耳蜗中tip-link 张力与静纤毛运动动力学研究*
2022-03-04徐旭马文凯姚文娟2
徐旭 马文凯 姚文娟2)†
1) (上海大学力学与工程科学学院,上海 200444)
2) (上海市应用数学和力学研究所,上海 200444)
诠释耳蜗的主动感音放大机制一直是未解的医学难题.这种机制与耳蜗中外毛细胞顶端的静纤毛运动密切相关,静纤毛运动又受到tip-link 张力与淋巴液流体力的调节.因此,研究静纤毛运动过程中tip-link 张力是诠释耳蜗的主动感音放大机制的重要环节.本文把静纤毛视为变形体,基于泊肃叶流动理论并结合分布参数模型,推导了静纤毛运动的解析解.研究了盖膜剪切荷载作用下静纤毛和淋巴液相互作用的动力响应以及tip-link 张力的变化规律.研究发现:当静纤毛的杨氏模量减小时,在小于峰值频率的区域,tip-link 张力显著增大,f2 的峰值频率减小.以往的研究将静纤毛作为刚体,势必导致低频声音信号作用减弱.当系数c=0 (无黏性阻力)时,f2 频率选择特性存在;当μ=0(无压力)时,f2 频率选择特性消失,因此淋巴液可能是通过在静纤毛间产生压强的方式来调节毛束的频率特性的.另外,盖膜剪切荷载频率越高,静纤毛轴弯曲越明显,发束内外域的压强差也越大.
1 引言
迄今为止,揭示耳蜗的主动感音放大机理仍然是医学难题[1].耳蜗具有围绕蜗轴盘旋2.5—2.75周的蜗管,它能够对宽频域的声音进行敏锐识别.这种功能来自于沿蜗管纵向排列的外毛细胞以及顶端分布的静纤毛和内淋巴液之间的相互作用及能量转换.较短静纤毛的顶端生长出tip-link 并向斜上方延伸至相邻较高静纤毛的侧壁[2,3].通常情况下,静纤毛会因受到外部作用而产生振动,这个振动过程受到发束与淋巴液耦合作用的调控,并在tip-link 中产生张力.耳蜗主动放大机制受到tip-link 张力的调节[4,5].
耳蜗感音放大机制是一种非线性的主动过程,这种非线性是由发束的运动提供的,发束运动和电致运动共同作用产生了耳蜗放大的特性[6].早在1992 年,Fettiplace 等[7]就发现单个毛细胞上的离子通道总数约为55 个,处于静纤毛数量(50—100)的最低水平,这说明单个毛细胞中离子通道数量可能少于静纤毛的数量.Beurg 等[8]和Furness 等[9]先后发现三排静纤毛中的最高的那排静纤毛表面完全没有离子通道蛋白,在中间和最低静纤毛的顶端与tip-link 下端相连的位置存在离子通道蛋白,这与Fettiplace 等[7]测量的通道与静纤毛在数量上具有一致性.离子通道在静纤毛上位置说明了tip-link 与离子通道蛋白存在着某种机械相互作用,这具有里程碑的意义.2013 年,Park 和Wei[10]利用泊松-能-普朗克方程进行三维数值模拟证实了tip-link 连接着发束的离子通道.
Jaramillo 和Hudspeth[11]早期的研究发现施加在发束上的力可以通过门控弹簧传递到离子通道.tip-link 门控与发束运动相结合,可以减小发束的机械刚度,从而有助于微小刺激的非线性放大.Lim 和Park[12]开发出静纤毛的宏观模型,仿生这种非线性放大,发现微小刺激的非线性放大是通过离子通道门控的刚度软化而机械产生的.Kachar等[13]的研究认为构成tip-link 的螺旋结构蛋白丝可能与门控弹簧串联,并且具有一定的硬度.进一步的研究发现tip-link 是通过钙黏蛋白23 (cadherin 23,CDH23)和原钙黏蛋白15 (protocadherin 15,PCDH15)的相互作用形成的[14-16].PCDH15的顺式二聚体结构对于感知机械刺激至关重要[17].Bartsch 等[18]发现单个原PCDH15 分子具有弹性,推测tip-link 可能是离子通道的门控弹簧,门控弹簧的力学性能可能受到tip-link 张力的影响.
以往的研究大多将静纤毛视为刚体,在研究发束机械特性时忽略了轴弯曲的影响.弹性静纤毛的研究大多运用有限元仿真模拟,或者通过忽略与淋巴液的流固耦合作用来做简化处理,或者只考虑静力加载的情况.Duncan 和Grant[19]建立了有限元模型来研究静纤毛的刚度和挠度.Cotton 和Grant[20,21]基于弹性理论建立了静力作用下的分布参数模型来研究静纤毛的挠度、刚度与几何结构、材料性能之间的关系.Nam 等[22]建立了考虑淋巴液黏滞阻力的三维有限元模型,研究了流体作用力和点荷载作用下tip-link 张力在发束中的分布情况,但该模型的流体作用力是作为外荷载施加在发束上的,并没有考虑发束与淋巴液的流固耦合作用.目前对发束特性的理论研究还不完善.鉴于此,本文基于分布参数模型以及泊肃叶流原理,推导了流固耦合作用下可变形静纤毛的无限自由度动力控制方程,分析研究了流固耦合中tip-link 张力与静纤毛运动规律.
2 理论模型
2.1 系统力假设
静纤毛在根部逐渐变细,形成一个倒置的锥体,锥体底面固定在毛细胞上,因此整体上呈现为上部为柱体结构和根部为锥体结构.单根静纤毛的简化模型是流体环境中的弹性梁结构.相比于上部柱体结构,由于根部锥体结构的长度较小,因此本文不对锥体结构进行细致研究,而是把其简化为扭转弹簧将柱体结构连接到毛细胞上.tip-link 在结构上为纤维高分子编织对或螺旋聚合物,相对僵硬[13].因此本文将tip-link 简化成两端铰接在相邻静纤毛上的弹性杆,其响应服从弹性理论.
相邻静纤毛的相对运动会对其间的淋巴液受到挤压,同时在侧壁表面产生压力.淋巴液压力与tip-link 张力均有由静纤毛的运动而产生,并反过来调节静纤毛运动.这两种力以内力的形式存在于发束系统中,但却以外力的形式作用在单个静纤毛上.为了便于表述,本文将tip-link 张力和淋巴液压力称为系统力.
本模型中lt1,lt2分别为tip-link 在高、中静纤毛侧壁上的连接点到根部的距离(如图1 所示).假设盖膜在高静纤毛顶端产生的水平剪力为Fp,tiplink 张力为FNj,淋巴液压力为FLi.对整个系统而言,Fp是已知的主动力,FNj和FLi是待求的被动力.它们的表达式分别假设为

图1 发束理论模型示意图Fig.1.Schematic diagram of theoretical model of hair bundle.
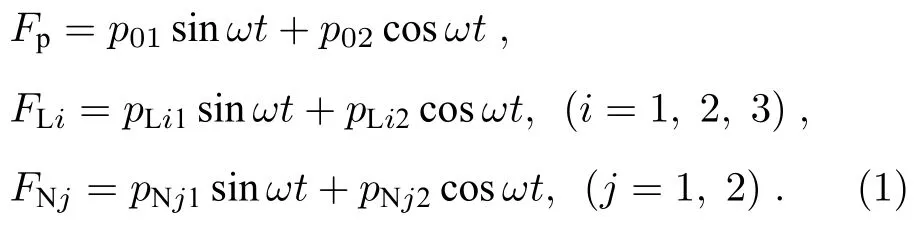
正、余弦函数相加的形式使简谐荷载表达更具有一般性,根据三角函数变换法则可知,正、余弦项系数的相对关系可以反映荷载的相位大小.其中下标i1,2,3 分别表示高、中、短静纤毛,下标j1,2分别表示连接高、中静纤毛2 和中、短静纤毛的tip-link.pNj1,pNj2为未知常数,pLi1,pLi2为x的未知函数.
2.2 弹性位移
研究了在盖膜和网状板运动产生的剪切作用下静纤毛的动力响应.根据叠加原理,位移动力响应可分解为两个部分,分别由静纤毛自身轴弯曲变形和根部偏转产生.前者可以简化为弹性悬臂梁模型,后者则为固定端为扭转弹簧铰接的刚性悬臂梁模型,如图1 所示.由于前者是弹性体,后者是刚体,因此,将由静纤毛自身轴弯曲变形产生的位移称为弹性位移,根部偏转产生的位移为刚性位移.
由达朗贝尔原理及无限自由度体系振型函数的正交性推导出的单根静纤毛的运动方程为
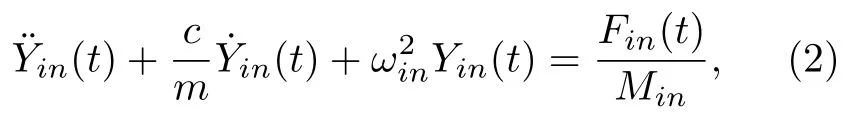
其中下标n为振型序号;c是淋巴液粘滞系数;m是静纤毛分布质量,即线密度;ωin是静纤毛i第n阶自振角速度;等号右边为广义力,Fin(t)为静纤毛i第n阶振型力,Fi(x,t)是静纤毛i受到的外力,φin(x)是静纤毛i的第n阶振型函数;为静纤毛i第n阶振型质量.
根据前文对荷载的假设可得,任一广义力均可表示为

Ai1n,Ai2n分别为广义力正、余弦分项系数,与荷载正、余弦分项系数相关.由振型叠加法可得静纤毛i的弹性位移yi1为

其中

与荷载频率及结构自身属性相关;ω是荷载角速度;阻尼比ςinc/(2mωin).由于位移响应的瞬态解随着时间的推移会趋近于0,因此这里的弹性位移将其忽略,只考虑了位移响应的稳态解.
2.3 刚性位移
实际情况下,静纤毛的柱体与锥体结构均会产生弯曲变形,并有相应的横向位移.相比于柱体,锥体的高度较小,由变形产生的横向位移也很小,对上部柱体结构的位移响应影响不大,但是锥体上平面产生的转动会带动柱体旋转,并随高度增加不断放大上部柱体的横向位移.本文假设锥体横截面半径呈线性变化(如图2 所示),并以锥体上平面转角作为静纤毛柱体轴的转角,记为θ.当静纤毛顶端有剪力FQ作用时,由卡氏第二定理得
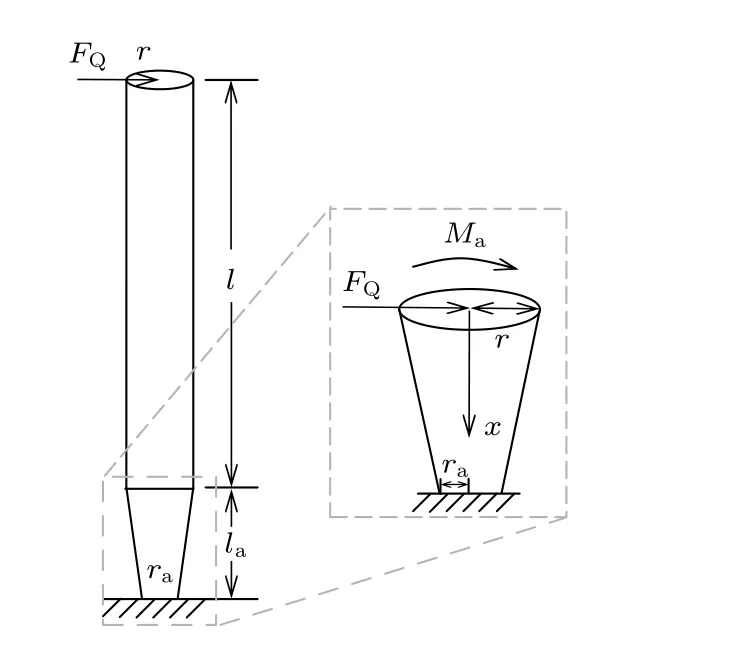
图2 单根静纤毛根部转角计算模型Fig.2.Calculation model of root rotation angle of a single stereocilia.

其中E是静纤毛的杨氏模量;r是静纤毛体半径;ra是锥体下平面半径;la为锥体的高;δ1可视为弯曲柔度;δ2为剪切柔度;MaFQl为锥体上平面受到的弯矩.
根据柔度法建立刚性轴静纤毛的运动方程为

其中和分别为tip-link 张力和淋巴液压力在锥体上平面产生的弯矩和剪力,对于高静纤毛还包括盖膜作用产生的弯矩和剪力.对于小变形的静纤毛,其刚性位移为

根据叠加原理,静纤毛的位移动力响应是弹性位移与刚性位移之和,即

2.4 tip-link 张力
tip-link 张力响应服从胡克定律,张力-位移关系如下:

其中ktipj和βj(j1,2)分别为对应tip-link 的弹性常数和与水平方向间的夹角.
2.5 相邻静纤毛间淋巴液流体压力
静纤毛运动过程中,淋巴液在行和列两个承载面(如图3(a)虚线椭圆形区域)之间流动.图3(a)中蓝色区域为发束的内域空间,淋巴液流体压强为以发束外域空间流体压强为零点时的相对压强,用pre 表示.当静纤毛在兴奋/抑制平面内振动时,各静纤毛间淋巴液压强在x2方向上相等,在x1方向淋巴液压强有所差异,因此压强梯度只存在于x1方向,这就使得淋巴液只通过行承载面在发束不同内空间域或内外空间域之间流动.在四根静纤毛围成的内域空间中,静纤毛运动时淋巴液的流向如图3(b)所示.

图3 发束横断面图 (a)压力分布的位置;(b)相邻立体纤毛之间的淋巴流线Fig.3.Top view of hair bundle:(a) The position of the pressure distribution;(b) lymphatic streamlines between adjacent stereocilia.
在行承载面上,淋巴液的流动假设为泊肃叶流动,速度方程为

根据方程(10)对速度在通道跨度上积分可得到通过不同行承载面的流量为

Δprei(i1,2,3)是静纤毛i左右两侧的压差.由于静纤毛行与列交点处的控制体积,满足全局连续性,因此不同行承载面上的流量满足如下关系:
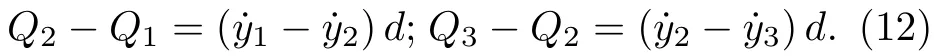
这里,d是静纤毛直径.联立方程(11)和方程(12)得相邻静纤毛间淋巴液压强的方程为

以上是发束从根部至高度为l3位置范围内的压强,通过 同样的方法可得出距离根部l3到l2位置范围内的压强为

对任意两点间的压强梯度进行积分,能够得到这两点之间的压强差.在此基础上,在得到发束内域空间的压强后,理论上本文可以计算出静纤毛表面的淋巴液压强分布.压强产生一个垂直于静纤毛表面的法向力,表面合力即为淋巴液压力FL.

以上是发束底部到高度l3的淋巴液压力计算公式.同理,可以得到距离根部l3到l2位置范围的压力计算公式为

基于上述过程,推导出系统力FNj和FLi的一组高阶方程组,求解该方程组,则得到系统力的解.本文用MATLAB 数值计算对上述方程进行求解.
3 结果与讨论
3.1 模型验证
基于已报道的豚鼠耳蜗测量数据建立了理论模型[13,23-32].由于微管在决定细胞形状方面起着主导作用,所以这里将静纤毛的杨氏模量大小取为微管的杨氏模量的实验值(1.2 GPa).各参数值如表1 所列.

表1 模型参数Table 1.Model parameters.
由于现有研究中关于静纤毛动态测量的数据缺乏,因此本文使用已发表的最高静纤毛的静刚度实验值来验证本文模型的正确性.由于本文的计算模型是动力响应,因此用激励频率趋于0 时,动荷载趋于静荷载,力幅值与位移幅值之比可视为静刚度的计算值,其频率响应曲线见图4,本文计算值与实验值的对比见表2.从图4 可以看到,激励频率趋近于0 时,最高静纤毛刚度K=6.25×10—4N/m (力幅值为10 pN,位移幅值为15.99 nm),这与Flock 和Strelioff[33]以及Szymko 等[34]实验数据相吻合,从而验证了模型的正确性.

表2 静刚度的实验值与解析值Table 2.Experimental value and analytical value of static stiffness.

图4 单根静纤毛刚度Fig.4.Stiffness of single stereocilium.
3.2 位移比
Tilney 和Tilney[23]拍摄的电子显微镜照片发现静纤毛根部蛋白丝(filaments)的弯曲会通过特殊的方式传递到上部所有蛋白丝中,揭示了静纤毛存在轴弯曲的生物学机理.Fridberger 等[35]通过快速共焦成像方法(a method for rapid confocal imaging)发现毛细胞静纤毛存在微小的挠曲.本文能够借助理论模型剖析这种挠曲产生的内在机理.为了考察轴弯曲对响应的贡献,引入位移比的概念.位移比是静纤毛上某点弹性位移与刚性位移理论最大值的比值,它能反映弹性位移相对比重.
杨氏模量为1.2 GPa 时,静纤毛顶端位移比的频域变化曲线如图5 所示.从整体上看,高静纤毛的位移比最大,中静纤毛次之,短静纤毛最小,这可能是由于长度不同造成的,因为高静纤毛顶端能产生更大的弹性位移积累.Cotton 和Grant[20]从静力学模型中发现不考虑轴弯曲的限制性假设会严重影响静纤毛的变形轮廓,且静纤毛越高,影响越大.本文的动力学模型同样能够得出与之类似的结果.另外,在频率小于1000 Hz 的频域内,三根静纤毛的位移比变化不明显,且数值较小;当频率大于1000 Hz 时,高静纤毛的位移比显著增大,进一步揭示了激励频率对轴弯曲贡献的影响.
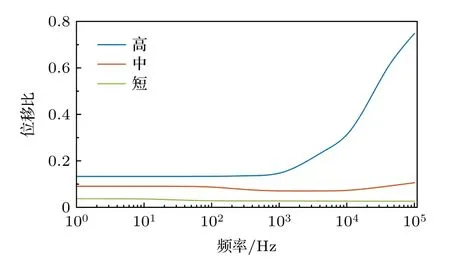
图5 静纤毛顶端位移比随频率的变化曲线Fig.5.Variation curve of displacement ratio of the top of stereocilia with frequency.
3.3 杨氏模量的影响
首先对tip-link 张力做了归一化处理,定义归一化张力fj(j1,2)是tip-link 张力幅值与盖膜剪切荷载幅值之比.作为测量tip-link 张力相对大小的无量纲指标,归一化张力可由下式求得

Vlajkovic 等[36]将成年大鼠在不同声压水平(sound pressure level,SPL)下暴露72 h 后发现,90 dB SPL 导致大鼠听觉阈值(临时阈值移位)发生微小而短暂的变化,而110 dB SPL 则导致听觉阈值(永久阈值移位)发生剧烈和永久性变化.Avinash 等[37]使用3D 显微镜技术检查豚鼠耳蜗的第二圈和第三圈时发现,在噪音暴露的耳蜗中,Factin 的解聚和聚合都可能在听觉过度刺激后发生在毛细胞的静纤毛中,而F-actin 含量与杨氏模量密切相关[38].也就是说,噪声在改变静纤毛中Factin 含量的同时会改变静纤毛杨氏模量.本文计算了不同杨氏模量(E=0.8,1.0,1.2,1.4,1.6 GPa)下1—105Hz 频域内的tip-link 归一化张力,并将结果绘制在图6 中.

图6 不同杨氏模量下tip-link 归一化张力随频率的变化曲线 (a) f1 ;(b) f2Fig.6.Variation curves of tip-link normalized tension with frequency under different Young's modulus:(a) f1;(b) f2.
发束独特的结构使其在足够宽的频域内具有一定的动力特性.当频率增加到到一定值时,归一化张力f1迅速减小,表现出类低通滤波特性,归一化张力f2仅在某一频率域内反应强烈,在该频域外迅速减小,表现出类共振特征.在低频域,f1和f2与杨氏模量均呈明显的负相关,即在杨氏模量降低后对低频的声音响应更强.可以看出,当杨氏模量足够小时,f2表现出类低通滤波和类共振特性叠加的现象,这可能会削弱发束的频率选择性.在高频时,杨氏模量对f1和f2影响不大.这一现象可能是由于淋巴液的黏性在高频区域对静纤毛的运动有更强的阻碍作用,而这一作用在低频区域不明显.此外,本研究还发现,杨氏模量降低会减小f2的共振频率,同时增加共振的峰值.
Duncan 和Grant[19]对静纤毛的有限元分析表明,发束刚度随静纤毛杨氏模量的增加而增加,而刚度增加的速度逐渐减小.本文的结果显示,当静纤毛杨氏模量定量增加时,曲线之间的距离逐渐减小,这与发束刚度增长速度的减小直接相关.产生这种结果的力学机制如下所述.
相邻静纤毛间的最大位移差和高静纤毛顶端最大位移分别表示为 Δy和 Δytip.由于,因此 Δytip∝Δy.将发束刚度表示为k.根据k和fj的定义可得,k和fj的乘积是一个常数,所以fj与刚度k一样是发束的固有属性.Duncan 和Grant[19]先前的研究结果用数学关系表达为:∂k/∂E >0和∂2k/∂E2<0.综上所述可以得出∂fj/∂E <0和∂2fj/∂E2>0,即fj随着杨氏模量的增加而减小,变化率逐渐减小.
3.4 黏滞阻力和流体压力的影响
淋巴液对静纤毛的影响表现为黏滞阻力和流体压力.黏滞阻力是静纤毛表面受到的摩擦力,与静纤毛轴的横向速度有关.关于黏滞阻力本文做了忽略淋巴液微观运动的隐含假设.流体压力是流体受压后对静纤毛的反作用力,与静纤毛表面垂直.为了方便考察,本文将控制黏滞阻力和流体压力的的系数使用不同符号来表示.本文分别计算了不考虑黏滞阻力(c=0)和不考虑流体压力(μ=0)的tip-link 归一化张力值,并与同时考虑黏滞阻力和流体压力的tip-link 归一化张力值进行比较,来研究两种力的存在对tip-link 张力变化的影响,结果如图7 所示.
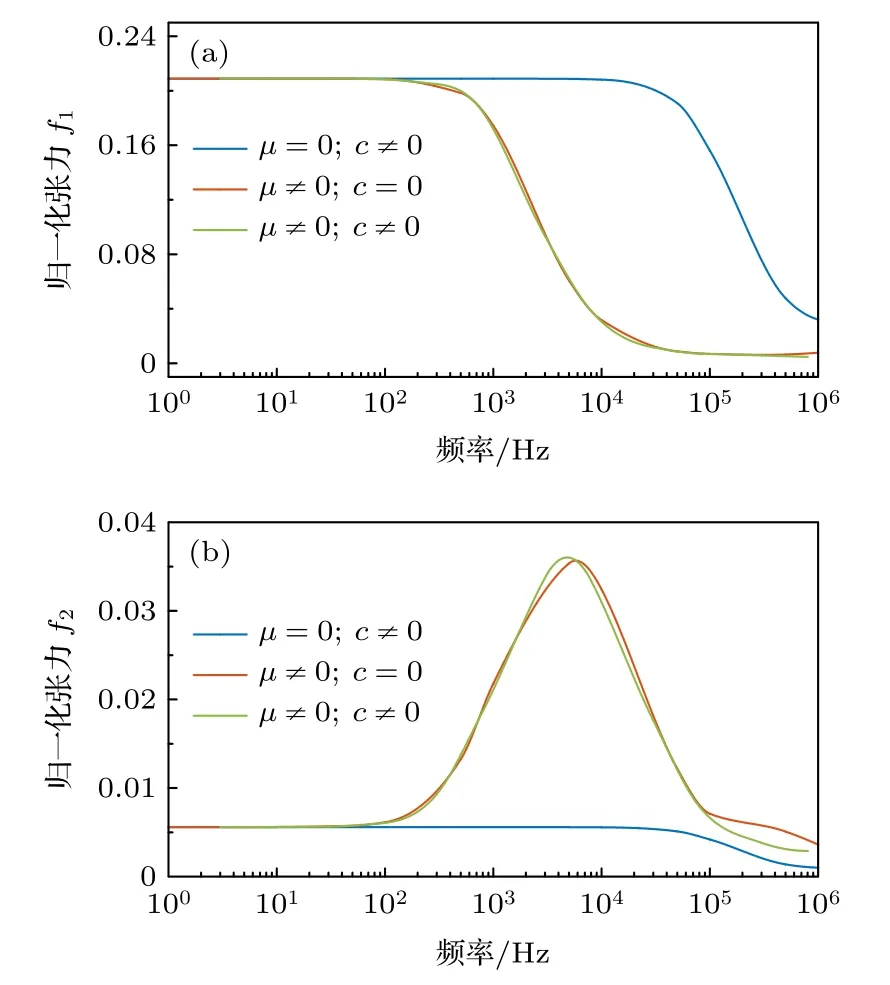
图7 不同流体作用方式下归一化张力随频率的变化曲线 (a) f1 ;(b) f2Fig.7.Variation curve of normalized tension with frequency under different fluid action modes:(a) f1 ;(b) f2 .
从图7 中可以看出,在不考虑黏滞阻力时(c=0),tip-link 张力基本不变,说明黏滞阻力对发束动力响应的影响不大.当不考虑流体压力时(μ=0),tip-link 归一化张力f1在更高的频率处迅速下降,即低通滤波的截止频率变大,但幅值变化不明显;f2在峰频附近不再出现类共振现象,峰频一定范围外的低频、高频域变化非常小.Kondrachuk[39]曾通过实验证明,周围流体环境产生的粘性阻力对静纤毛束的挠度影响不大.本文对可变形静纤毛的理论研究给出了与Kondrachuk 实验相似的结果,并进一步表明淋巴液压强的存在是产生tip-link 张力共振性质的必要条件.
3.5 相邻静纤毛间的流体压强
Kozlov 等[40]通过将高分辨率实验和数值模拟相结合的方法发现静纤毛紧密排列能有效地约束它们之间的流体.在低频时,流体动力耦合较弱,发束运动主要由tip-link 调节.高频时,流体动力耦合较强,运动主要受淋巴液调节.前文的计算结果显示,静纤毛间的淋巴液压强随时间动态变化,在特定范围内简谐波动,因此存在压强幅值.记高、中纤毛和中、短纤毛间压强幅值分别为和.将距离根部0.7 μm(短静纤毛的中点)和2.1 μm(中静纤毛高于短静纤毛部分的中点)的位置分别标记为A 和B,如图8(a)所示.计算在1—106Hz 频率范围内A,B 两高度处的最大压强,结果如图8(b)所示.从图8(b)可以看出,随着频率的增加,压强整体呈“阶梯”上升趋势.当频率超过100 Hz 时,压强幅值迅速增加,整体表现为在低频区域较小,高频区域较大.这说明随着盖膜剪切荷载频率的增加,淋巴液对静纤毛的作用越来越强,这一定程度上解释了tip-link 张力在高频区域急剧衰减的现象.本文计算的静纤毛间流体压强的理论变化与Kozlov 等[40]之前的实验结果一致.另外哺乳动物耳蜗中一些高频发束明显缺乏水平顶部连接器[41],这可能与淋巴对静纤毛的强烈限制有关.
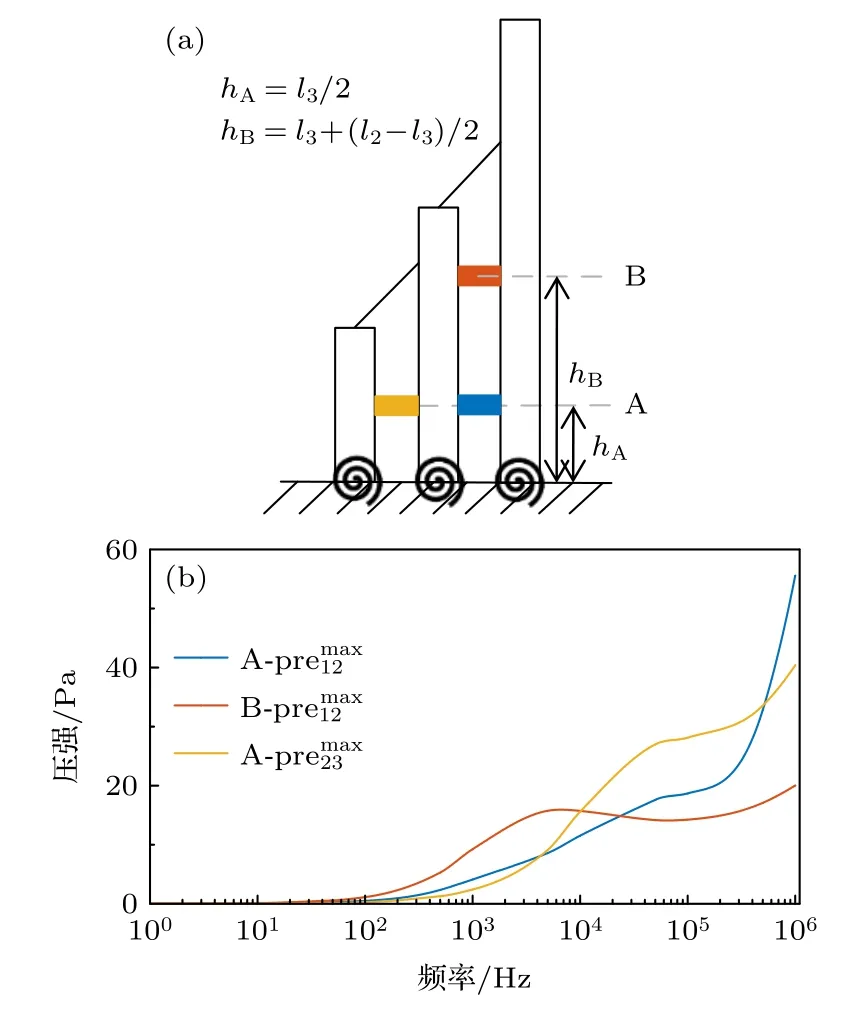
图8 A 和B 处的压强 (a) A 和B 在发束中的位置;(b) A和B 处压强随频率的变化曲线Fig.8.The pressure at A and B:(a) The position of A and B in the hair bundle;(b) the variation curve of pressure at A and B.
4 结论
本文将静纤毛视为吻合真实耳蜗中纤毛产生弯曲变形的工作状态的弹性体,推导出发束在简谐荷载作用下的运动解析方程,探索了静纤毛与淋巴液的耦合作用及运动规律,并扩展了泊肃叶流.将经典动力学与扩展泊肃叶流相结合,建立了流固耦合下的发束分布参数的理论解析模型.并进行了多个频域分析.通过分析,得出以下结论:
轴弯曲在静纤毛高度上产生位移的非线性积累,静纤毛越高这种积累作用越明显.在动力荷载作用下,高静纤毛轴弯曲对位移响应的贡献最大,且这一贡献容易受到频率变化的影响.低频荷载下,高静纤毛位移响应主要来源于根部偏转,高频率下高静纤毛轴弯曲明显变大,位移响应由轴弯曲和根部偏转共同产生.
实验发现暴露在噪声中的耳蜗,F-actin 含量的变化会影响静纤毛刚度.本文发现静纤毛杨氏模量减小会增加归一化张力f2的峰值,并降低其峰值频率,且低频盖膜剪切荷载下归一化张力的幅值均会增大.由于tip-link 与离子通道相连,归一化张力变化会影响离子通道开启的概率,改变耳蜗对相应频率声音的感知能力,进而影响发束的频率选择性.因此,以往将静纤毛作为刚体研究,会使得耳蜗对低频声音信号的响应被低估.以静纤毛作为变形体的模型能够比较准确的描述tip-link 张力与静纤毛运动规律,为噪声环境引发的听力障碍提供了相应的理论解释.
先前的实验表明,淋巴液黏滞阻力对静纤毛的挠度影响不大.忽略黏滞阻力时,tip-link 张力变化很小,忽略静纤毛间压强时,tip-link 张力明显变化,f2的共振峰消失,因此认为淋巴液通过在静纤毛之间产生压强的方式来调节tip-link 张力的共振性质.淋巴液的存在对发束频率特征的产生是至关重要的.静纤毛运动在低频域主要受tip-link 的调节,在高频域主要受淋巴液压强的调节.根据前文对静纤毛间压强的定义,该压强实际上是静纤毛振动引起的发束内外域空间的压强差.盖膜剪切荷载频率越高,头发束内外域空间的压强差就越大,这种压强差对静纤毛运动的调节作用有待深入研究.
