色氨酸卤化酶研究进展
2022-03-09雷鑫亭刘晓祺唐文竹李宪臻
雷鑫亭, 刘晓祺, 唐文竹, 李宪臻
(大连工业大学 生物工程学院,辽宁 大连 116034)
卤代化合物,是与制药、化学和农化工业相关的生物活性物质的重要中间体或成分[1],它存在于各种天然产品中,包括氯霉素、吡咯多霉素、植物生长调节剂噻诺林和抗真菌剂吡咯硝酸酯等[2]。在农业化学中,大约20%的小分子药物和30%的活性化合物是卤代物。其中色氨酸卤化物应用最为广泛,色氨酸又称β-吲哚基丙氨酸,其结构中的吲哚环是卤化物进行取代的位置[3]。常见的卤素有氟、氯、溴和碘,其中氯和溴的卤代产物是目前最常见也是研究最多的。近年来随着对卤化酶研究的逐渐深入,酶合成法以其优点正逐渐成为合成卤代物的主要方法[4]。尤其是色氨酸卤化酶对底物的高选择性使色氨酸可以在特定的位置进行卤代,得到人们想要的产物。另外,对色氨酸卤化酶结构的改变也可以改变其卤代的位置,因此色氨酸卤化酶逐渐得到关注。本文对色氨酸卤化酶的分类和来源、酶学性质、异源表达、结构与功能以及催化机理等方面的最新研究进展进行了综述。
1 色氨酸卤化酶的分类和来源
黄素依赖型卤化酶(Flavin-dependent halogenases, FDHs)是一类依赖黄素参与,催化天然产物卤化生成卤代化合物的酶[5-6]。其中研究最多的是色氨酸卤化酶(tryptophan halogenase),它是将卤素原子特异性地卤化在色氨酸中的吲哚环上,产生相对应的卤化产物的一种酶。根据卤素在吲哚环上取代位置的不同可以分为色氨酸2-卤化酶[7]、色氨酸5-卤化酶、色氨酸6-卤化酶和色氨酸7-卤化酶。自从1959年在海洋中发现第一株产生色氨酸卤化酶的Caldariomycesfumago(烟霉卡尔霉菌)[8-9]之后,目前已知能够获得色氨酸卤化酶的菌株有如下几种:Streptomycestoxytricini(毒三素链霉菌)[10]、Lechevalieriaaerocolonigenes(产气列契瓦尼尔氏菌)[11]、Streptomycesalbogriseolus(白浅灰链霉菌)[12]、Streptomycesalbus(白色链霉菌)[13]、Streptomycesviolaceusniger(紫黑链霉菌)[14]、Saccharomonosporasp.(糖单孢菌)[15]、Pseudomonasfluorescens(荧光假单胞菌)[16],其中以放线菌为主。
2 色氨酸卤化酶的酶学性质
作为最重要的色氨酸卤化酶之一,色氨酸7-卤化酶PrnA是Hammer等[16]在1997年从P.fluorescens中的硝吡咯菌素生物合成途径中发现的。PrnA此前已被证明可以催化色氨酸以外的许多吲哚衍生物的氯化反应。其分子量在55~60 kDa之间,最适作用温度为30 ℃。在这个生物合成途径中同时还发现了PrnC,也是色氨酸7-卤化酶[17]。RebH也是色氨酸7-卤化酶,它来自L.aerocolonigenes,同时存在的还有一种黄素还原酶RebF。RebH催化L-色氨酸氯化反应的Kcat值为1.4 min-1,Km值为2.0 μmol/L[18]。从Kutzneriasp. 744中扩增出KtzQ、KtzR和KtzS的基因,分别为色氨酸7-卤化酶、色氨酸6-卤化酶和黄素还原酶。其中KtzQ和KtzR进行串联作用,催化L-色氨酸氯化反应生成6,7-二氯-L-色氨酸。通过对KtzQ的动力学研究发现,以色氨酸和6-氯-色氨酸为底物的Km值均≤2 μmol/L,Kcat分别为0.19 min-1和0.07 min-1。KtzQ是一种区域特异性色氨酸7-卤化酶,在未经修饰的L-Trp的7位有卤化倾向,但也会催化6-氯底物的卤化[19]。
目前已经报道的色氨酸6-卤化酶有SttH、Thal、BorH、Th-Hal、Tar14、KtzR和SatH。从S.toxytriciniNRRL 15443中获得的SttH作用温度范围为20~55 ℃,作用pH范围为4.5~8.5,且在40 ℃和pH 6时活性最大。通过His6标记的SttH在大肠埃希菌中表达后, SDS-PAGE检测显示其大小为59 kDa[10]。来自S.albogriseolus的Thal也被称作ThdH,30 ℃,pH 7.2时活性最高。经过纯化后该酶分子量约为60 kDa。Thal的Kcat值为2.8 min-1,与色氨酸5-卤化酶PyrH(0.5 min-1)、色氨酸7-卤化酶PrnA(0.1 min-1)和RebH(1.4 min-1)的Kcat值相当[12,20]。BorH与BorF是由氯化双吲哚生物碱Borregomycin A的生物合成基因簇编码的两个蛋白质。BorH氯化色氨酸的最佳反应温度为45 ℃,Kcat为4.42 min-1,Km为9.78 μmol/L。该酶属于嗜热性的色氨酸卤化酶[21]。同样作为嗜热性的卤化酶Th-Hal则来自于S.violaceusniger。其最适反应温度也为45 ℃,Kcat和Km值分别为5.1 min-1和20.4 μmol/L[14]。海洋细菌Saccharomonosporasp.可以催化L-色氨酸转化为L-4-氯尿氨酸(L-4-Chlorokynurenine),这一过程需要四种酶进行催化,分别是Tar13、Tar14、Tar15和Tar16。其中Tar14为色氨酸卤化酶,其动力学参数Kcat和Km值分别为0.4 min-1和12 μmol/L[15]。前文提到的KtzR,除了可以将色氨酸卤化为6-氯-色氨酸,还可以将7-氯-色氨酸卤化为6, 7-二氯-色氨酸[18,22]。并且以7-氯-色氨酸为底物的Kcat (1.4 min-1)比以色氨酸为底物的Kcat(0.08 min-1)高约18倍,而Km值则仅为后者的1/7,因此KtzR对于7-氯-色氨酸的选择性高于未经修饰的色氨酸。SatH是由Lee等[13]通过将BorH的基因进行BLAST后在S.albus中发现的一种色氨酸6-卤化酶,与Th-Hal、 SttH和PyrH相比,在氯化反应中,SatH催化生成6-氯-色氨酸的产率仅低于Th-Hal,但在溴化反应中,SatH的产物得率最高。
相比于色氨酸6-卤化酶,色氨酸5-卤化酶的报道相对较少,只有SpmH、PyrH、MibH和XszenFHal。SpmH来源于Streptomycessp. SCSIO 03032,Liu等[23]通过基因缺失,体外表达的方式确定了其为色氨酸5-卤化酶。来自于StreptomycesrugosporusLL-42D005(卢氏链霉菌)中的PyrH,其最适反应温度为30 ℃,最适反应pH为7.2[24]。Foulston等[25]通过缺失分析确定了Microbisporacorallina(珊瑚小双孢菌)中几个mib基因的功能并说明了MibH是5位的色氨酸卤化酶。Jérémy等[26]克隆了Xenorhabdusszentirmaii(嗜线虫致病杆菌)中148个可能的卤化酶基因,表达后发现只有Fhal16对色氨酸具有卤化活性并命名为XszenFHal。其最适反应温度为25 ℃,Kcat值为4.44 min-1,Km值为58.22 μmol/L。
对于催化2号位取代的色氨酸卤化酶的研究较少。有报道的色氨酸2-卤化酶是CdmE,Rachid等[27]对ChondromycescrocatusCm c5(藏红花软骨霉状菌)的软骨酰胺生物合成基因簇进行了鉴定,证明CdmE是色氨酸2-卤化酶。
3 色氨酸卤化酶的异源表达
目前已知生产色氨酸卤化酶的菌大多是放线菌,因此对于该酶的研究多集中于基因克隆以及在E.coli(大肠埃希菌)中的异源表达[28-29]。从P.fluorescensBL915菌株中分离到一个包含四种酶的基因簇(prnABCD)[17],其中prnA基因产物催化L-色氨酸氯化生成7-氯-L-色氨酸。后来Burd等[30]将prnA基因连接到pUC18上,将重组质粒转入E.coliTG1中进行表达。同时又对在P.aureofaciensACN中发现的prnA基因进行研究,发现来自不同细菌的同种酶具有96%的同源性。从L.aerocolonige的基因组中扩增出色氨酸7-卤化酶RebH和RebF的基因[31],连接到pET28a上,转入E.coliBL21中进行表达。通过体外酶反应发现在没有RebF的情况下,RebH表现出低水平的氯化活性。通过改变两种酶组分的比例发现,RebF/RebH的比例为3∶1时活性最佳[18]。Zeng等[10]从S.toxytriciniNRRL 15443中克隆了色氨酸6卤化酶基因stth,连接至pET28a中并转入E.coliBL21 CodonPlus(DE3)-RIL中成功进行了表达。Lee等[13]从S.albusN-16041扩增出基因sath,克隆到pET28a中并转入E.coliBL21(DE3)中进行表达,得到色氨酸6-卤化酶SatH。
4 色氨酸卤化酶结构与功能
为了进行色氨酸卤化酶一级结构与功能的分析,选取了具有代表性的10个色氨酸卤化酶,它们均来自于细菌,分别是催化2号位卤化的CmdE,催化5号位卤化的PyrH和MibH,催化6号位卤化的Thal、KtzR和SttH,催化7号位卤化的RebH、KtzQ、PrnA和PrnC。通过在NCBI数据库中搜索获得它们的氨基酸序列后,使用CLC Sequence Viewer软件对这10组氨基酸序列进行比对后发现(图1),色氨酸卤化酶中有3个高度保守的关键基序,分别是GxGxxG、WxWxIP和FxxPxxSxG。其中N-末端附近的GxGxxG基序可能是与黄素进行结合的位点,而序列中间的WxWxIP推测是为了防止酶催化单加氧反应而存在的,最后的FxxPxxSxG基序目前并没有明确的认识。从图1中可以看出KtzR并没有GxGxxG基序,分析可能是因为它是用来催化底物7-氯-色氨酸卤化的,很可能还存在其他的酶来帮助其完成反应。目前虽然确定了KtzR具有卤化酶特征,但哌嗪部分的氯化还没有被详细阐明,因此需要对Kutzneriasp.进行进一步的基因组分析。
从最早得到PrnA的晶体之后,已经有1个色氨酸5-卤化酶(PyrH),2个色氨酸7-卤化酶(PrnA和RebH)和4个色氨酸6-卤化酶(SttH、Th-Hal、Thal和Tar14)的晶体结构被解析出来。以PrnA为例(图2A),其结构为1.95 Å,由2~518个残基组成,是一个二聚体。每个单体都是一个单一的结构域,形状像一个盒子,一面连着一个三角形的金字塔。这个盒子推测是FAD的结合模块,被两个大的β折叠所占据。异咯嗪环的C4-N5边缘位于反向平行的β折叠的一个面上方。序列比对表明,在黄素依赖型卤化酶中,只有黄素结合模块是保守的[32]。这些晶体结构的阐明极大地促进了对反应机理和区域选择性的理解[33-34]。
色氨酸卤化酶的产物立体选择性非常高, 即每种酶通常只能有一个取代卤化位置,得到的产物结构十分确定[31]。对于天然底物L-Trp,依赖于黄素的色氨酸卤化酶的卤化位点不是由底物的电子因素决定,而是由底物在活性位点的取向决定的。在色氨酸7-卤化酶PrnA与底物色氨酸复合物的三维结构中(图2B),赖氨酸(K79)和谷氨酸残基(E346)位于底物附近,两者都是酶催化所必需的氨基酸。Flecks等[35]认为次氯酸与赖氨酸和谷氨酸残基的协同作用能增加卤素原子的亲电性,并且还能确保卤素离子的正确定位。吲哚环中的其他活性位置被大的芳香族氨基酸色氨酸(W455)和苯丙氨酸(F103)所遮蔽,底物被夹在中间。色氨酸的羧酸和氨基与两个酪氨酸残基和一个谷氨酸残基相互作用,吲哚的NH基团与肽键氧原子形成氢键,导致吲哚环的7号位指向隧道(酶中的一条通道),进而使得色氨酸吲哚环的7号位置成为卤素原子唯一可以进行结合的位置。因此PrnA的结构决定了其只能催化色氨酸在7号位进行卤化。该理论同时被色氨酸5-卤化酶PyrH的结构所证实,在色氨酸5-卤化酶PyrH的三维结构中,催化所涉及的氨基酸残基与异丙嗪环的位置与它们在色氨酸7-卤化酶PrnA中的位置相同。同样在色氨酸5-卤化酶PyrH与底物色氨酸复合物的三维结构中,色氨酸吲哚环的5号位置是指向隧道的,因此进行的是5位上的卤化[8]。

图1 色氨酸卤化酶氨基酸序列比对Fig.1 Amino acid sequence alignment of tryptophan halogenasesRebH, GenBank:AAN01206.1;Thal, GenBank:ABK79936.1;KtzQ, GenBank:ABV56597.1;PrnA, GenBank:AAB97504.1;KtzR,GenBank:ABV56598.1;SttH, GenBank:ADW94630.1;PyrH,GenBank:AAU95674.1;MibH,GenBank: ADK32653.1; PrnC, GenBank:AAY92872.1; CmdE, GenBank:CAJ46693.1
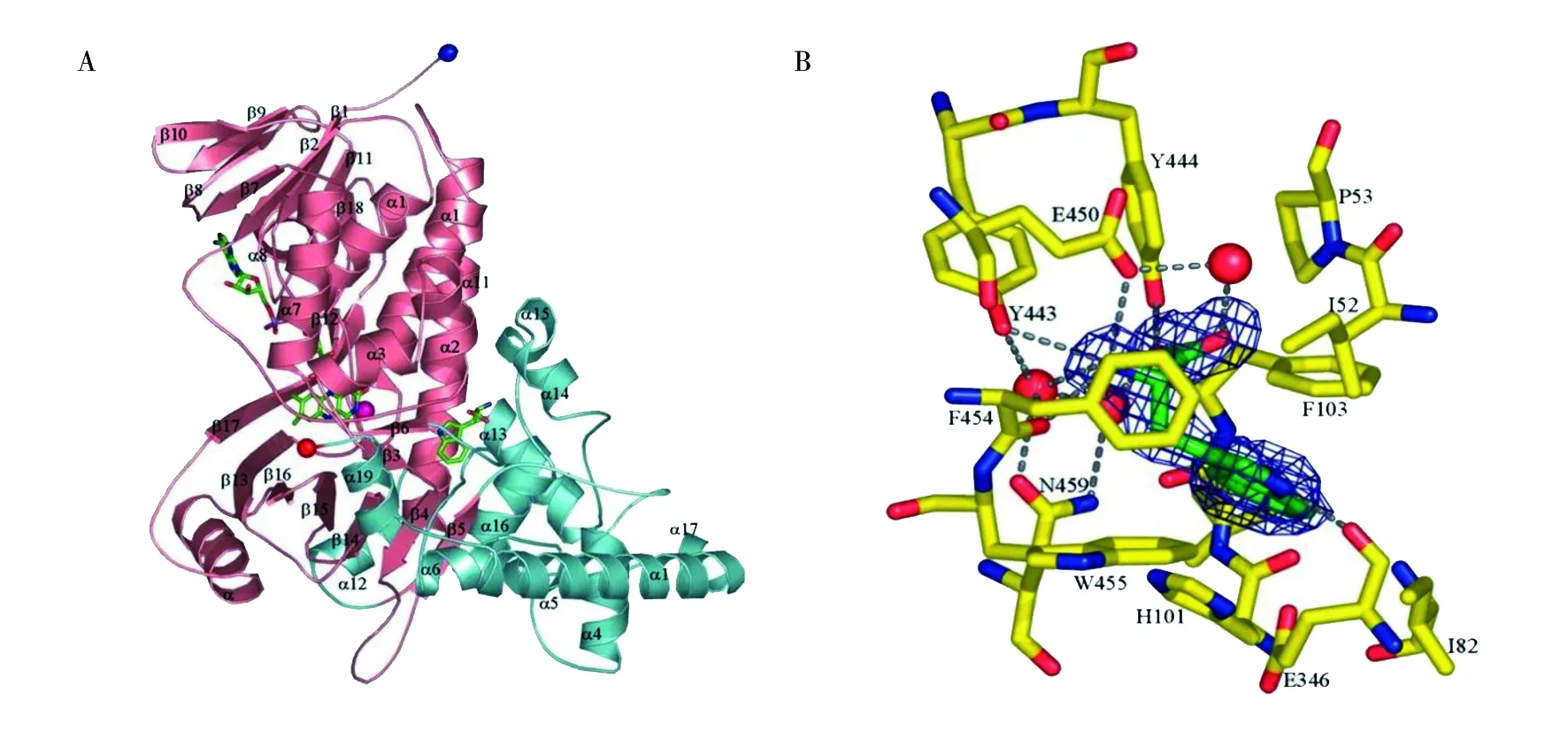
图2 色氨酸卤化酶PrnA结构(A)以及与底物的结合位点(B)[32]Fig.2 Structure of tryptophan halogenase PrnaA(A) and bingding site with substrate(B)[32]
2010年,Lang等[8]对PrnA的氨基酸残基苯丙氨酸(F103A)进行突变,使其从7位卤化变为5位卤化为主,首次证明了色氨酸卤化酶的区域选择性可以通过定点突变来实现。对色氨酸6-卤化酶Thal进行研究发现,尽管Thal在序列和结构上更接近色氨酸7-卤化酶RebH而不是色氨酸5-卤化酶PyrH,但Thal活性位点上的吲哚结合形式更接近PyrH。Moritzer等[36]将Thal活性位点上的5个氨基酸与RebH中的相应氨基酸进行了交换,产生了5个突变体Thal-RebH5。Thal-RebH5突变体对色氨酸的总体转化能力与野生型Thal相似,但其氯化和溴化的区域选择性几乎完全从6位转变为7位,虽然仍有少量6位卤代产物存在。因此,以少量残基为靶点的基于结构的蛋白质工程是改造色氨酸卤化酶的有效途径之一[37]。
5 色氨酸卤化酶的催化机理
色氨酸卤化酶属于黄素依赖型卤化酶,目前关于黄素依赖型卤化酶的催化机理已经基本明确。要使底物卤化,依赖于黄素的卤化酶需要还原的黄素腺嘌呤二核苷酸(FADH2)、分子氧和卤化物盐(通常是Cl-或Br-)共同参与[38]。通常黄素还原酶与黄素依赖型卤化酶一起存在于生物合成的基因簇中,形成一个双组分系统,例如RebH和RebF。因此其作用机理可以分为两步:还原反应和卤化反应。还原反应是黄素还原酶将FAD还原为FADH2。之后进入卤化反应,先前生成的FADH2与黄素依赖型卤化酶中的FAD结合模块结合,FADH2就会与分子氧反应生成黄素过氧化氢(FAD(C4a)-OOH)。然后,FAD(C4a)-OOH与卤化物阴离子反应,卤素变成次卤酸(HOX),黄素过氧化氢变成FAD(C4a)-OH。次卤酸向酶活性部位扩散,主要是向催化必需的赖氨酸残基的ε-氨基移动,在那里底物发生卤化反应。动力学分析证实,底物卤化发生在黄素氧化还原反应完成之后,底物不参与从FADH2到FAD(C4a)含氧物再到FAD的反应[36]。图3为催化机理示意图。
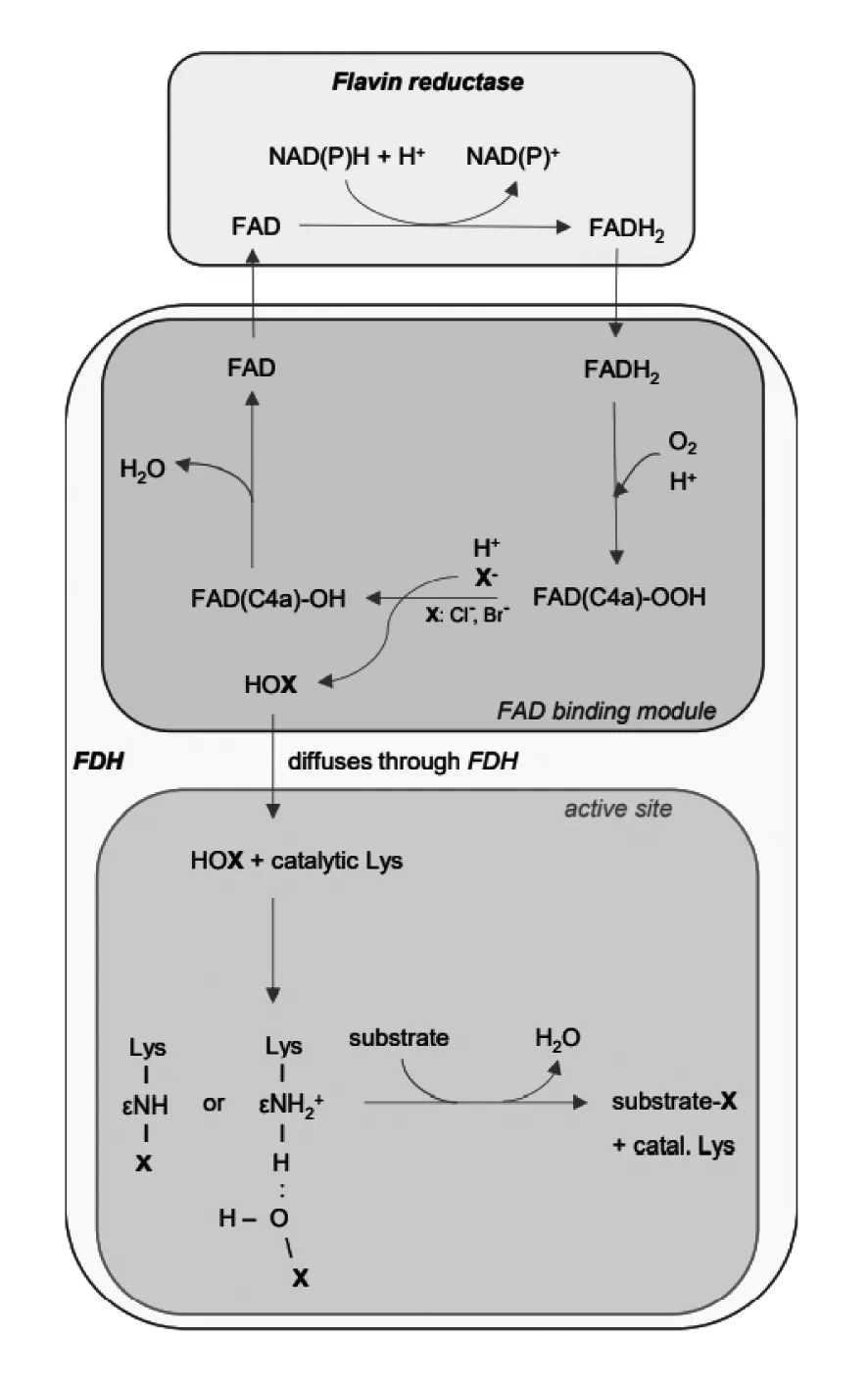
图3 黄素依赖型卤化酶(FDH)催化机理图[36]Fig.3 Catalytic mechanism of flavin dependent halogenase (FDH)[36]
色氨酸卤化酶与黄素依赖型卤化酶的催化机理基本相同,只是反应底物为色氨酸。而色氨酸被卤化的位置是通过底物相对于催化必须的赖氨酸的ε-氨基的定位来决定的。底物色氨酸的5、6、7号位与赖氨酸的接近程度,可以决定色氨酸在哪个位置发生卤化。
此外人们对FAD还原机制的细节知之甚少。例如,FAD是在与黄素依赖型卤化酶结合的情况下被还原,还是必须从卤化酶上解离后才被还原,目前还不完全清楚[36]。
6 色氨酸卤化酶的应用
卤代芳香化合物有广泛的应用,是医药、农用化学品和其他贵重材料的重要合成成分[39]。传统的卤代芳香化合物的生产需要苛刻的反应条件,往往需要有害的试剂、催化剂和溶剂。另外反应选择性较差,会产生多种副产物。不仅副产物分离困难,并且由于其在环境中的毒性或持久性而难以处理。因此,利用自然界的卤化酶实现更清洁和更有区域选择性的卤化反应一直是研究的重点[9]。
色氨酸卤化酶具有在特定位置卤化的特点,可以用来生产相应的色氨酸卤化衍生物。软骨酰胺(Chondramides)是含有2氯色氨酸的化合物,色氨酸残基可经氯化修饰成软骨酰胺B和软骨酰胺D的中间产物。研究表明,软骨酰胺是由CmdE(色氨酸2-卤化酶)在色氨酸吡咯环的2号位氯化产生的[27]。吡咯烷霉素B(Pyrroindomycin B)是一种对革兰阳性菌包括耐甲氧西林金黄色葡萄球菌(MRSA)有效的抗生素,是从S.rugoporus中分离出来的。其结构包括一个聚酮大环和三糖连接到从氯化色氨酸衍生的吡咯吲哚部分。氯化反应发生在吲哚环的5位,催化反应的酶是MibH(色氨酸5卤化酶)[25]。噻诺林(Thienodolin)是一种从S.toxytricini中分离出来的吲哚硫酚生物碱,其中关键的卤化反应是由两个色氨酸6卤化酶Thal和SttH共同催化完成的[10]。Kutznerides是从Kutzneriasp.中分离得到的具有抗真菌和抗细菌活性的环状六肽,在6位和7位都有氯原子。与噻诺林类似,Kutznerides也是由两个酶共同作用形成的,分别为KtzQ(色氨酸6-卤化酶)和 KtzR(色氨酸7-卤化酶)[19]。瑞贝卡霉素(Rebeccamycin)是从Saccharothrixaerocolonigenes(气生菌落诺卡氏菌)中分离出来的一种抗肿瘤化合物,其关键性中间产物是在RebF的存在下,RebH催化L-Trp区域特异性氯化生成的7-氯色氨酸[18]。
目前已知的色氨酸卤化酶对氯原子有明显的偏好性,对于溴原子活性较低,其他卤素碘和氟则基本没有活性[40]。氯化色氨酸和溴化色氨酸在自然界中通常不以游离形式存在,而是作为复杂结构中的生物合成前体存在。目前有文献报道可以在低氯、高溴浓度的培养基中培养能表达RebH和RebF的谷氨酸单胞菌,从而生产7-Br-Trp[41]。另外,虽然色氨酸卤化酶PrnA不能碘化其天然底物色氨酸,但可以催化一些非天然底物的碘化,尽管其转化率仅在1%左右[42]。
7 展 望
目前国内外对于卤化酶的研究还不够深入,特别是一些卤代天然产物的生物合成途径尚未完全明确。其中有机溴化合物在海洋中普遍存在,含氯化合物则主要来自于陆地,而天然有机碘化合物相对较少,含氟化合物更是少之又少。因此继续开发新的卤化产物,并找到催化卤化的关键酶就显得尤为重要。
在众多卤化酶中,色氨酸卤化酶是其中被研究最广泛的一类。虽然已经获得了多个色氨酸卤化酶晶体,但是仍不能完全揭示其结构和催化机理的相互关系,特别是卤化位点的选择性机理还不十分明确。本课题组(微生物与生物催化课题组)最近从1株海洋放线菌中克隆到一个新的色氨酸卤化酶基因,与已知的色氨酸卤化酶的同源性较低。将其在E.coli(大肠埃希菌) BL21(DE3)中进行异源表达后发现,该酶可以催化色氨酸卤化产生两种产物;通过液质联用以及核磁共振氢谱测定,确定这两种卤化产物分别为7-氯色氨酸和6, 7-二氯色氨酸。因此这是一种全新的双功能色氨酸卤化酶,目前正在进行这一新酶的结晶相关研究,同时也在通过理性和非理性设计改造该色氨酸卤化酶,希望能增强其卤化活性,同时获得更多新的色氨酸卤化产物。
另外,对于色氨酸卤化酶的研究主要集中于5位、6位和7位,而对于能卤化色氨酸吡咯环上其他位点的卤化酶的研究则较少,需要不断寻找和发现新的色氨酸卤化酶。宏基因组数据分析是寻找新的卤化酶的有力工具,其巨大优势在于其不仅提供了可培养微生物的基因组数据,而且还提供了非可培养微生物的基因组数据。但是,从没有明显特征的有机体中确定酶的天然底物仍然是一个不小的挑战。不断发展的现代化技术将为色氨酸卤化酶基因的发现、功能的确定以及结构的改造提供极大的便利,酶法卤化取代化学方法将是一个必然趋势。随着研究的深入,关于色氨酸卤化酶结构和功能的研究必将会有新的突破,卤化产物的人工合成也将进入新的发展阶段。
