不同生境下川麦冬根围土壤丛枝菌根真菌多样性
2022-03-09杨蕊毓邓錡璋田丽平田嘉旺邱成书刘红玲
杨蕊毓,邓錡璋,2,田丽平,田嘉旺,邱成书,刘红玲*
(1 成都师范学院 化学与生命科学学院,成都 611130; 2 石河子大学 生命科学学院,新疆石河子 832003)
麦冬(Ophiopogonjaponicus)为百合科沿阶草属多年生常绿草本植物,医学上普遍认为其在消炎、抗癌等方面有效[1],主要生产于四川和浙江,分别称为川麦冬和杭麦冬。麦冬喜温暖湿润,在降雨充沛的气候条件5~30 ℃下才能正常生长[2],而四川大部分属于亚热带湿润和半湿润地区,年均降雨量充沛,因此为麦冬提供了良好的生长条件,这也是四川野生型麦冬资源丰富的原因之一。随着新兴技术的发展,野生麦冬被过度采挖,其生长环境的破坏也逐渐加剧,野生资源日益枯竭,而栽培麦冬由于土地、技术等资源的限制,产量及品质仍存在大量问题。
丛枝菌根(AM)真菌是普遍存在于根围的一种有益真菌,它能够与陆地生态系统90%以上的植物形成共生关系,以促进植物吸收养分且能影响植物对自身次生代谢物的积累[3]。AM真菌在提高药用植物的药用成分方面效果显著,接种AM真菌能促进黄花蒿地上部分挥发油收油[4],提高半夏块茎鸟苷、生物碱等药用成分的含量[5]。AM真菌在与植物形成菌根后,其根外菌丝不断扩展到土壤之中,穿过土壤颗粒的间隙,其分泌的有机物质等会直接影响到土壤的化学性质[6]。菌根共生双方的多样性对维持生态系统可持续性具有不可代替的作用[7]。
目前,已有大量关于药用植物AM真菌资源的研究,但对药用植物川麦冬根围土壤 AM 真菌资源的研究未见报道。本研究利用高通量测序技术,通过分析2种不同生境川麦冬根围土壤的 AM 真菌多样性,旨在揭示2种生境川麦冬根围土壤 AM 真菌多样性和优势群落分布特点,探讨AM真菌群落分布差异的驱动因子,为AM 真菌应用于麦冬生产提供理论依据和技术支持。
1 材料和方法
1.1 研究区概况
采样地分别为四川省成都市温江区(N 30°41′48.36″, E 103°49′57.78″)和四川省攀枝花市仁和区(N26°8′44.26″, E101°48′30.39″),其中成都温江地处成都平原,四周有群山环抱,平原内四季分明,日照少、气候温和,降雨充沛,属亚热带季风气候区,海拔600 m左右,土壤类型为水稻土。而攀枝花市仁和区属亚热带温带多种气候类型,该区夏季长,四季不分明,而具有旱、雨季分明,昼夜温差大,气候干燥,小气候复杂多样等特点,海拔约1 500~2 000 m,土壤类型为红壤。
1.2 样品采集
本试验以川麦冬为研究对象,选取2种生境,栽培环境(C)取自成都温江麦冬栽培基地,野生环境(N)为攀枝花仁和区平地镇金沙江干热河谷地区。于2020年9月采集2种生境川麦冬根围土壤。每个生境随机选取麦冬20株,每株按东南西北四个方位取样,除去地表表层杂物后,采集10~20 cm左右的土层土样及根系装入塑料袋中,放置于冰盒带回实验室。土壤样品一部分自然风干用于土壤理化性质测定,一部分存放于-80 ℃冰箱用于分子鉴定。
1.3 土壤理化性质的测定
土壤理化性质的测定参考鲍士旦《土壤农化分析》[8],土壤有机质含量用重铬酸钾容量法-外加热法、土壤全氮含量用半微量凯氏法、土壤全磷用HClO4-H2SO4法、土壤全钾含量用火焰分光光度法、土壤有效磷含量用0.5 mol·L-1NaHCO3法、土壤速效钾含量用NH4OAc浸提-火焰光度法。
1.4 AM真菌的分子鉴定
1.4.1 川麦冬根围土壤总DNA的提取使用土壤DNA提取试剂盒(Ezup Column Soil DNA Purification Kit)提取不同样地的川麦冬根围土壤总DNA,完成基因组DNA抽提后,利用1%琼脂糖凝胶电泳检测抽提的基因组DNA。
1.4.2 巢式PCR扩增利用AM真菌特异引物(表1)对所提DNA进行巢式-PCR扩增。每个样本设置3个重复,将同一样本的PCR产物混合,2%琼脂糖凝胶电泳检测,AxyPrepDNA凝胶回收试剂盒(AXYGEN公司)回收PCR产物,使用Tris-HCl洗脱,2%琼脂糖电泳检测。参照电泳初步的定量结果,将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统(Promega公司)进行检测定量,之后按照每个样本的测序量要求,进行相应比例的混合。
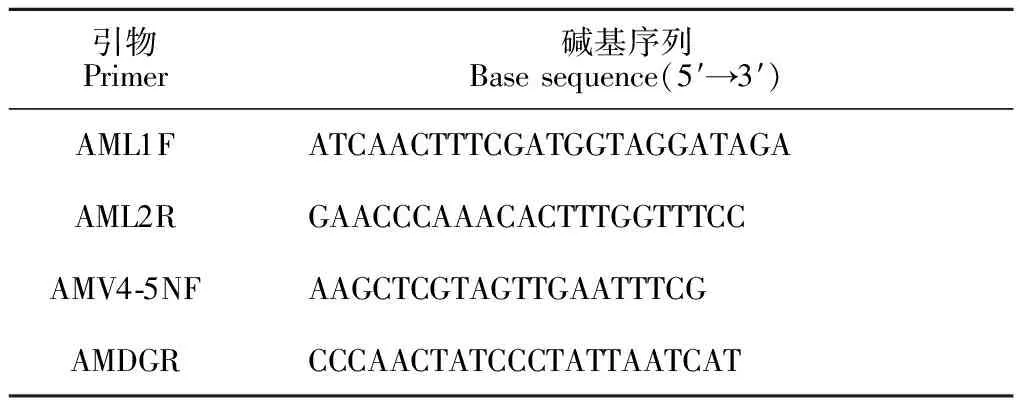
表1 巢式-PCR扩增引物
1.4.3 Miseq文库的构建与测序通过PCR将Illumina官方接头序列添加至目标区域外端;使用凝胶回收试剂盒切胶回收PCR产物;Tris-HCl缓冲液洗脱,2%琼脂糖电泳检测;氢氧化钠变性,产生单链DNA片段。文库构建试剂使用TruSeqTM DNA Sample Prep Kit。利用Illumina公司的Miseq PE300平台进行测序(委托于上海美吉生物医药科技有限公司)。
1.5 生物学信息分析
平台测序得到的PE reads首先根据overlap关系进行拼接,同时对所得序列的质量进行质控及过滤,区分不同样本后进行OTU聚类分析及物种分类学分析。根据不同的相似度水平,对序列进行OTU水平划分,通常以97%相似水平的OTU进行生物信息学统计分析。
为了得到每个OTU所对应的物种分类信息,采用RDP classifier贝叶斯算法对97%相似水平的OTU代表序列进行分类学统计分析,并分别在各个分类学水平:域(Domain)、界(kingdom)、门(Phylum)、纲(Class)、目(Order)、科(Family)、属(Genus)和种(Species)统计各样本的群落物种组成。
1.6 统计学数据分析
土壤理化性质、AM真菌多样性指数采用SPSS 19.0进行单因素方差分析,提取97%相似度OTU,利用mothur计算Alpha多样性指数,计算ACE、Shannon及Simpson等多样性指数,用Origin 2019分析绘制不同科、属水平下丰度图,R语言circlize包分析绘制不同样本与种水平的关系图,Pearson分析法及R语言cor函数分析绘制土壤理化因子与AM真菌多样性指数、属水平丰度之间的相关性。
2 结果与分析
2.1 不同生境川麦冬根围土壤理化性质
2个生境川麦冬根围的土壤理化性质见表2。表明野生环境(N)下川麦冬根围土壤全钾、全磷、有效磷含量均显著低于人工栽培环境(C)(P<0.05)。而野生环境下川麦冬根围土壤全氮、有机质含量均显著高于人工栽培环境(P<0.05)。此外,2个生境的速效钾含量则表现出无显著差异。
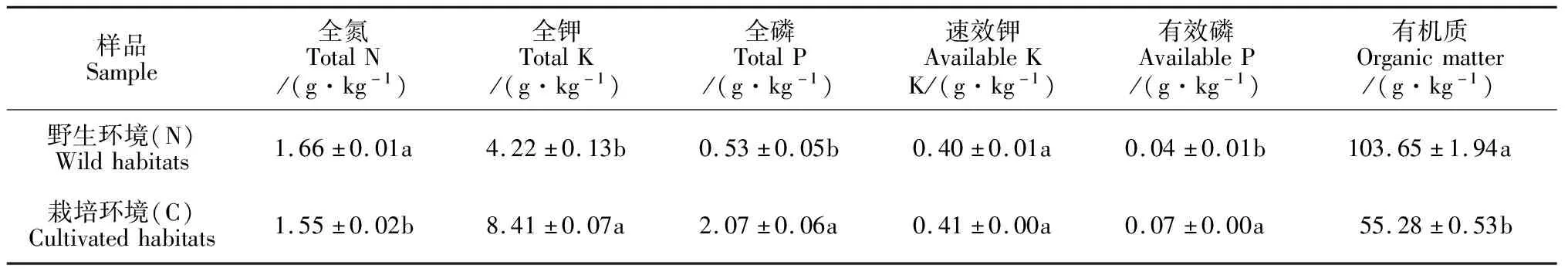
表2 土壤理化性质
2.2 不同生境川麦冬根围AM真菌多样性
使用生物学Alpha多样性指数ACE、Shannon、Simpson表征川麦冬根围土壤中AM真菌的多样性(图1)。结果显示,野生环境(N)川麦冬根围土壤AM真菌多样性指数ACE、Shannon均显著大于栽培环境(C)麦冬根围土壤AM真菌,而Simpson指数则相反。
2.3 AM真菌分子多样性鉴定分析
基于AM真菌18S rRNA基因的高通量测序,分析了不同生境川麦冬根围土壤中AM真菌种属多样性。以97%为阈值对测序序列划分操作分类单元(OTU),共获得31个OTU,31个OTU分属于3个科3个属10个种。分析发现,野生型川麦冬根围AM真菌群落组成结构与人工栽培型差异显著(图2-4)。人工栽培型川麦冬根围鉴定出AM真菌1个科1个属6个种,1个科为球囊霉科(Glomeraceae),1个属为球囊霉属(Glomus),6个种分别为Glomus-MO-G18-VTX00064、Glomus-MO-G22-VTX00125、Glomus-MO-G23-VTX00222、Glomus-Wirsel-OTU16-VTX00156、Glomus-acnaGlo2-VTX00155和Glomus-group-B-Glomus-acnaGlo7-VTX00057。而野生环境中川麦冬根围鉴定出AM真菌3个科3个属7个种,3个科分别为无梗囊霉科(Acaulosporaceae)、多孢囊霉科(Diversisporaceae)和球囊霉科,3个属分别为无梗囊霉属(Acaulospora)、多孢囊霉属(Diversispora)、球囊霉属,7个种分别为Acaulospora-Acaulospora1-3-VTX00201、Diversispora-sp.-VTX00058、Glomus-GlAd3.3-VTX00289、Glomus-MO-G22-VTX00125、Glomus-MO-G23-VTX00222、Glomus-group-B-Glomus-acnaGlo7-VTX00057和Glomus-group-B-Glomus-lamellosu-VTX00193。两个生境共有1个科为球囊霉科,共有1个属为球囊霉属,共有3个种为Glomus-MO-G22-VTX00125、Glomus-MO-G23-VTX00222和Glomus-group-B-Glomus-acnaGlo7-VTX00057。两个生境的优势科均为球囊霉科,优势属均为球囊霉属。
2.4 土壤理化性质与AM真菌多样性的相关性
对不同生境川麦冬根围土壤理化因子与AM真菌多样性指数进行相关性分析(图5),川麦冬根围土壤AM真菌多样性指数与土壤理化因子存在相关性。AM真菌ACE指数和Shannon指数与土壤理化因子存在相同的趋势,均与土壤全钾(TK)、全磷(TP)呈显著负相关,与土壤全氮(TN)呈显著正相关,与土壤速效钾(AK)、有效磷(AP)呈负相关,与土壤有机质(OM)呈正相关。而AM真菌Simpson指数与土壤全钾(TK)、全磷(TP)呈显著正相关,与土壤全氮(TN)呈显著负相关,与土壤速效钾(AK)呈正相关、与土壤有效磷(AP)呈显著正相关,与土壤有机质(OM)呈显著负相关。其中,TK、TP、TN为不同生境川麦冬根围土壤AM真菌多样性的主要驱动因子。
2.5 土壤理化性质与AM真菌群落结构的相关性
对不同生境川麦冬根围土壤理化因子与AM真菌属水平相对丰度进行相关性分析发现(图6),无梗囊霉属和多孢囊霉属均与土壤TP、AP、TK显著负相关,与土壤OM、TN显著正相关,球囊霉属则表现出与土壤TP、AP、TK显著正相关,而与土壤有机质、碱解氮显著负相关。进一步分析发现,无梗囊霉属和多孢囊霉属与土壤理化因子的相关性变化趋势与球囊霉属与土壤理化因子的相关性变化趋势相反,而球囊霉属与无梗囊霉属和多孢囊霉属均呈显著负相关。其中,TK、TP、TN、AP、OM为不同生境川麦冬根围AM真菌群落结构的主要驱动因子。
3 讨 论
3.1 不同生境下川麦冬根围AM多样性
微生物的多样性指数是用来描述某一微生物类型的个体数和均匀度的重要指标,可以直接地反映某一微生物群落的种类丰富度[9]。本研究中,野生环境多样性指数ACE和Shannon指数均显著大于人工栽培川麦冬,而Simpson指数则相反。进一步说明野生环境下川麦冬根围AM真菌丰富度显著大于人工栽培环境。大量研究已经证实,AM真菌产生的根围菌丝能帮助植物获得更为丰富的营养[10],这一结果说明在野生环境中麦冬需要更丰富的AM真菌帮助其生长发育,而人工栽培下,由于施肥等人为因素的影响,麦冬根围AM真菌丰度有所减少。杨高文等[11]研究表明,AM真菌的多样性较高时有利于植物群落的稳定性,并促进植物生产力,这可能是野生麦冬成药品质高于栽培的原因之一。
3.2 不同生境下麦冬根围AM真菌群落结构
AM 真菌的分类鉴定是分析 AM 真菌多样性的关键。近年来,通过分析 18S rDNA基因序列鉴定 AM 真菌种类,因其与传统的形态学鉴定方法相比更具科学性和准确性,成为 AM 真菌分类研究的重要手段。潘佩蕾等[12]利用分子鉴定在杭麦冬根际土壤分离鉴定了10种菌根孢子,其中球囊霉属和巨孢囊霉属为主要优势类群。同样的本研究通过Illumina平台测序对野生和栽培两种不同生境下的麦冬根围AM真菌进行分析,两个生境共鉴定出3个科3个属,且不同生境麦冬根围土壤AM真菌优势属均为球囊霉属,可作为麦冬植株筛选优势菌种的依据。该结果与大量研究结果类似,球囊霉属在中国广泛分布,适应性强,属于广谱性共生系统真菌[13-14]。可能是由于球囊霉属能利用自身菌丝及共生菌根的根段进行繁殖[15-16],相比于其他种属其具有更强的产孢能力和侵染宿主根系。有研究表明不同生境下植物根围AM真菌多样性存在显著差异,程俐陶等[17]对贵州地区栽培及野生半夏进行传统形态学鉴定,发现野生半夏根围土壤中AM真菌种类、数量及活力均高于栽培半夏。周丽思等[18]采用巢式-PCR 和 DNA 克隆测序技术分析了西双版纳地区野生与栽培绞股蓝根内 AM真菌的 18S rDNA 序列,结果显示,相比于栽培绞股蓝,野生绞股蓝根内的AM真菌种类更加丰富。本研究中,在野生环境下麦冬根围共鉴定出AM真菌3个属7个种,而人工栽培麦冬根围鉴定出AM真菌1个属6个种。野生环境下麦冬根围AM真菌显然更为丰富,与前人研究结果一致。
3.3 不同生境麦冬根围AM真菌群落多样性与土壤理化因子的相关性
AM真菌广泛存在于土壤环境中,植物从AM真菌定殖中受益,在很大程度上取决于土壤环境条件[19]。曹敏等[20]对重庆地区茅苍术根际丛枝菌根真菌多样性和土壤因子进行冗余分析表明,根际土壤因子对AM真菌的群落组成和丰度存在显著影响,是导致AM真菌群落结构地理分布格局差异的重要原因之一。本研究中,栽培麦冬和野生麦冬根围土壤理化因子全钾、全磷、全氮、速效钾、有效磷及有机质含量均表现出显著差异,且野生环境多样性指数Ace和Shannon指数均显著大于人工栽培麦冬,Simpson指数则相反,说明土壤理化因子对AM真菌分布存在影响。进一步对土壤理化因子和AM真菌多样性指数ACE、Shannon及Simpson进行相关性分析发现,土壤理化因子TK、AK与AM真菌多样性指数ACE和Shannon均呈显著负相关,与Simpson指数呈显著正相关。该结果与张含眉等[21]研究结果相似,但与张之为等[22]所得出土壤K含量减少,促使AM真菌类群呈现上升趋势的研究结果相反,推测一定范围内丰富钾能促进菌根真菌的形成,而过高的钾反而抑制AM真菌菌丝的发育。同时,本研究中土壤理化因子TP、AP与AM真菌多样性指数ACE和Shannon均呈显著负相关,而Simpson指数则相反。与王化秋等[23]根系速效磷与AM真菌丰富度指数均显著负相关的研究结果类似,而与杨春雪等[24]得出土壤磷与AM真菌物种丰富呈显著正相关的结果相反。同样地,推测土壤内一定浓度的磷能促进菌根共生体的发育,而过高的磷含量则抑制菌丝的形成。李楠海等[25]提出在80 mg/kg磷水平,接种丛枝菌根真菌对桔梗的促生效果最佳。AM真菌与麦冬的接种效应相关研究表明[26],土壤中K含量直接影响菌根的形成,在浓度约为0.18 g/kg时,促进麦冬菌根的形成,K浓度大于0.24 g/kg时侵染率显著降低,说明过高的K不利于菌根的发育。另外,本研究土壤理化因子TN与AM真菌多样性指数ACE和Shannon均呈显著正相关,而Simpson指数则相反。该结果与Jansa等[27]研究结果相反。推测土壤N含量对AM真菌菌丝的形成以及分布的影响存在一定的阈值。土壤有机质与AM真菌多样性指数ACE和Shannon均呈正相关,而Simpson指数则相反。与刘敏等[28-29]研究结果相似。相关研究说明土壤有机质含量过高,肥力过剩,抑制菌根的功能发挥和AM真菌的生长,导致AM真菌丰度下降[30]。同样有研究发现,AM真菌对其所处环境的养分状况较为敏感,丰度过高的养分供应将对AM真菌孢子的萌发和菌丝的生长产生影响,间接地减少AM真菌对宿主植株的侵染与共生[31]。
栽培麦冬根围土壤全磷、全钾、速效钾、有效磷含量均显著大于野生麦冬,这与栽培麦冬人工施磷肥、钾肥有关。而栽培麦冬根围AM真菌多样性指数小,可能是因为栽培条件下,麦冬根围营养充足,不需要与AM真菌侵染形成菌根结构来获得其余营养。有研究表明,在减量施肥状态下,植物根系AM真菌的菌丝侵染率高于常规施肥[32]。孔垂豹等[33]探究不同施肥水平下AM真菌对甘蔗生长的影响表明,在减量施肥水平下,AM真菌能更好地发挥对植物的促生效益。
3.4 麦冬根围AM真菌群落结构与土壤理化因子的相关性
土壤理化因子作用于宿主植物从而间接地影响了AM真菌的生长繁殖与分布[34]。高甜甜等[35]对不同品种甘草根际土壤AM真菌群落多样性进行分析发现,不同属的AM真菌与宿主植物根系土壤理化性质之间所存在的相关性各不同。本研究对麦冬根围土壤理化因子与AM真菌属水平群落结构进行相关分析发现,土壤磷、钾、氮含量和有机质丰度均对AM真菌属水平群落结构存在显著影响。其中AM真菌无梗囊霉属、多孢囊霉属相对丰度与土壤磷含量显著负相关,而AM真菌球囊霉属与其则呈显著负相关的趋势。以往有研究表明,球囊霉属能在磷含量较高的土壤环境中繁殖,并能提高宿主对磷的吸收[36]。进一步分析发现,球囊霉属与无梗囊霉属和多孢囊霉属均呈显著负相关。有研究表明,处于互利共生关系的微生物呈正相关,而处于竞争关系的微生物则呈负相关[37],故推测球囊霉属与其他两个属存在竞争关系。
4 结 论
本实验利用高通量测序技术对不同生境下麦冬根围AM真菌多样性进行分析,共鉴定出AM真菌3个科3个属10个种。在不同生境下球囊霉属均为麦冬根围优势属,可作为麦冬植株菌种筛选的实验依据。不同生境下麦冬根围AM真菌多样性差异显著,野生麦冬根围AM真菌较为丰富。进一步对土壤驱动因子和AM真菌多样性指数及属相对丰度进行Pearson分析发现,AM真菌群落分布及结构组成与土壤理化因子显著相关,但不同的因子相关性表现存在差异,而同一土壤驱动因子对AM真菌发育的驱动存在一定的阈值,过高的土壤肥力不利于菌丝的发育。土壤N、P、K和有机质含量是影响麦冬根围AM真菌群落结构的主要因素,且不同种属的AM真菌对其所处生境的需求不同。本研究为进一步优化麦冬的栽培措施和合理施肥提供了理论参考。
