植物多酚-蛋白质复合物生物活性及应用研究进展
2022-03-03曲也直高雅然乌日娜武俊瑞
杨 慧,曲也直,高雅然,乌日娜,武俊瑞*
(沈阳农业大学食品学院,辽宁 沈阳 110000)
多酚类化合物是植物体内重要的次生代谢产物,主要存在于人们日常食用的大量果蔬中。目前,文献中已报道的多酚高达10 000多种。多酚具有显著的抗氧化、抑菌、抗过敏和抗衰老等功能,并作为天然色素的重要组成成分和抗氧化剂的主要来源,被广泛应用于食品和医药行业[1]。根据结构,多酚类化合物可以分为多酚单体和聚合多酚(图1)。其中,多酚单体根据其碳骨架特征又可划分为酚酸类、黄酮类化合物和少量1,2-二苯乙烯和木酚素,主要包括绿原酸、咖啡酸、异黄酮、黄酮和花青素等;聚合多酚为多酚单体的低聚或多聚物,统称单宁类物质,包括水解型和缩合型多酚,如没食子酸和原花青素等[2]。
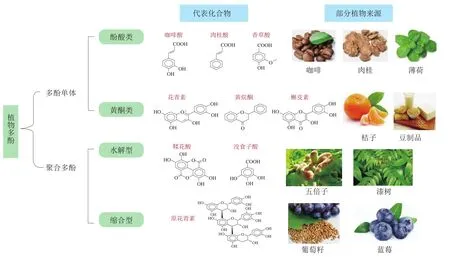
图1 植物多酚的分类及代表化合物[2]Fig.1 Classification and representative compounds of plant polyphenols[2]
蛋白质是乳类、肉类、蛋类、谷物类、豆类等食品体系中的重要组成成分,其功能性直接影响食品的特性。多酚-蛋白质共存现象在豆奶(大豆蛋白-黄酮)、奶茶(乳蛋白-茶多酚)、植物蛋白饮料(植物蛋白-植物多酚)等人们的日常饮食中普遍存在。研究表明,多酚类化合物具有独特的结构和理化性质,能在食品加工、储存、消化过程中与蛋白质发生相互作用,形成多酚-蛋白质复合物,进而改变或增强两者的功能以及生物利用度[3-4]。如表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)与牛乳中β-乳球蛋白(β-lactoglobulin,β-Lg)的复合物抑制人恶性黑色素瘤A375细胞和食管癌TE-1细胞活性的能力较单独的EGCG分别提高了69.0%和63.7%[5];茶多酚在泥鳅微冻贮藏过程中能显著改善其肌原纤维蛋白的表面疏水性及溶解度[6];姜黄素和鸡蛋卵白蛋白(ovalbumin,OVA)的复合作用不仅增强了姜黄素的抗氧化活性,还提高了其溶解度和光稳定性[7-8]。因此,多酚与蛋白质的复合作用被认为是有效改善蛋白质和多酚功能特性从而扩大其应用领域的一种安全、方便的策略。
本文主要从植物多酚与蛋白质的关系出发,阐述影响多酚与蛋白质相互作用的因素;同时重点总结了近几年多酚-蛋白质复合物的抗氧化性、消化性、致敏性和抑菌性等生物活性的研究进展及其在食品医药领域中的应用;并基于此对植物多酚-蛋白质复合物未来的研究方向进行了展望,以期为多酚和蛋白质的合理高效利用及扩大应用范围提供理论参考。
1 多酚-蛋白质相互作用
多酚与蛋白质通过相互作用可以形成多酚-蛋白质复合物,但不同类型多酚与不同蛋白质间的相互作用存在一定的差异[3]。影响多酚与蛋白质相互作用的因素主要分为多酚类型、蛋白质类型、化学结合类型和反应条件等(图2)。
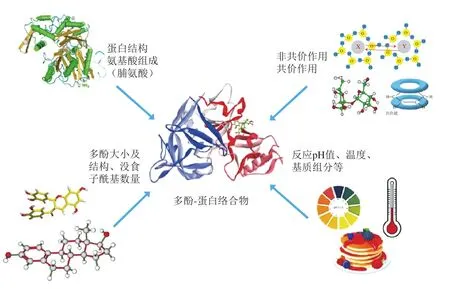
图2 影响多酚-蛋白质相互作用的主要因素[3]Fig.2 Main factors affecting polyphenol-protein interactions[3]
1.1 多酚类型的影响
多酚分子大小、没食子酰基数量和结构灵活性是影响多酚-蛋白质复合物形成的重要因素。具体表现为缩合单宁比水解单宁更易与蛋白质发生复合。Bohin等[9]研究发现原花青素中的黄烷-3-醇与牛血清白蛋白(bovine serum albumin,BSA)的相互作用随着其聚合度的增大而增强,而分子质量相近的没食子酸和阿魏酸与BSA的结合量相近,且简单酚类物质与蛋白质的结合能力较聚合程度高、分子质量大的多酚化合物弱。Dobreva等[10]通过对(+)-儿茶素、(-)-表儿茶素、(-)-表儿茶素没食子酸酯、苹果苷-3-葡萄糖苷、鞣酸、原花青素B4、没食子酸、原花青素B2和原花青素低聚物9 种多酚与BSA及α-淀粉酶的结合能力进行比较发现,多酚与蛋白质的相互作用还随其没食子酰基数量的增加而增强,这可能是由于没食子酰基含有可与蛋白质分子形成氢键和疏水作用力的苯环和自由羟基,从而增强了多酚与蛋白质的亲和力。此外,多酚分子的灵活性越高、自由没食子酰基数量越多,其与蛋白质的结合位点越多,亲和力越强。Frazier等[11]报道了多酚分子结构的灵活性会影响BSA与明胶结合多酚的数量,即多酚可以通过分子内旋转,从而更好地暴露结合位点,与蛋白质发生复合反应。Sekowski等[12]也指出构象变化能力有限的刚性鞣花单宁与蛋白质的相互作用往往较弱。
1.2 蛋白质类型的影响
蛋白质的氨基酸组成和结构也能显著影响多酚-蛋白质复合物的形成。蛋白质和多酚发生相互作用的一个必要条件为蛋白质/肽需要具有舒展的构象和相当大比例的脯氨酸,以便其形成松散的螺旋结构和疏水性斑块,从而与酚环结构相结合。其中,富含脯氨酸的蛋白质(proline-rich proteins,PRPs)通常就具有开放的随机卷曲或类似胶原的螺旋结构,更易与多酚产生氢键和疏水相互作用[13]。Amoako等[14]研究发现具有高弹性、多随机螺旋结构的蛋白质比结构紧凑折叠的蛋白质更易与多酚结合。根据等电点和糖基化程度可将PRPs分为3 类:酸性、碱性和糖基化PRPs,其中碱性PRPs比酸性和糖基化PRPs更易与多酚结合,大分子质量PRPs与多酚的亲和力较小分子质量PRPs强[15]。除脯氨酸外,富含精氨酸和芳香族氨基酸(如色氨酸和酪氨酸)的蛋白质也易与多酚通过疏水基团相互作用。Soares等[16]研究发现(+)-儿茶素与α-淀粉酶(富含脯氨酸和色氨酸)的亲和力高于球蛋白(BSA)。
1.3 化学结合类型的影响
多酚与蛋白质相互作用力一般分为非共价作用(疏水作用和氢键)与共价作用(共价键和离子键)[17]。但不同多酚与不同蛋白质的相互作用方式存在一定的差异。例如,多酚类化合物在碱性溶液中易氧化成相应的醌,醌是一种反应性亲电中间体,容易与蛋白质侧链中赖氨酸、蛋氨酸、半胱氨酸和色氨酸等亲核物质发生不可逆的共价结合;而蛋白质与多酚非共价结合主要由于疏水作用和氢键,且为可逆反应,其中疏水作用发生在蛋白质脂肪族、芳香族氨基酸与多酚的苯环之间,氢键产生在蛋白质的羰基(蛋白质氨基酸及肽键的羰基)与多酚的羟基之间。研究发现,原花青素中的酚羟基和疏水区可以分别与小麦谷蛋白的羰基和疏水氨基酸结合,β-Lg与染料木素和山柰酚结合过程中起主要作用的也是疏水相互作用力[18-19]。此外,蛋白质的H受体位点和多酚所携带的羟基之间的氢键在增强和稳定多酚-蛋白质复合物结构方面起重要作用[20]。值得注意的是,蛋白质与多酚的复合通常会同时存在不同程度的非共价和共价作用力,但主要是非共价作用力。
1.4 其他影响因素
溶液pH值、反应温度、离子浓度、食物基质成分等因素也会影响多酚-蛋白质相互作用,其中溶液pH值不仅能够改变多酚的化学结构和氧化聚合程度,还能改变蛋白质表面的静电荷分布、分子构象及溶解度,而反应温度主要通过影响氢键和疏水作用力,进而影响多酚-蛋白质相互作用。Prigent等[21]对单分子酚类化合物绿原酸(5-O-咖啡酰奎宁酸)与BSA、溶菌酶和α-乳白蛋白的非共价相互作用进行了表征,发现5-O-咖啡酰奎宁酸和BSA、溶菌酶的复合作用随着温度的升高而降低,且在酸性条件下,pH值对5-O-咖啡酰奎宁酸与BSA的复合作用没有显著影响,但随着pH值的增加,5-O-咖啡酰奎宁酸和溶菌酶间的复合作用增强,这可能是因为5-O-咖啡酰奎宁酸在碱性环境下自身氧化形成多个自由基或醌基,促进了多酚与蛋白质的共价结合。反应体系中高浓度的盐离子也会减少多酚与蛋白质的结合位点数量,从而影响其相互作用。Rawel等[22]研究发现,槲皮素与BSA的结合常数随溶剂中NaCl浓度的增加而降低。此外,食品基质中的多糖、脂质等成分也会与多酚反应从而减少多酚与蛋白质的结合位点,进而抑制多酚-蛋白质的相互作用[23]。
综上所述,多酚与蛋白质的相互作用受多酚类型(分子大小、没食子酰基数目和结构灵活性)、蛋白质类型(氨基酸组成和结构)、溶液pH值(蛋白质的电荷分布以及多酚稳定性)和反应温度(蛋白质构象和非共价结合力)影响较大。然而,实际研究中往往还有其他因素的影响,且不同种类多酚与蛋白质反应之间可能存在协同或拮抗作用。因此在食品生产实际应用中,根据蛋白质选择合适的多酚类型和反应条件,是保证其具有最佳生物活性的重要前提。
2 多酚-蛋白质复合物生物活性变化
多酚与蛋白质的相互作用会改变两者的功能和生物利用度,接下来重点归纳了近些年文献中报道的多酚-蛋白质复合物在抗氧化性、消化性、致敏性和抑菌性方面的变化。
2.1 抗氧化性
膳食多酚是人体抗氧化剂的主要来源,其抗氧化机制多为抑制参与生成过量活性氧类物质的促氧化酶活力、螯合过渡金属离子或者清除自由基等。如膳食黄酮类多酚可通过抑制还原型辅酶II发挥体内抗氧化剂的作用,还原型辅酶II被认为是血管壁中超氧阴离子的主要来源[24]。研究表明,多酚-蛋白质的相互作用对其复合物抗氧化性有正反两方面。一方面,多酚-蛋白质的相互作用对多酚清除自由基活性有掩蔽作用,降低了多酚的抗氧化潜能。Rohn等[25]研究发现槲皮素与BSA共价结合后,减少了蛋白质与酚羟基结合的某些必需氨基酸(赖氨酸的ε-氨基和半胱氨酸的硫醇基团)的数量,从而使复合物的抗氧化活性下降了79%。Medina等[26]证实了EGCG等茶多酚与乳铁蛋白的相互作用减少了反应体系中游离EGCG的含量从而降低了其抗氧化活性。Serafini等[27]也指出乳蛋白与巧克力中黄酮类多酚发生作用后,降低了黄酮的抗氧化活性。Xiao Jianbo等[28]研究发现膳食黄酮类多酚与乳蛋白的亲和力越强,其复合物1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力越弱,推测多酚类化合物与蛋白质结合会影响其自身的解离,从而降低其抗氧化性。另一方面,槲皮素与大豆分离蛋白(soy protein isolate,SPI)、α-生育酚与血浆蛋白的结合均提高了复合物的抗氧化活性,绿原酸与乳清分离蛋白的共价作用也显著增强了其抗氧化活性,且抗氧化活性同绿原酸结合量成正比,推测可能是多酚与蛋白质结合后,蛋白质的保护作用抑制了胃肠道中促氧化酶活性,使多酚顺利通过上消化道转运到结肠[29-31]。
2.2 消化性
在机体胃肠道中,多酚类化合物与蛋白质的相互作用也可以改变蛋白质的消化性,其相互作用对消化性的影响同样具有两面性。一方面,许多研究表明多酚会导致蛋白质的消化率下降。Rawel等[32]通过比较绿原酸-BSA复合物与天然BSA的胰蛋白酶消化率发现,绿原酸-BSA复合物经蛋白酶消化24 h的消化率等同于天然蛋白消化5 min,即多酚降低了蛋白质的消化率,这主要归因于其相互作用破坏了BSA中半胱氨酸、赖氨酸和色氨酸等必需氨基酸结构。Viva等[33]的研究指出多酚的加入降低了菜豆蛋白的溶解度,使其产生沉淀,进而降低了蛋白质的消化率。Zhang Yan等[34]通过模拟体外消化实验发现黑豆蛋白的水解度随着其总多酚含量升高而降低,这可能是由于多酚与蛋白质结合后,削弱了蛋白酶水解肽键的能力。类似的,鸡肉蛋白、鱼肉蛋白、牛肉蛋白与茶多酚结合后同样降低了其消化性[35]。另一方面,也有文献报道多酚能够改善蛋白质的消化率。Jiang Lianzhou等[36]研究指出黑米花青素与SPI的结合提高了蛋白质的消化率,消化率随着黑米花青素结合量的增加而增大,且消化产物具有较高的抗氧化性。儿茶素与β-伴大豆球蛋白的相互作用改变了蛋白质的二级结构,增加了其与消化酶的结合位点数量,进而提高了蛋白的消化率[37]。由于消化道是一个复杂的稳态内环境,目前对于多酚、蛋白质和消化酶类在人体消化环境中的相互作用及其对蛋白质消化率影响的作用机制仍需进一步探索。
2.3 致敏性
不同多酚在不同的反应条件下会对蛋白质二级结构中α-螺旋、β-折叠、β-转角和无规卷曲产生不同的影响。研究发现,茶多酚与牛乳酪蛋白的相互作用会使酪蛋白中的α-螺旋、β-折叠相对含量减少,无规卷曲结构相对含量增加,且结合力随着酚羟基数目的增加而增强[38]。此外,EGCG与SPI的相互作用也会导致SPI结构中的β-折叠向α-螺旋转换,且随着EGCG浓度的增大,这种转换作用加强[39]。蛋白质结构的变化可能会改变过敏原构象性表位的空间结构或者线性表位的组成,且这些变化均具有升高、降低甚至消除蛋白质致敏性的可能。换言之,多酚与致敏蛋白形成复合物可能导致后者结构改变,从而间接调节过敏反应。现有文献报道牛乳中乳清蛋白和鸡蛋中的OVA均可与绿原酸相互作用,通过改变蛋白的二级结构,破坏其构象性表位或线性表位组成,降低牛乳和鸡蛋致敏性[40-41]。姜黄素、苹果多酚与OVA的复合作用也可通过抑制辅助型T细胞2(T helper 2 cell,TH2)型相关细胞因子白细胞介素(interleukin,IL)-4、IL-5和IL-13的分泌来有效减弱其过敏反应症状[42-43]。此外,OVA也可与儿茶素、槲皮素共价结合来降低鸡蛋的致敏性[44-45]。进一步研究发现,儿茶素还可以有效降低人嗜碱性粒细胞表面免疫球蛋白(immunoglobulins,IgE)高亲和力受体FcɛRI mRNA的表达,从而抑制效应细胞发生脱颗粒和组胺释放,从而达到缓解过敏反应的效果[46]。类似的,海洋来源的活性物质褐藻多酚也是通过此机制降低了花生过敏原(Ara h 1、Ara h 2、Ara h 3、Ara h 6)的致敏性[47]。Plundrich等[48]利用化学信息学方法研究了越桔果实中42 种不同多酚类物质与花生过敏原Ara h 2结合并抑制其免疫球蛋白IgE结合表位的可能性,免疫印迹结果显示花青素和绿原酸与Ara h 2作用后,其致敏性分别下降了37%和50%。Tantoush等[49]也指出绿茶多酚与β-Lg、Ara h 1和Ara h 2可形成易肠胃消化降解的复合物,从而导致其致敏性下降。值得注意的是,非黄酮类多酚化合物白藜芦醇与OVA的结合暴露了蛋白内部的IgG和IgE结合位点,使得OVA的潜在致敏性增强[50]。综上所述,多酚-蛋白质复合物致敏性降低多是由于其结构变化导致表位被破坏,而有关致敏蛋白与多酚的结合位点及作用力类型、复合物对免疫细胞的影响等报道较少。
2.4 抑菌性
植物多酚作为新型生物保鲜剂,因其来源广泛、作用温和、抑菌效果好和不良副作用较少而备受关注,常被应用于肉制品、水产品和果蔬制品。现有文献表明,多酚与蛋白质的相互作用会影响其抑菌效果。Fu Shalu等[51]研究发现绿原酸-明胶的复合物相较于绿原酸,对大肠杆菌、铜绿假单胞菌、单核细胞增生李斯特菌和葡萄球菌活性的抑制作用显著增强。迷迭香酸与乳清蛋白共价结合后的复合物也具有显著的抗金黄色葡萄球菌活性,而当乳清蛋白单独作用时,并不能抑制任何细菌的生长,这可能取决于乳清蛋白反应基质中多酚生物活性的有效性[52]。此外,在豆奶发酵过程中添加茶多酚可以有效促进保加利亚乳杆菌的生长,抑制嗜酸乳杆菌和嗜热链球菌的繁殖[53]。ben Arfa等[54]通过向SPI中添加一定量的肉桂醛和香芹酚,发现其复合物会对灰霉病菌、大肠杆菌的生长具有抑制作用。综上所述,多酚与蛋白质的复合物通常具有良好的抑菌特性,其抑菌机制可能是改变了微生物细胞膜的通透性,并影响其细胞内某些酶的表达,或破坏其细胞壁等[55]。然而,不可忽视的是,目前多酚-蛋白质复合物还存在着抑菌范围小、抑菌效果弱等问题。
近年来文献中报道的多酚-蛋白质复合物在抗氧化、消化性、致敏性和抑菌性方面的变化总结如表1所示。

表1 多酚-蛋白质复合物生物活性变化Table 1 Biological activities of polyphenol-protein complexes
总体而言,多酚与蛋白质的相互作用对两者的功能和生物利用度都具有显著的影响,尤其是在抗氧化性、消化性、致敏性和抑菌性方面。然而,现有的多酚-蛋白质复合物的生物活性测定多是基于体外测定,且活性差异与结构改变、机体肠道菌群变化的内在关联还不太清楚,需在今后进一步研究。
3 多酚-蛋白质复合物的应用
多酚-蛋白质复合物由于特有的结构和生物活性,可以被制备为功能性乳液、脂质体、微胶囊、纳米颗粒等新型复合体,还可以作为金属纳米粒子的稳定剂[56]。本文就近年来关于多酚-蛋白质复合物应用的文献进行分析总结,重点综述了其在新型抗氧化乳化剂、可食用膜及药物缓释载体方面的应用。
3.1 新型抗氧化乳化剂
研究发现,多酚与蛋白质的复合物作为新型抗氧化乳化剂,不仅具有良好的乳化性能,还可以有效保护易氧化食物基质中的活性物质[57]。Yi Jiang等[58-59]研究发现儿茶素-β-Lg复合物和儿茶素-α-乳球蛋白复合物均可作为乳化剂包埋β-胡萝卜素,且能够有效抑制β-胡萝卜素发生氧化降解。绿原酸-乳铁蛋白、EGCG-乳清蛋白复合物也可以显著提高乳化液中β-胡萝卜素的化学稳定性[60-61]。Feng Jin等[62]研究指出儿茶素-OVA复合物包埋的鱼油乳液具有粒径小、贮存稳定性好、黏度小等特点,且复合物具有较高的抗氧化活性和界面活性,对鱼油乳液中脂质氧化的抑制作用优于OVA。类似的,儿茶素-蛋清蛋白复合物不仅具有较高的乳液稳定性,还具有较强的抗氧化活性和界面活性[63]。儿茶素-米糠蛋白复合物形成的稳定水包油乳液具有较好的黏性及黏弹性[64]。此外,大米分离蛋白与阿魏酸的复合物具有稳定的乳液稳定性,且可以通过降低过氧化氢、三聚氰胺和己醛的浓度,有效抑制脂肪氧化降解[65]。Wei Zihao等[66]通过EGCG共价修饰不同乳蛋白(α-乳白蛋白、β-Lg、乳铁蛋白、酪蛋白),发现不同多酚-蛋白质复合物的乳化性质取决于所使用的酚类化合物和蛋白质的类型。目前,有关多酚-蛋白质复合物作为新型抗氧化乳化剂的科学报道还比较少,已有文献中蛋白的种类主要集中在乳源蛋白。此外,环境应力是否会对多酚-蛋白质复合物乳液的冻融稳定性、盐稳定性和热稳定性产生影响,以及复合物的乳化机制、毒性、生物利用率等仍需进一步详细探究。
3.2 可食用膜
近年来,蛋白膜由于其对环境的友好性和可生物降解性,成为食品保鲜行业的研究热点,但易滋生细菌、机械强度低和水蒸气阻隔性能弱等缺点限制了其应用。研究表明,植物多酚可以作为蛋白质交联剂添加到可食用膜中,不仅能提高其机械强度和阻隔性能,还能抑制菌活性、减缓食品氧化,达到延长保质期的效果[67]。Araghi等[68]将鱼明胶与咖啡酸复配成鱼明胶膜,发现咖啡酸提高了膜的水蒸气透过性和氧气透过性。同样的,玉米蛋白可食用膜中单宁酸的添加使其具有更高的抗拉强度和较低的水蒸气透过率[69]。Choi等[70]通过测定不同氧化酚类物质(包括鞣酸、咖啡酸和绿茶提取物)对明胶-姜黄素膜机械屏障性能的影响,发现氧化酚醛具有的交联作用增强了膜的拉伸强度,降低了其断裂伸长率、水蒸气透过率和水溶解度;在不同种类膜中,咖啡酸-明胶-姜黄素膜具有最优的机械和屏障性能;且这些膜均具有较强的抗氧化活性,可作为活性包装材料来延长新鲜猪肉的货架期。绿茶多酚提取物对明胶膜的机械强度、阻隔性也具有增强作用,且体外抗氧化能力(DPPH自由基清除能力、铁还原能力)随多酚添加量的加大而升高[71]。此外,向大豆蛋白膜中添加芦丁可以有效降低水蒸气的透过性,这可能是由于芦丁与大豆蛋白的相互作用导致膜的结构更紧密,降低了膜基质中水分子流动性并限制水通过膜迁移[72]。同样,向大豆蛋白膜中添加丁香酚也可以增加其水分子阻力,从而降低水蒸气透过性、透气性和透油性;在实际应用中,丁香酚-大豆蛋白膜还能起到抑制圣女果淀粉降解和果胶质转化、减缓其软化速度的作用[73]。不可忽视的是,目前多酚-蛋白质可食用膜的应用还受到诸多限制,如植物多酚的添加会使膜的厚度增加,且多酚自身的颜色和气味也会扩散到食品中等。此外,在成膜过程中,植物多酚与蛋白质间的相互作用尚不完全清楚,仍有研究空间。
3.3 药物缓释载体
由于多酚-蛋白质复合物不仅具有良好的抗氧化性和稳定性,还具有较大的负载面积、易制备和较好的组织黏性等优点,常在医药领域被用作多种药物控制输送的载体。Liu Fuguo等[74]研究发现,基于EGCG-玉米蛋白复合的纳米颗粒不仅可以保护姜黄素和白藜芦醇不被降解,还能够保持其抗氧化活性,提高其生物可及性。绿原酸-β-Lg复合的纳米颗粒同样可以保护EGCG不被降解,还能控制EGCG在模拟胃肠消化液中的释放速率[75]。Fan Yuting等[76]的研究指出咖啡酸-BSA偶联/玉米蛋白纳米颗粒对白藜芦醇的降解有明显的保护作用,且能够显著提高白藜芦醇的热稳定性、紫外光稳定性和抗氧化活性。此外,姜黄素-高粱醇溶蛋白纳米颗粒膜也可以提高口服型药剂的稳定性[77]。值得注意的是,酚类化合物不仅能稳定蛋白水凝胶的结构,还能刺激水凝胶抑制有害酶活性和细菌生长,从而促进伤口愈合,且复合物的凝胶性质与共价连接的酚类物质的数量密切相关[78]。Hu Bing等[79]首次发现,在高浓度蛋白淀粉样纤维的条件下,多酚类小分子化合物够通过疏水作用、π-π共轭作用、氢键作用吸附到球蛋白表面,驱动处于液晶态的纳米纤维超分子自组装,在空间形成饱含水分的三维立体网状多层次结构水凝胶,且多酚类物质的载药量显著增加(质量分数达到4.0%),稳定性得到明显改善,并具有广谱抗菌作用。然而,多酚-蛋白质复合物在药物缓释载体应用中也存在一些问题,例如载药量和载药效率较低、药物爆发性释放等,尚有提高空间。
近年来关于多酚-蛋白质复合物的应用研究如表2所示。

表2 多酚-蛋白质复合物的应用Table 2 Applications of polyphenol-protein complexes
综上所述,多酚-蛋白质复合物在食品和医药生产领域具有很高的应用价值,但目前大多数报道集中于其机理和结构的表征研究,而对多酚-蛋白质复合物的实际应用研究较少,且复合物的生物安全性仍有待商榷。值得注意的是,采用蛋白质-多糖-多酚相互作用形成的三元复合物也是近年来的新型材料研究热点,它具备了3 种物质的功能特性,更适宜于不同食品体系中的应用。
4 结 语
植物多酚与食物蛋白质间主要是非共价疏水相互作用,然后通过氢键稳定结构,并受蛋白质类型(蛋白质大小、构象、氨基酸组成)、多酚类型(大小、结构灵活性、没食子酰基数量)及反应条件(pH值、温度、离子浓度)等的影响。然而目前大多数研究都倾向于多酚在高温或强碱条件下共价修饰蛋白质侧链氨基酸,生成稳定的复合物,以便符合食品加工业的要求,但强烈的反应条件易引起食品中其他营养元素损失等问题。此外,不同种类多酚与蛋白质反应体系中是否存在协同或拮抗效应,食品新技术(超声、等离子体、静高压等)的应用是否会影响多酚-蛋白质间的作用力,仍待进一步研究。
多酚-蛋白质复合物在很大程度上保持了植物多酚的化学性质,同时赋予蛋白质分子许多新的功能。然而,现有的多酚-蛋白质复合物种类十分有限,研究重点集中于抗氧化、消化、抗过敏和抑菌等活性方面,但其他生物活性(如抗糖尿病、抗癌细胞增殖活性、保肝活性等)探索较少。值得注意的是,现有的多酚-蛋白质复合物的生物活性测定多是基于体外测定,且活性差异与结构改变、机体肠道菌群变化的内在关联还不太清楚,需要进一步探讨。同时,还应详细研究蛋白质与多酚相互作用的最适反应条件,以优化食品加工过程,最大限度地提高食品的营养和功能特性,增强其在食品加工过程或消化条件下的稳定性。
多酚-蛋白质复合物在食品和医药领域中有很好的应用前景,尤其在新型抗氧化乳化剂、可食用膜及药物缓释载体方面的应用。但是,植物多酚的提取难度大、化学结构存在多样性,且多数研究仅关注单一多酚或者单一配合物的使用,不仅实际应用研究较少,还缺少其生物安全性的评估。今后需加强多酚-蛋白质复合物体外细胞培养和体内动物毒理方面的研究,以探索多酚-蛋白质复合物材料的潜在毒性,并加强蛋白质与多酚复合物的性质研究,开发新型功能材料。
