瓜果类蔬菜果皮厚度发育调控研究进展
2022-02-28龚成胜王述彬刘金兵潘宝贵郭广君刁卫平
龚成胜 王述彬 刘金兵 潘宝贵 郭广君 刁卫平
(江苏省农业科学院蔬菜研究所/江苏省高效园艺作物遗传改良重点实验室,南京 210014)
引言
我国是世界上最大的蔬菜生产国、消费国和出口国,蔬菜产业的稳步健康发展有助于保障民生、保证粮食安全、促进国家经济发展。“八五”科技攻关以来,“优质、高抗、高产”成为了蔬菜作物重要的育种目标,并取得了显著成效。在瓜果类蔬菜优质育种目标中,果皮性状是一个重要的评定指标。果皮通常指的是果实的外皮或外壳,果皮厚度是与产量、品质及耐存储性等密切相关的重要农艺性状。通过与产量的相关分析,发现果皮厚度与西瓜、辣椒等瓜菜作物产量具有密切相关性,果皮越厚果实的产量也相对更高[1-4]。另外,果皮厚度与蔬菜品质紧密相关,果皮中富含糖类、有机酸、氨基酸、类黄酮等丰富的代谢物[5],能够为人体提供丰富的营养物质,且辣椒等蔬菜作物由于具有较厚的果皮而减少了农药的残留[6]。此外,蔬菜果皮厚度与耐储存性之间多存在正相关关系,如番茄的果皮厚度与外观、手感、裂果性以及耐储运特性之间有紧密联系[7-8];西瓜果实的厚度与硬度和裂应力之间具有正相关性[9];外果皮越厚的加工番茄在放置一段时间后的腐烂率相对越低[10]。
随着重要蔬菜基因组测序以及重测序工作的推进,生物技术研究使蔬菜育种进入‘高速’时代[11],并获得了多个与目标性状连锁的关键QTL[12]。通过分子标记辅助育种,开发出与西瓜果皮颜色、黄瓜果肉厚度等重要性状连锁的分子标记,能够在实验室实现对目标性状的快速鉴定,节约了成本并大大提高了育种效率[13-14]。同样,全面了解果皮厚度性状的遗传基础,并利用现代分子手段对蔬菜果皮厚度目标性状快速筛选,可为优质品种改良奠定基础。
近年来随着消费水平的提高,人们对具有适宜果皮厚度的优质蔬菜品种的需求在增加。不同蔬菜品种由于食用部位等方面的差异,往往对果皮的要求也存在差异。比如果皮较厚的西瓜在增加耐裂性的同时,可能会减少果实可食用部分以及改变风味品质;加工为主的辣椒多具有较薄的果皮厚度,而鲜食类型的甜椒品种的果皮厚度较厚。因而,基于实际生产需求的不同,生产合适果皮厚度的专用瓜菜新品种才能实现品种的多元化、特色化、优质化。全面深入开展果皮厚度形成的的生理机理及分子遗传基础研究,并通过与目标性状连锁分子标记的挖掘及利用基因编辑等技术实现基因的定点改造,有助于加速适宜果皮厚度的瓜果类蔬菜新品种的选育和生产。
1 控制瓜果类蔬菜果皮厚度的关键因素
1.1 环境因素
光照、温度、水分、肥料等因子是蔬菜作物正常生长发育的环境因素,通过改善这些环境因素可以对果蔬果皮厚度产生影响。优化光照强度和光照质量有助于改善作物的农艺性状,程群柱等[15]研究发现,当日平均温度 19 ℃,日平均光照强度 504 μmol/(m2·s)处理时,果皮厚度和果实质量等农艺性状要显著高于其他处理;在肥料喷施的研究中发现,喷施诺普丰叶面肥和硒肥能够减少西瓜果皮的厚度[16-17];而灌溉量和灌溉方式等水分管理措施对西瓜、辣椒和甜瓜果皮厚度的影响较小[18-20]。因此,可以通过改变环境条件优化果皮厚度,实现栽培管理措施对果皮厚度的有效调控。
1.2 外源化学物质
近年来,外源化学物质已被广泛的应用于作物生长发育的调控。植物生长调节剂同天然激素一样,具有调控生长发育的作用,同样可对果皮厚度产生影响;如外源喷施GA3增加了辣椒的果皮厚度,对果实的产量和品质产生了一定的影响[21];喷施适宜浓度的α-萘乙酸钠(α-NaNAA)显著增加了脱落酸、赤霉素和玉米素的含量,且有助于辣椒果皮厚的增加[22]。另外,化学诱变剂调控果皮厚度的研究也取得了一系列进展,适宜浓度的CaCl2处理能够增加外果皮细胞壁的厚度,提高番茄果实的硬度[23];化学试剂甲基磺酸乙酯(EMS)能够诱导番茄果实大小和果皮厚度的变化[24]。随着化学物质对植株生长发育调控应用的日益广泛,挖掘更多能诱导果皮产生差异的化学物质,定向改变果皮的厚度和产量,生产符合市场需求的果蔬。
1.3 遗传调控
遗传因素是导致果皮厚度差异的最关键因素。分析生物信息发现,番茄含糖量和果皮厚度的主分量占据总遗传信息量的80.96%[25]。辣椒果皮厚度具有较高的遗传和表型变异系数,说明性状的遗传范围广[26]。此外,刘仲齐等[8]研究发现,具有易裂基因型的番茄果皮相对更薄。易丽聪等[27]利用SNP 标记将64 份西瓜材料划分为4 个亚群,发现果皮厚度是区分不同亚群西瓜的关键指标。果皮厚度作为重要的数量遗传性状,其遗传因素是造成果蔬果皮厚度形成的关键调控因子。
2 蔬菜果皮厚度的细胞结构研究进展
果皮主要包括外果皮、中果皮和内果皮三部分[28]。外果皮具有角质层和气孔,通常由1~2 层细胞组成;中果皮主要由相对较大的薄壁细胞组成;内果皮位于最里层,细胞相对较小且多由多层石细胞构成。对果皮细胞结构的研究有助于阐释果皮厚度形成的生物学发育过程,可为果皮的细胞发育和遗传调控提供理论研究参考。
细胞层数和细胞大小等都是造成果皮厚度差异的生理基础。在蔬菜作物中,细胞结构差异往往与其生物学功能相适应。研究发现,不同物种、不同品种以及不同倍性的瓜菜果皮厚度及细胞结构均存在差异。加工番茄的果皮由外果皮、中果皮和内果皮三部分组成,薄壁细胞形态由外果皮到内果皮由圆变方,内果皮细胞以小而紧密排列的形式存在[29]。在具有较厚果皮的加工黄瓜品种中发现[30],其表皮细胞小且排列紧密,薄壁细胞由小到大的过渡较为缓慢,而果皮较薄的品种基本上无明显的表皮层。刘春香等[31]研究发现,黄瓜表皮细胞的厚度与果实硬度之间具有极显著相关性,并提出选育合适表皮厚度的黄瓜新品种既能提升感官评价,又能够增加果实的耐储运特性。詹园凤等[9]研究发现,厚果皮的西瓜品种果皮的细胞排列致密且形成了清晰的细胞团结构,而果皮薄的西瓜品种的表皮组织则具有更长的表皮细胞以及更大的外果皮和中果皮细胞。另外,果皮的厚度与果实的裂应力以及果实的硬度之间呈现显著相关性,在抗裂型西瓜表皮存在从小细胞到大细胞的转变[32]。刘鹏等[33]研究发现,多倍体西瓜的果皮厚度要显著大于其同源二倍体西瓜,这种差异性与表皮中的石细胞等组织细胞的形态结构有密切关系。
3 遗传基础研究进展
3.1 蔬菜果皮厚度的遗传分析
不论是自然群体还是分离群体,果皮厚度的分布规律研究发现该性状是典型的数量性状。如在自然群体中,番茄的果皮厚度呈正态分布[34];在F2S7重组自交系群体中,西瓜果皮厚度总体上呈正态分布,为典型的数量性状[35];在F2分离群体中,甜瓜的果皮厚度呈现正态分布[36]。
在经典遗传学中,数量性状的形成受到基因型和环境的共同调控[37],开展遗传分析是从基因层面解析果皮厚度形成的基础。果皮厚度是连续变异的数量性状,并受基因调控。在对西瓜果皮厚度的遗传分析中发现,其西瓜果皮厚度遗传符合C-0 模型,具有正向的加性和显性效应,且多基因上位性效应累计也表现为正向,为典型多基因控制的数量性状[38];江海坤[39]通过孟德尔遗传对西瓜(L-103 和X-101)果皮绿皮皮层的厚度进行遗传分析,发现该性状主要受1 对显性等位基因控制,而通过数量性状主基因+多基因遗传分析则认为该性状受2 对加性-显性-上位性主基因和加性-显性多基因控制。因此,尽管果皮厚度在不同样本中的遗传模型存在一定的差异,但分析结果表明果皮厚度的形成可能受到多个基因的调控且可能受到主基因的控制。
3.2 基于正向遗传学进行基因定位研究
随着宏基因组测序以及重测序工作的完成,为蔬菜作物重要农艺性状的多样性研究提供了丰富的遗传变异信息[40],基于正向遗传学手段解析蔬菜果皮厚度的研究已经取得了一定的进展(表1)。全基因组关联分析(GWAS)、遗传图谱和集团分离分析法(BSA)是最为常见的基因定位方法,且前两种方法更适合对数量性状进行基因定位研究。全基因组关联分析主要应用于对自然群体中目标性状的基因定位,并搭建起表型与基因型之间的桥梁[41]。袁欣捷等[42]以194 份辣椒为研究材料,在2、4、8、10 号染色体上获得了与果肉厚度关联的SSR 标记,且最高贡献率达到20.48%,为辣椒优异基因的挖掘与分子标记辅助育种奠定了理论支撑。LEE 等[43]结合GWAS 和RIL 群体构建的遗传图谱获得了与辣椒果实相关的主要驯化性状(果实长度、宽度、质量、果皮厚度和果实位置)的关联位点,并完成了候选基因的预测。Xie 等[44]在完成了对冬瓜基因组测序的基础上,也通过全基因组关联分析获得了在2、3、11 和12 号染色体上与果皮厚度关联的SNP 位点。Kim 等[34]对162 份番茄种质资源的果实性状进行全基因组关联分析,并在2、9、12 号染色体上获得了果皮厚度显著关联位点。
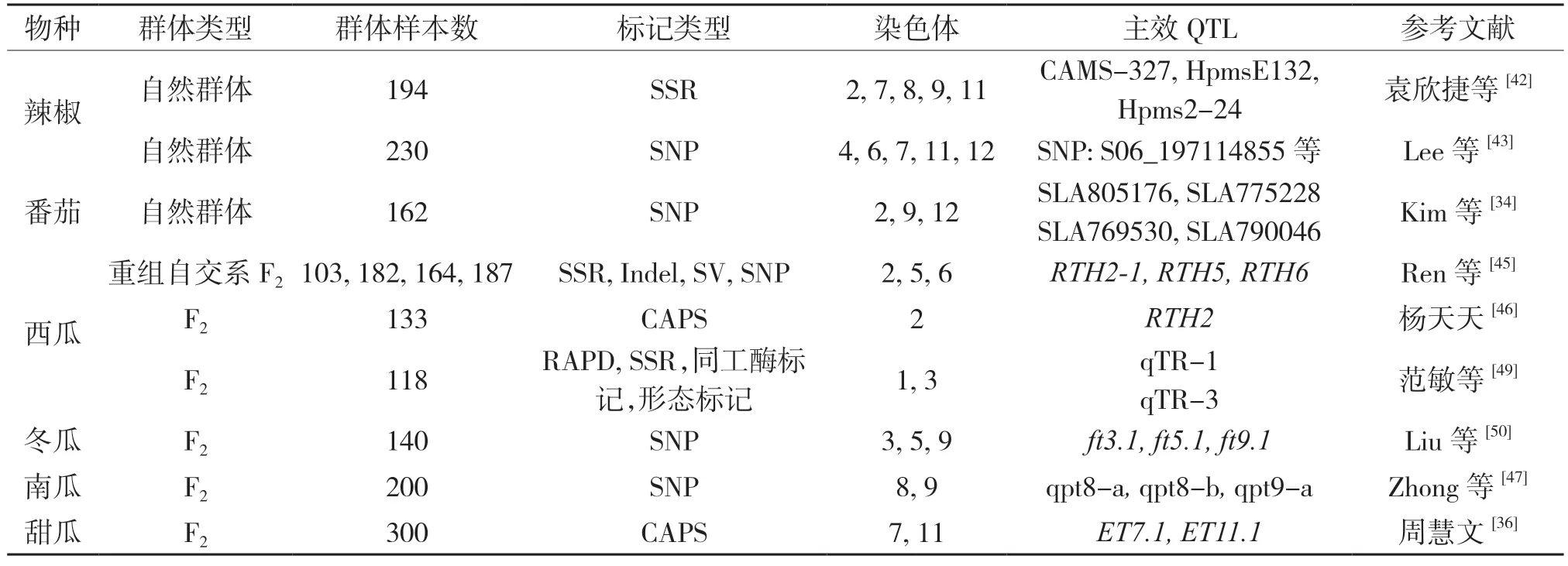
表1 近年来部分瓜果蔬菜果皮厚度基因定位信息
在分离群体中,利用遗传图谱对果皮厚度等数量性状的基因定位提供了有效工具,并在蔬菜作物果皮厚度的基因定位研究中取得了进展。如Ren等[45]通过整合4 个遗传图谱,实现了西瓜果实长度、糖含量等12 个农艺性状的关键QTL 位点,包括位处2、5、6 号染色体上与果皮厚度相关的QTL 位点RTH2-1、RTH5、RTH6。杨天天[46]基于遗传图谱也在2 号染色体上获得了一个与西瓜果皮厚度相关的QTL 位点RTH2。Zhong 等[47]完成了对南瓜高密度遗传图谱的构建,并获得了与果皮厚度等12 个重要农艺性状连锁的QTL 位点,为候选基因的挖掘与分子标记育种提供了理论支撑。在冬瓜、甜瓜等作物中也获得了与果皮厚度相关联的关键QTL 位点[36,48-50]。
3.3 关键基因调控果皮厚度的形成
基因对大田作物如玉米籽粒种皮厚起到重要的调控作用[51],也调控树木的增粗[52]。关键基因同样对园艺作物的果皮厚度起到重要的调控作用。果皮厚度往往和细胞的分裂和扩张密切相关,调控细胞扩张的结构基因在调控果皮厚度形成的进程中起到了重要的调控作用。在番茄细胞的复制与分化研究中,SlCCS52A基因表达水平与番茄细胞的核内复制特异相关,该基因的下调导致番茄细胞减小,果实尺寸较小[53];Musseau 等[54]结合细胞水平、转录水平以及代谢水平的数据,发现SlGBP1控制细胞的增殖和细胞分化,在调节细胞凋亡的进程中发挥重要作用,该基因的突变导致番茄的中果皮细胞变薄。在细胞扩张的研究中,Mori 等[55]研究发现SlBIM1a过表达的番茄能够导致果皮细胞变小,而SlBIM1a沉默的植株的果皮细胞变大,且SlBIM1a能够与SlBZH1相互作用来影响BR 靶基因的转录调控,进而表明SlBIM1a是果皮细胞扩张的关键调控因子;Swinnen 等[56]研究发现SlKIX8和SlKIX9功能的缺失导致番茄细胞的扩张以及果皮厚度的增加;CDKA1的过表达能够通过调控番茄细胞的大小,改变果皮的厚度[57]。
此外,转录因子能够通过控制结构基因改变细胞大小及果皮厚度。如bHLH 家族转录因子基因SlPRE2的沉默导致番茄果皮厚度和果实大小等性状的减小[58];SlARF7的mRNA 水平降低转基因番茄的细胞扩张,并形成了较厚的果皮[59];RNA 干扰(RNAi)策略沉默Sl-IAA17产生比野生型番茄更大的果实,结合果实器官组织学分析表明这种表型的差异与较厚的果皮有关[60]。
4 果皮厚度的研究应用与发展前景
果皮厚度作为反映蔬菜果实品质的重要指标,在辅助预测杂种优势以及辅助群体种质资源划分等方面得到了研究应用。在辅助预测杂种优势方面,由于果实质量和果皮厚度是育种中两个重要的农艺性状,Bahari 等[61]通过计算瓜皮厚度、果实质量等农艺性状的一般配合力(GCA)和特殊配合力(SCA),筛选了适合商业栽培的西瓜杂交组合。在辅助蔬菜种质资源的划分研究方面,陈萍等[62]研究发现,果皮厚度在119 份西瓜种质中的变异系数较大,为31.4%,并根据果实质量、果皮厚度以及含糖量这三个重要的农艺性状将这些种质划分为了10 类,为种质资源的创新与利用提供了理论参考。
随着特色专用型果蔬品种需求的增多,立足产业实际发展需求,培育更符合市场需求的果蔬新品种势在必行。表型特征的形成被认为是环境与基因相互作用的结果,通过环境因素改变性状、加速育种进程约在150 年前已有相关的研究[63],积极探究适宜的光照、温度、水肥等事宜条件,可缩短传代周期,加速新品种选育。另外,在明确果皮厚度结构的基础上,可以深入挖掘与果皮厚度形成相关联的其他农艺性状和代谢物等,既有望实现对多个关联性状的改良,加速聚合育种技术发展;反之亦然,又可以通过对关联代谢物的内源调控或者外源喷施,实现对果皮厚度的定向调控。
瓜果类蔬菜果皮厚度的分子调控研究中,目前缺乏有效的分子标记,以及难以精准定位调控主基因,因而需要通过加强正向遗传学手段对蔬菜果皮厚度进行基因定位。笔者认为在基因定位研究方面,既可以扩大遗传作图群体,并结合自然群体和分离群体进行多群体分析以获得更多的遗传变异信息,进而明确果皮厚度的遗传规律;也可以通过重测序技术构建高密度遗传图谱、整合图谱,并结合GWAS、BSA 等定位方法实现对目标性状的定位;可通过结合多组学(基因组、转录组、蛋白组等)数据,挖掘确定调控目标性状差异的关键候选基因[64];针对有效分子标记较少的问题,需要通过精细定位缩小候选区间,挖掘与性状关联或者开发连锁的有效标记,并进一步结合基因编辑、转基因等技术鉴定候选基因的功能,培育符合市场需求的新品种。
果皮厚度作为与产量、品质、抗性等相关的农艺性状,具有重要的研究应用价值。在未来,随着蔬菜果皮厚度在生理生化代谢、遗传调控基础以及分子辅助育种等方面深入研究,果皮厚度性状的生物学机理将会更加清晰,可用于实现果皮厚度人工栽培调控。
