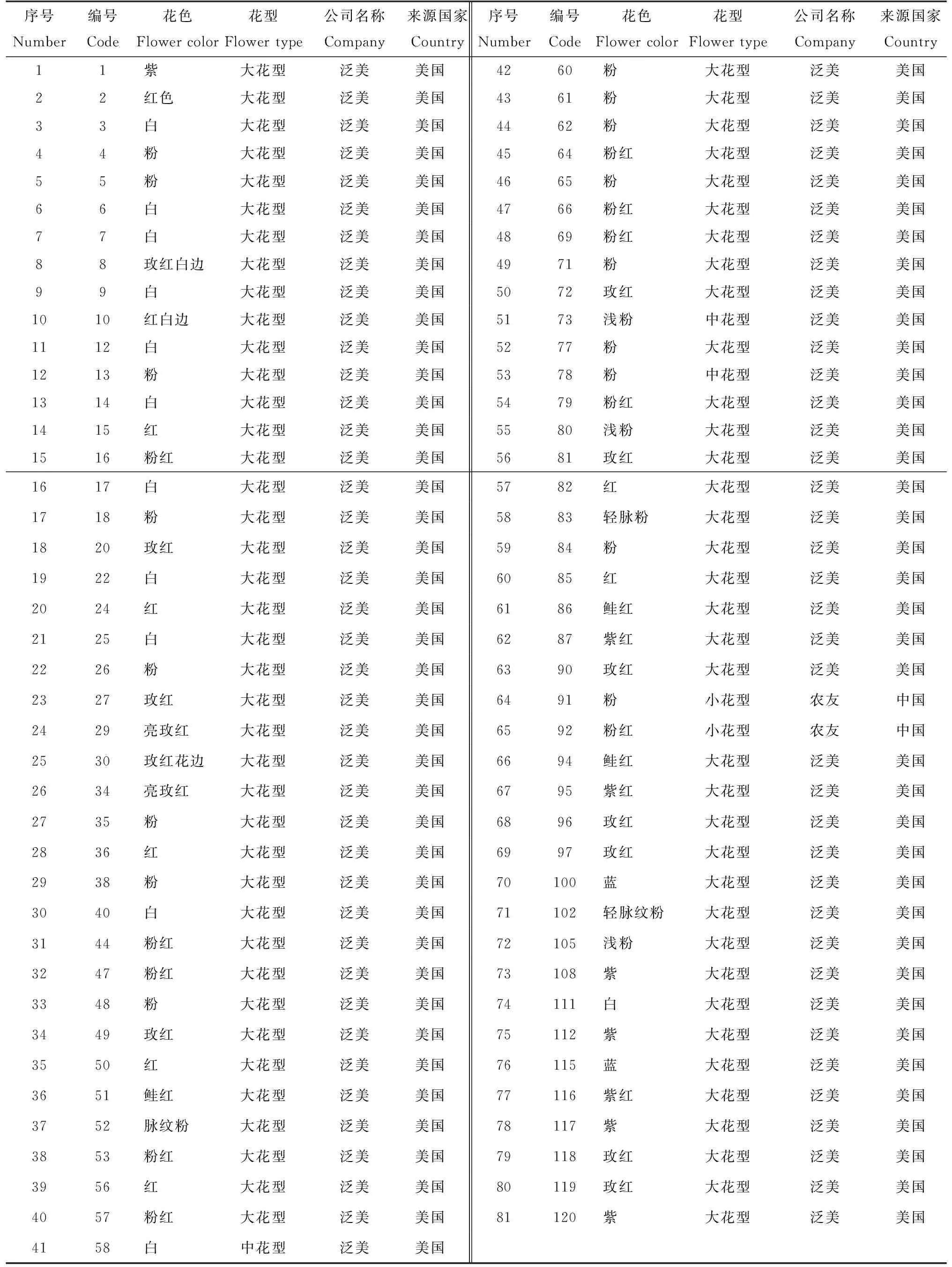
表1 81个矮牵牛自交系性状表Tab.1 Name and origin of Petunia hybrid germplasm used in the experiment
1.2 试验方法
AFLP试剂盒购自北京鼎国生物科技有限公司,试验方法参考VOS等[3]的方法。每个样品总DNA 200 ng,经限制内切酶(2 μL ECORI和2 μL MSel)消化后,在消化片段末段加入人工接头。
共使用8对选择性引物。其碱基组合为:H-AAG/M-CTC、H-ACA/M-CTC、H-AAG/M-CTT、H-ACT/M-CTT、H-AGC/M-CTC、H-AGC/M-CTT、H-ACC/M-CTC、H-AGG/M-CTT。
采用十六烷基三甲基溴化铵法提取矮牵牛幼叶基因组DNA,利用序列相关扩增多态性分子标记对基因组DNA进行多样性和相似性分析。在6%变性聚丙烯酰胺凝胶上分离扩增片段。凝胶用硝酸银染色,根据电泳结果得到“0”或“1”基质。多态位点数、多态位点百分比、有效等位基因数和遗传多样性指数由POPGENE版本32估算。利用NTSYS pc-V.2.1估计的遗传相似性对81个矮牵牛自交系按非加权组平均法(unweighted pair-group method with arithmetic means,UPGMA) 进行聚类分析,并通过Tree plot模块生成聚类图。
2 结果与分析
2.1 扩增结果
采用3个选择性碱基的引物组合,获得较好的扩增效果,电泳条带数目丰富,分带清晰(表2和图1)。8对引物组合扩增出216个遗传位点,81个矮牵牛自交系共获得24 351条扩增带,全部为多态性条带,多态性比率100%。81个矮牵牛自交系扩增带分布较均匀,扩增带最少的是258条,最多的是478条,平均每对引物扩增出3 044个多态性位点。
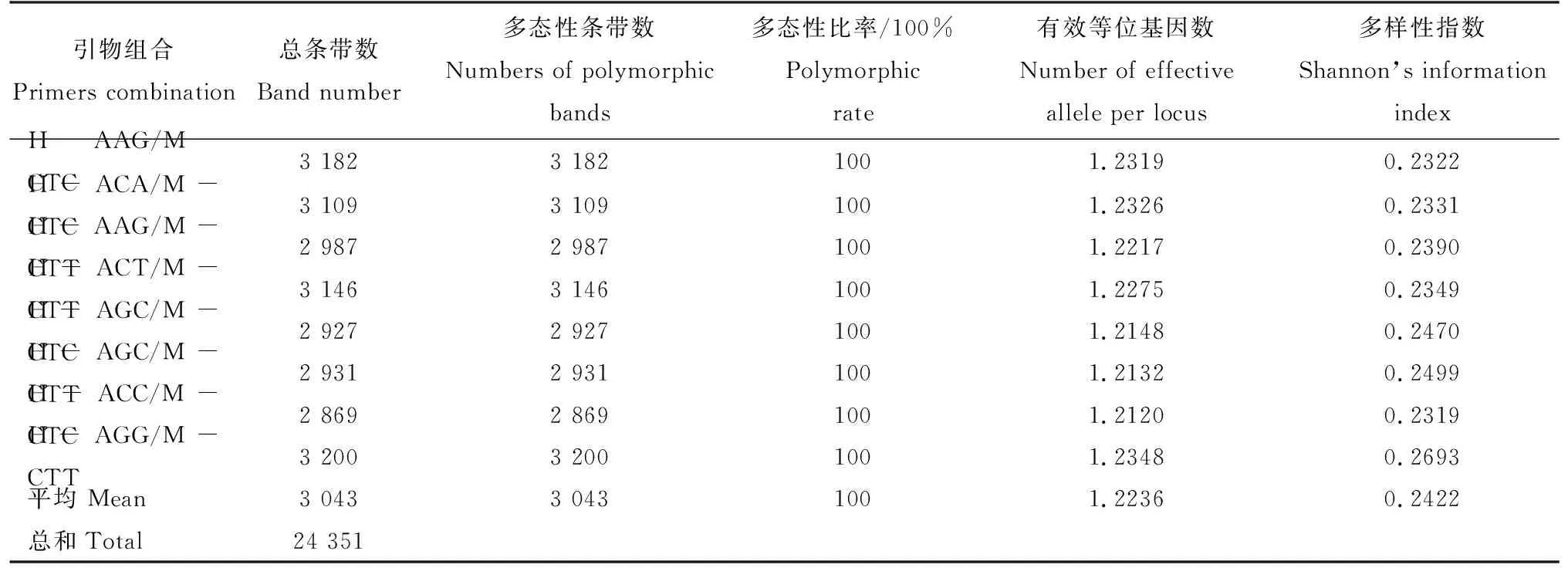
表2 8对引物扩增的多态性条数Tab.2 The number of polymorphic bands generated by 8 primer pairs
2.2 聚类分析
通过杰卡德相似系数,计算出遗传相似性系数在0.2443~0.6233之间,平均值0.4338,每个位点的有效等位基因数1.2236,平均多样性指数0.2422;遗传相似性系数最高的是4号和5号,达到0.6233,最低的是9号和92号,为0.2443。按非加权组平均法进行聚类,构建聚类图(图2)。在聚类图中,取遗传相似性系数水平0.36时,将聚类图中81个矮牵牛自交系分为3个类群。
2.3 利用AFLP对81个矮牵牛自交系的聚类结果
81个供试的矮牵牛材料中,全部为自交系。花色有红、粉、白、玫红、浅粉、蓝、紫红、紫等类型。相似系数0.36时,可将81个矮牵牛自交系分为3个类群:91和92号2个自交系,花色分别为粉和粉红的小花型自交系,单独聚为第Ⅰ类;58号、73号和78号,花色分别为白、浅粉和粉,为中花型自交系,聚为第II类。由于在传统的杂交育种中,一般会根据花径来进行自交系选育,长期以来,形成大花型自交系和小花型自交系。花径相近的自交系亲缘关系较近,这与叶为[4]的研究结果是一致。
其余76个大花型自交系聚为第Ⅲ类。由于这一类群的自交系数量比较大,根据色系的不同,将其分为8个小类群。100号花色为蓝,单独聚为A类群,115号为蓝,单独聚为B类群;22号花色为白,单独聚为C类群;60号、65号、66号、72号、80号、64号、77号、69号、81号、79号和71号共计11个自交系,均由北京市园林绿化科学研究院课题组从美国泛美种子公司的矮牵牛品种‘波浪’系列的玫红品种实生变异单株选育而来,为大花粉和浅粉系,聚为D类群;108和120号花色是紫,聚为E类群。102号轻脉纹粉和105号浅粉聚为F类群。14号、17号、35号、18号、90号、83号、84号、49号和53号共计9个自交系,分别为粉红、玫红、粉、轻脉纹粉等,聚为G类群;其余的归为H类群。
第Ⅲ类群中的H类型还可进一步细分为3个更小的类群。第①小类为编号1号、9号、3号和6号,花色为紫和白;第②小类编号为2号、85号、47号、4号、5号、12号、40号、30号、13号、20号、27号、15号、24号、36号、25号、38号、29号、34号、16号、48号、52号、50号、82号、56号、7号、111号、87号、116号、119号、96号、18号、97号、61号、62号、44号、57号、26号、95号、8号、10号、51号、86和94号,共计43个自交系;第③小类为编号112号和117号自交系,花冠紫色聚为一类。在第②小类中,主要集中白、浅粉、玫红、红、紫红、鲑红几个色系。这与徐进等[6]用SRAP进行矮牵牛F1代遗传关系的聚类结果基本一致,但略有不同。
3 讨 论
该研究通过筛选的8对AFLP引物组合对81个矮牵牛自交系进行多样性分析,共扩增出24 351条谱带,全部为多态性条带,多态性条带比率为100%。AFLP灵敏度高,能对亲缘关系较近的种质资源材料的亲缘关系进行遗传关系分析。AFLP聚类分析与形态学聚类总体结果一致。除11个变异株培育成的自交系外,81个矮牵牛自交系可分为3个大的类群。这一结果与花型为基础的聚类分析一致[4]。
81个矮牵牛自交系中,大花型的自交系占绝大多数。由于这个类群的自交系数量多,根据不同花色,将其分成蓝色、白色、粉色、紫色、浅粉色-轻脉纹以及粉红-玫红等不同亚类群。分子聚类结果为北京市园林绿化科学研究院的自育品种知识产权的保护提供重要依据。Ⅲ类群的D亚类群是北京市园林绿化科学研究院课题组根据自然突变单株选育的粉色系自交系,与其他自交系的遗传关系较远,可作为新品种选育的重点材料。
Ⅲ类群还有一部分自交系组成比较复杂,将其归为H亚类群,并进一步细分为3个更小的类群。第①小类为白色系+紫色系,由4个自交系组成;第②小类白色系+玫红等暖色系,由43个自交系组成;第③小类是2个紫色自交系。粉红色系和紫色、蓝色系自交系聚为不同的类群,同时白色系与冷暖色调的自交系分别聚在一起。在徐进等[6]的报道中,白色品种只和紫色等冷色调品种聚在一起。该试验结果是白色与粉色、红色系聚在一起,同时另一些白色自交系与紫色系聚在一起。而在传统的矮牵牛杂交试验中,粉色单株的分离后代会出现一些白色自交系,某些紫色单株在高温下会出现白色的花朵。传统杂交试验表明一些白色自交系与紫色自交系和粉色自交系的遗传关系都很近。该研究得出的分子聚类结果与杂交试验结果相吻合。分析原因可能是由于徐进等[6]试验材料采用的是F1代品种,白色品种可能分别由遗传背景较远的2个白色自交系杂交而成,与其他颜色的杂交一代遗传关系较远。而该研究所用材料全部来源于自交系,在自交过程中,白色自交系由其他颜色自交系分离而来,因而遗传关系较近。
在UPGMA聚类图上,81个矮牵牛自交系可根据花径和花色进行分类。这与吴志祥等[5]研究结果略有不同。分析可能是由于该研究采用的81个矮牵牛自交系最早由表型性状优良的单株自交多代而来,花型相同、颜色相近的自交系亲缘关系较近。而吴志祥等[5]采用的是来源于世界各地的杂交一代品种,由不同遗传背景的自交系杂交而来的可能性较大。由于遗传背景复杂,无法单一根据花型和花色进行完全聚类。对于矮牵牛育种来讲,世界各地的育种公司之间的遗传材料的交流比较频繁,同一系类不同颜色材料之间亲缘关系较远。同一品种的亲本可能来自遗传背景相差较大的自交系,这有利于矮牵牛杂交育种培育新品种。
根据UPGMA分子聚类图,D亚类群自交系全部由自然突变而来,与其他自交系遗传背景不同,在将来的育种工作中可作为重点育种材料。亲缘关系越远的自交系杂交,所得到的杂交一代的杂种优势有可能越强。在相应花径、花色范围内,尽量选择亲缘关系较远的自交系杂交,其后代的杂种优势出现的可能性越大。
