低蛋白水平日粮添加复合酸化剂对断奶仔猪后肠微生物的影响
2022-02-28王同振刘丽晓郝瑞荣李清宏
王同振,徐 荣,刘丽晓,郝瑞荣,李清宏
(山西农业大学 动物科学学院,山西 太谷 030801)
肠道内栖息着数量庞大且代谢活跃的微生物菌群,构成了一个复杂的生态系统。肠道菌群的数量和结构对宿主的营养代谢和健康免疫起到重要的作用,影响肠道菌群变化的因素主要包括宿主自身健康状况、饮食结构和生活方式等。研究表明,日粮是引起肠道微生物数量和组成变化的主要因素,其作用方式更为直接迅速。而日粮中的蛋白质作为后肠微生物发酵利用的营养物质,对肠道微生物区系的改变更为明显。
早期断奶仔猪由于自身消化系统和免疫系统发育不完善,加之饲粮、环境和心理等多种因素改变容易引起断奶应激,造成仔猪肠道微生物区系紊乱、免疫功能降低,仔猪极易出现腹泻、生长迟缓甚至死亡。腹泻是导致断奶仔猪死亡的重要原因,以往通过在饲料中添加大量的抗生素以预防仔猪腹泻,加强仔猪抗病能力。当前在饲用抗生素禁用的背景下,通过营养调控手段提高动物生长性能和肠道健康水平已成为大势所趋。研究表明,低蛋白水平日粮对断奶仔猪生长性能没有显著影响但可以降低仔猪腹泻率,减少毒性物质(如氨和胺类)对肠道的损伤,促进肠道健康;复合酸化剂(Compound acidifier,CA)可以降低胃肠道pH,刺激消化酶活性、提高营养物质的消化率,改善肠道菌群结构,抑制有害微生物定植,促进有益菌的生长。目前,对于低蛋白水平日粮和CA对断奶仔猪肠道健康影响的研究绝大部分是单独进行的,而对两者的配合使用鲜有报道。因此,本试验利用高通量测序技术(Illumina MiSeq)和液相色谱串联质谱(LC-MS/MS)方法探究低蛋白水平日粮添加CA对仔猪后肠微生物菌群及其代谢组学的影响,以期为低蛋白水平日粮和CA在猪生产中的合理应用提供理论依据,为缓解断奶仔猪应激的营养措施提供新的思路。
1 材料与方法
1.1 试验动物与设计
本试验选用体况相似、胎次相近的28日龄断奶的“杜洛克×长白×大白”三元杂交仔猪27头,按体重相近(9.87±0.14 kg)的原则,随机分为3个组,每组3个重复,每个重复3头猪。对照组饲喂正常蛋白水平日粮(NP,粗蛋白水平20%),试验组分别饲喂中等蛋白水平日粮+复合酸化剂(MP+CA,粗蛋白水平18%,复合酸化剂添加水平8 g/kg)和低蛋白水平日粮+复合酸化剂(LP+CA,粗蛋白水平16%,复合酸化剂添加水平8 g/kg)。预饲期3 d,正式期28 d。
1.2 试验日粮
以玉米-豆粕型日粮为基础,参照NRC(2012)11~25 kg阶段猪营养需要配制成粉状配合饲料。对照组日粮蛋白水平为20%,处理组日粮蛋白水平分别为18%和16%。为了准确配制不同蛋白水平日粮,并使日粮满足NRC(2012)标准回肠可消化氨基酸(SIDAA)水平,在处理组日粮中补充赖氨酸、蛋氨酸、苏氨酸和色氨酸4种必需氨基酸以满足仔猪断奶后的生长需要,同时补充大豆油使得3组之间消化能一致。试验日粮组成及营养水平见表1。复合酸化剂购自安徽五粮泰生物工程股份有限公司,主要组分为柠檬酸(≥35%)、乳酸(≥5%)、乙酸、丙酸和丁酸等。
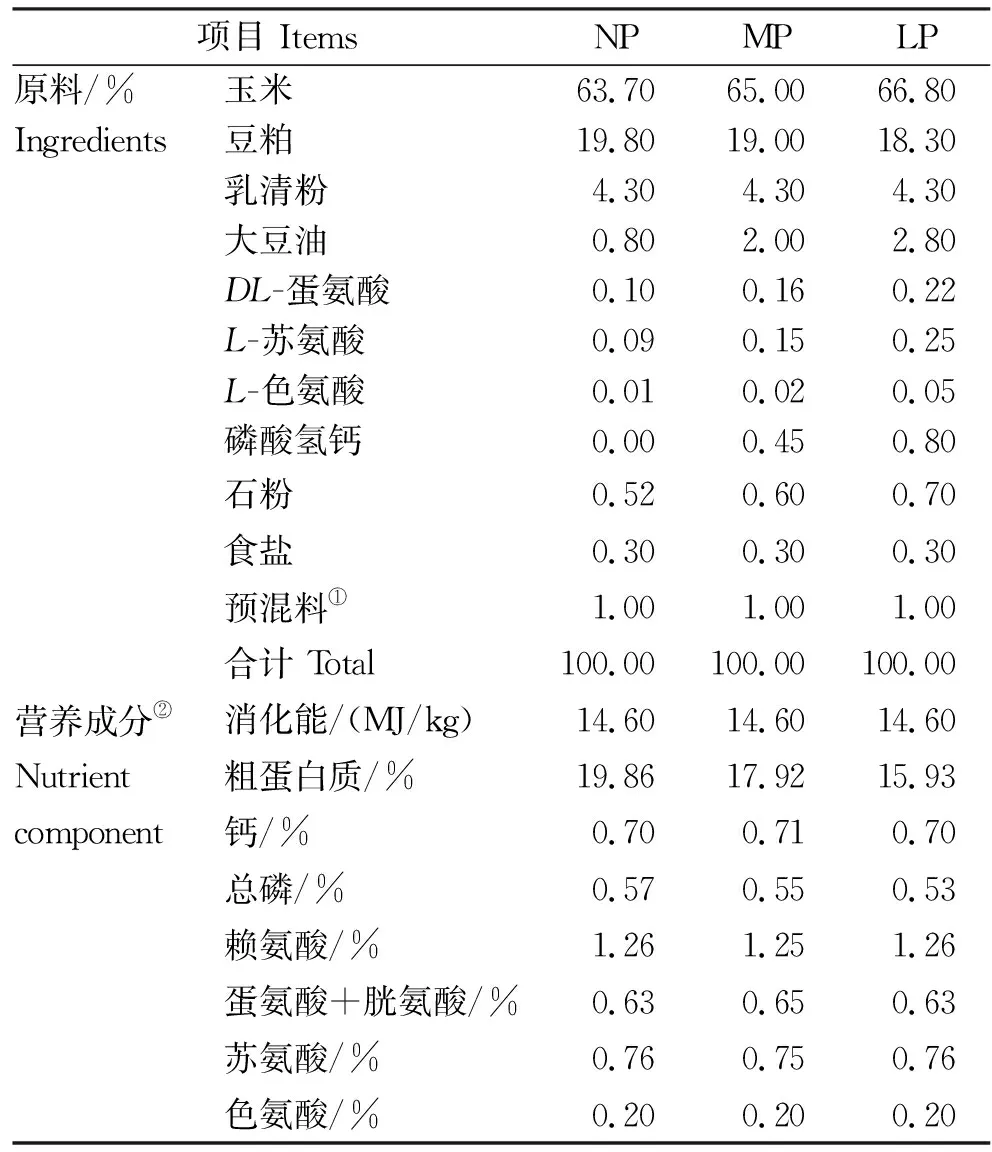
表1 试验日粮组成及营养水平Table 1 Ingredients and nutrient levels of the experimental diets
1.3 饲养管理
动物饲养试验于2019年8月在山西农业大学实验动物管理中心进行,采用全封闭式管理,所有试验猪均在同一猪舍内分栏饲养。猪舍地面为全漏缝塑料地板,每个饲养栏都配有不锈钢乳头式自动饮水器和硬质塑料料槽。在进猪前对猪舍进行消毒处理。保持猪舍卫生干净整洁。猪舍温度控制在25~27 ℃,相对湿度控制在70%左右。试验期间,猪舍消毒与仔猪驱虫、免疫均按猪场常规管理程序进行。
1.4 样品采集
于试验第28天清晨对仔猪进行称重、采血, 按照屠宰顺序提前2 h饲喂以确保仔猪后肠道充盈,便于食糜收集。从每个重复挑选1头接近该重复平均体重的仔猪进行屠宰,待胸腹腔打开后,结扎各个肠段,取相同部位盲肠、结肠食糜,装入灭菌的2 mL冻存管中,立即放入液氮中速冻并随后转入-80 ℃冰箱中贮存,用于肠道微生物测序和代谢物分析。
1.5 测定指标与方法
1.5.1 16S rRNA高通量测序
1.5.1.1 DNA提取与PCR扩增 取盲肠和结肠食糜样品各18份共36份,DNA提取和PCR扩增参照Zhang等方法。使用E.Z.N.A.D4015 Stool DNA Kit(Omega Bio-tek,美国)试剂盒提取样品总DNA。本试验选择细菌16S rRNA基因的V3~V4可变区为测序区域,引物序列为338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')。扩增体系如下: 5×FastPfu缓冲液4 μL,2.5 mmol/L dNTPs 2 μL,上、下游引物(5 μmol/L)各0.8 μL,FastPfu聚合酶(TransGen,中国)0.4 μL, DNA模板 10 ng,补充ddHO至20 μL。利用ABI GeneAmp9700(Applied Biosystem,美国)进行PCR扩增,扩增程序为:95 ℃预变性3 min,27个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),最后72 ℃延伸10 min,10 ℃保存。
1.5.1.2 Illumina Miseq测序 PCR产物经DNA凝胶回收试剂盒(Axygen,美国)回收,采用2%琼脂糖凝胶进行电泳检测。根据定量结果,将PCR产物用QuantiFluor-ST 蓝色荧光定量系统(Promega,美国)进行定量检测。样品在Miseq PE 300平台进行检测(上海美吉生物医药科技有限公司)。
1.5.1.3 数据处理与统计分析 使用FLASH(v.1.2.11)和Qiime(v.1.8.0)软件对测序序列进行拼接、筛选和过滤等处理,数据去杂方法和参数参照刘晶的方法。使用UCHIME(v.4.2.40)软件鉴定和去除PCR扩增产生的嵌合序列,获得最终的有效数据。利用Mothur(v.1.31.2)软件做Rarefaction分析,并基于样品OTU水平进行微生物Alpha多样性分析,计算Alpha多样性指数,并通过SPSS 22.0软件进行显著性检验。利用R软件进行基于Bray-curtis距离的PCoA主坐标分析。利用R软件中Vegan包的ANOSIM分析检验群落组间相似度,组间差异是否显著大于组内差异,从而判断分组是否有意义。根据SILVA细菌数据库,对OTU代表序列进行分类学分析,根据分析结果,计算门(Phylum)和属(Genus)两个分类学水平上样本的群落物种组成,用R语言工具绘制物种群落组成柱形图。利用线性判别分析LEfSe(Linear discriminant analysis Effect Size)寻找各组间在丰度上有显著差异的物种(LDA阈值大于3)。
1.5.2 液相色谱-质谱联用(LC-MS)分析
1.5.2.1 粪便样本处理 待测样品在冰上解冻后,称量样品50 mg转移至离心管中,加入500 μL -20 ℃预冷的70%的甲醇水(含l μg/mL的2-氯苯丙氨酸作为内标),涡旋3 min,然后在冰水浴中超声10 min,取出再涡旋1 min,静置后在4 ℃、12 000 r/min下离心10 min,吸取上清液到对应进样瓶中,用于LC-MS/MS分析。
1.5.2.2 色谱-质谱条件 采用Shim-pack (SHIMADZU)超高效液相色谱仪,使用液相色谱柱Waters ACQUITY UPLC HSS T3 C18(2.1 mm×100 mm,1.8 μm)对粪便样品进行色谱分离。液相色谱A相为超纯水(0.04%的乙酸),B相为乙腈(0.04%的乙酸);流速为0.4 mL/min,柱温为40 ℃,进样量为2 μL。样品质谱参数参照WEI等所提方法。
1.5.2.3 数据预处理 将原始数据导入Progenesis QI 软件中进行基线过滤、峰识别、积分、保留时间校正、峰对齐,最终得到一个保留时间、质荷比和峰强度的数据矩阵,然后参照曹青青的方法对数据进行预处理。
1.5.2.4 筛选差异代谢物 首先对进行差异比较的分组样品进行偏最小二乘分析(Partial Least Squares-Discriminant Analysis,PLS-DA),获得变量重要性投影值(Variable Importance in the Projection,VIP),然后结合Welch’s检验中的-value来进一步筛选出不同分组间的差异代谢物。筛选标准为VIP值>1且<0.05。
1.5.2.5 代谢通路分析 结合多元统计分析和单变量分析选出差异代谢物,应用MetaboAnalyst 5.0分析,对代谢物进行可视化图分析,通过京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路数据库,对差异代谢物所在通路进行富集分析,并做进一步的筛选,找到与代谢物差异关联性最高的关键代谢通路。
2 结果与分析
2.1 低蛋白水平日粮添加复合酸化剂对断奶仔猪后肠微生物组成的影响
2.1.1 微生物测序质量分析 利用Illumina Miseq平台对断奶仔猪盲肠和结肠36个样品所提取的菌群DNA进行16S rRNA基因V3~V4区测序,共得到1 987 885条有效序列数,序列平均长度为419.62 bp。检测样品的Sobs指数稀释曲线趋于平缓,不再随序列的增加而变化,说明测序深度已基本覆盖样本中的所有物种,测序质量良好。
2.1.2 Alpha多样性分析 由表2可知,在盲肠中,与NP组相比,MP+CA和LP+CA组Alpha多样性指数均无显著性差异(>0.05)。在结肠中,与NP组相比,MP+CA组Alpha多样性指数均无显著性差异(>0.05);LP+CA组Sobs指数、Ace指数和Chao指数均显著提高(<0.05)。结果表明,在低蛋白水平日粮中添加CA显著提高了结肠微生物群落的丰富度,对盲肠和结肠微生物群落的多样性没有显著影响。

表2 盲肠和结肠微生物的Alpha多样性指数Table 2 Alpha diversity index of cecum and colon
2.1.3 PCoA分析 PCoA 散点图(图1)显示NP组与MP+CA和LP+CA组盲肠和结肠样本分布距离较远,组间样本点没有交集,主坐标成分PC1和PC2分别占有36.11%、18.29%和34.44%、19.89%的样本组成差异,表明NP组与MP+CA和LP+CA组样本的微生物群落组成是可以相互区分的,通过相似性分析(ANOSIM)进一步证实了MP+CA和LP+CA组后肠微生物菌群结构发生极显著改变(<0.01)。

图 1 盲肠(A)和结肠(B)样品基于Bray-curtis的PCoA分析Fig. 1 PCoA analysis of cecal (A) and colonic (B) samples based on Bray-curtis distance
2.1.4 物种组成及差异分析 在属水平上,盲肠和结肠样本中相对丰度大于1%的细菌属分别于有25和34个,其共有物种中相对丰度较高的细菌属有乳酸杆菌属()、链球菌属()、罕见小球菌属()、严格梭菌属1(___1)、埃希氏-志贺菌属(-)等(图2A,图2B)。

图2 盲肠(A)和结肠(B)微生物在属水平上的物种组成Fig. 2 Species composition of cecal (A) and colonic (B) microbe at Genus level
基于LEfSe的多级物种差异判别分析表明,NP组与MP+CA组和LP+CA组盲肠和结肠样品在门、科和属等分类水平上均有相对丰度显著差异的细菌。如在属水平上,在盲肠中,与NP组相比,MP+CA组显著差异的细菌有互营球菌属()、普雷沃氏菌属_9(_9)和欧陆森氏菌属()等;LP+CA组显著差异的细菌有严格梭菌属1、毛螺菌科NK4A136群(Lachnospiraceae_NK4A136_group)和普雷沃氏菌科NK3B31群(Prevotellaceae_NK3B31_group)等(图3A)。在结肠中,与NP组相比,MP+CA组显著差异的细菌有、互营球菌属、和克里斯滕森菌科R7群(Christensenellaceae_R_7_group)等;LP+CA组显著差异的细菌有瘤胃菌科_UCG_014(Ruminococcaceae_UCG-014)、普雷沃氏菌科NK3B31群和克里斯滕森菌科R7群等(图3B)。
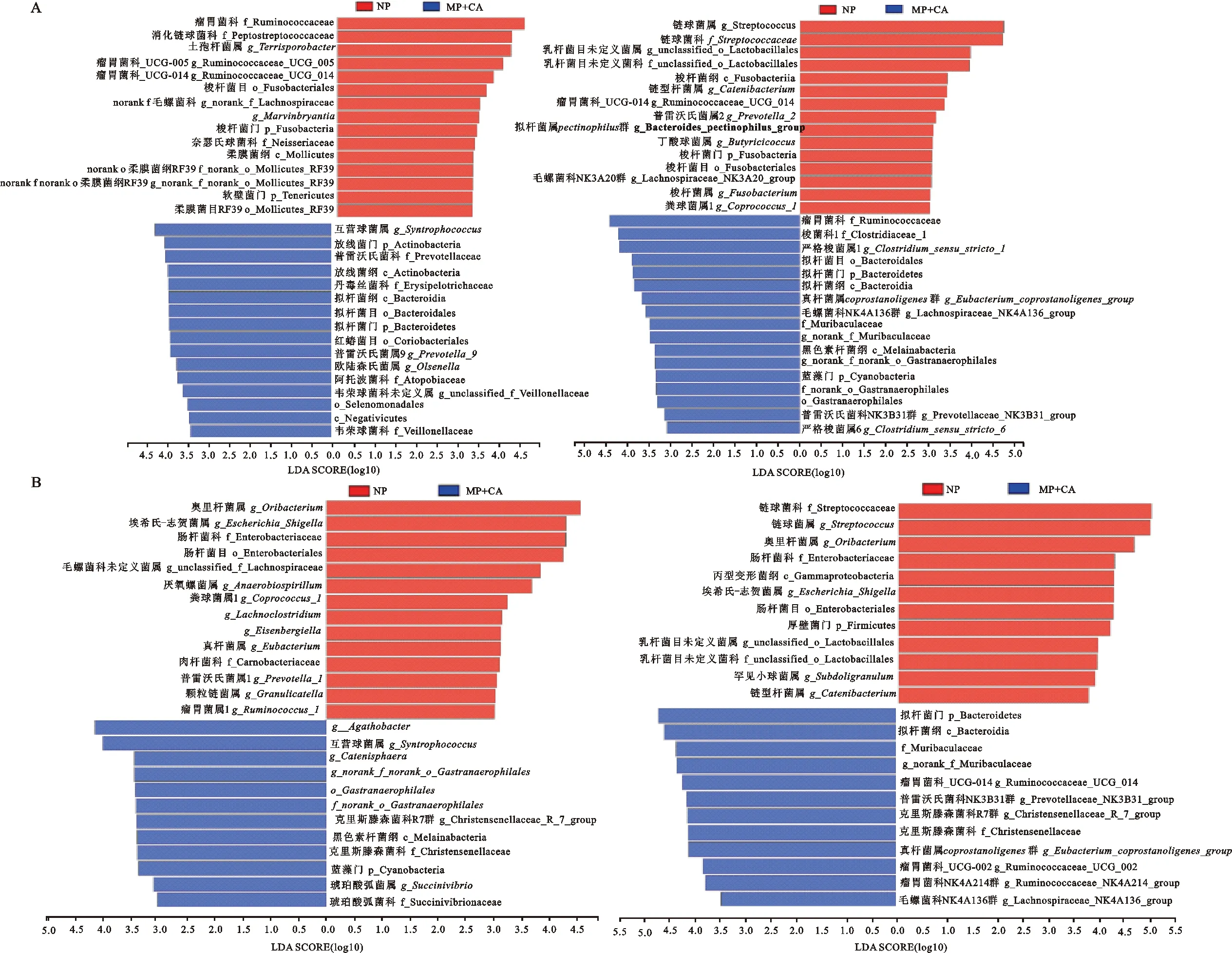
图3 盲肠(A)和结肠(B)微生物LEfSe差异分析LDA分值图Fig. 3 LDA score chart of cecal cecal (A) and colonic (B) microbe LEfSe difference analysis
2.2 低蛋白水平日粮添加复合酸化剂对断奶仔猪后肠微生物代谢组的影响
2.2.1 差异代谢物筛选 如图4所示,在盲肠中,NP组与MP+CA组和LP+CA组分别筛选出26和42种差异代谢物,其中共有的差异代谢物有7种,分别是三碘甲状腺原氨酸(3,3',5-Triiodo-L-Thyronine)、2-甲基丁酰肉毒碱(2-Methylbutyroylcarnitine)、-(+)-苹果酸(-(+)-Malic acid)、尿嘧啶核苷(Uridine)、核黄素(Riboflavin)、吲哚-3-甲醇(Indole-3-carbinol)和柠苹酸(Citramalic Acid)核苷(Uridine)和核黄素(Riboflavin)等。在结肠中,NP组与MP+CA组和LP+CA组分别筛选出47和69种差异代谢物,其中共有的差异代谢物有16种,包括丁香酸(Syringic Acid)、邻二甲苯(o-Xylene)、金合欢烯(Farnesene)、癸二酸(Sebacate)、2-乙酰基-5-甲基呋喃(2-Acetyl-5-methylfuran)、壬二酸(Azelaic Acid)、奎宁(Quinine)、三碘甲状腺原氨酸(3,3',5-Triiodo-L-Thyronine)、氧化芳樟醇(Linalyl oxide)、18-羟基皮质(甾)酮(18-Hydroxycorticosterone)、2'-脱氧胞苷-5'-单磷酸(2'-Deoxycytidine-5'-Monophosphate)、辛二酸(Subericacid)、5-羟基己酸(5-Hydroxyhexanoic Acid)、DL-3,4-二羟基苯基二醇(DL-3,4-Dihydroxyphenyl glycol)、对苯二酚(Hydroquinone)和葡萄糖醛酸(D-Glucoronic Acid)等。
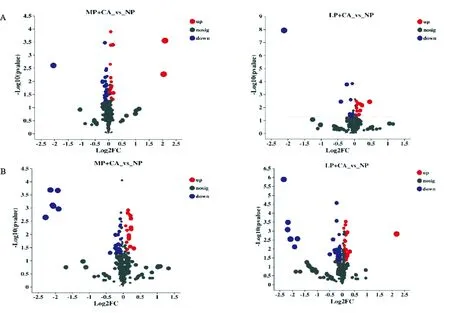
图4 盲肠(A)和结肠(B)差异代谢物火山图Fig. 4 The volcano map of the difference metabolites in the cecum (A) and colon (B)
2.2.2 代谢通路富集分析 分别对盲肠和结肠中筛选出的共有差异代谢物进行KEGG功能富集分析。结果表明,盲肠中差异代谢物主要富集在核黄素代谢(Riboflavin Metabolism)、嘧啶代谢(Pyrimidine Metabolism)和甲状腺激素合成(Thyroid hormone synthesis)等代谢通路上;而结肠中的差异代谢物则主要富集在甘油磷酸穿梭(Glycerol Phosphate Shuttle)、甲状腺激素合成(Thyroid hormone synthesis)和核黄素代谢(Riboflavin Metabolism)等代谢通路上(图5)。

图5 盲肠(A)和结肠(B)代谢通路富集分析图Fig. 5 Enrichment analysis diagram of metabolic pathways in the cecum (A) and colon (B)
3 讨 论
关于低蛋白水平日粮对肠道微生物区系丰富度和多样性报道并不一致,Peng等报道指出,适度降低日粮粗蛋白水平有利于肠道健康,增加了结肠细菌多样性,减少了有害蛋白质发酵产物的产生。Zhou等研究表明,降低日粮蛋白质水平3个百分点并不影响盲肠和结肠微生物的丰富度和多样性。在本研究中,在低蛋白水平日粮中添加复合酸化剂显著提高了结肠微生物群落的丰富度,对盲肠和结肠微生物群落的多样性没有显著影响。本研究相似性分析发现,NP组与MP+CA和LP+CA组微生物群落在PCoA散点图上被清晰地分隔开来,表明低蛋白水平日粮添加CA能够改善仔猪后肠微生物区系。
猪肠道中的细菌主要以厚壁菌门、拟杆菌门、变形菌门和放线菌门细菌为主,其中厚壁菌门和拟杆菌门约占整个菌群的90%,是肠道菌群中的优势菌种。在本试验中,盲肠中以厚壁菌门和变形菌门为优势菌群,而结肠中物种相对丰度最高的两个细菌门类分别是厚壁菌门和拟杆菌门。研究发现厚壁菌门是主要的纤维素分解菌,能够将纤维素降解为挥发性脂肪酸供宿主利用,而拟杆菌门细菌主要参与日粮中碳水化合物和蛋白质降解,在后肠微生物发酵及微生态平衡中发挥着关键作用。本研究中,在低蛋白水平日粮中添加CA降低了结肠中厚壁菌门的相对丰度,提高了拟杆菌门的相对丰度,同时厚壁菌门和拟杆菌门的比值下降,这可能是因为低蛋白水平日粮影响了宿主的代谢过程。厚壁菌门和拟杆菌门的比值可以反映宿主的脂质代谢情况,比值提高会引发与肥胖相关的代谢疾病,这与低蛋白水平日粮导致肥育猪背膘增厚、胴体无脂瘦肉率降低的现象一致。变形菌门的细菌大多为致病菌,包括大肠杆菌、志贺氏菌等,变形菌门细菌大多与宿主营养代谢紊乱或肠腔炎症有关。在本试验中,低蛋白水平日粮添加CA显著降低了盲肠和结肠中变形菌门的相对丰度,说明日粮蛋白水平降低和CA的添加抑制了致病菌的繁殖,原因可能是蛋白水平降低减少了这些细菌的发酵底物,同时添加酸化剂所带来的酸性环境对革兰氏阴性菌具有抑制作用。因此,在低蛋白水平日粮中添加复合酸化剂有利于促进动物肠道健康。
瘤胃菌科和普雷沃氏菌是猪肠道内主要的纤维降解菌,广泛分布在后肠中,可以分解纤维素产生短链脂肪酸。此外,普雷沃氏菌还参与了宿主碳水化合物和氨基酸的代谢,促进肝糖原储存。在本试验中,与NP组相比,在低蛋白水平日粮中添加CA显著提高了盲肠和结肠中严格梭菌属_1、毛螺菌科NK4A136群、克里斯滕森菌科R7群、瘤胃菌科_UCG-014、瘤胃菌科NK4A214群等属水平物种的相对丰度。其中,毛螺菌科广泛存在于哺乳动物肠道内,通常均被认为是有益菌,能够发酵纤维素产生SCFAs(如丁酸),SCFAs能够降低肠道pH,阻止有害微生物增殖,维持肠道微生物稳态,同时还可以抑制炎症性结肠炎。本研究中毛螺菌科比例提高,可能是由于低蛋白水平日粮营养成分中淀粉的含量明显增加,因此可供毛螺菌科代谢的底物增多。
核黄素(Riboflavin),又称维生素B,广泛参与生物体内生物氧化与能量代谢,与碳水化合物、蛋白质、核酸和脂肪的代谢有关,可提高机体对蛋白质的利用率,促进生长发育。研究表明猪在生长期核黄素缺乏会降低猪的日增重和抗氧化能力。在本试验中,与对照组相比,低蛋白水平日粮添加CA组后肠中均检测出差异代谢物核黄素,且都表现为显著上调,对差异代谢物进行的代谢通路分析也都富集在核黄素代谢通路上,这表明低蛋白水平日粮添加CA显著影响了核黄素的代谢,核黄素表达量上调进而影响了其它营养物质的代谢过程,促进了机体生长发育。
4 结 论
本试验结果表明,在低蛋白水平日粮中添加CA,显著提高了断奶仔猪后肠微生物群落的丰富度,对微生物群落的多样性没有显著影响;在门水平上,显著提高了后肠中厚壁菌门和拟杆菌门的相对丰度,降低了变形菌门的相对丰度;在属水平上,显著提高了后肠中互营球菌属、严格梭菌属1、毛螺菌科NK4A136群、克里斯滕森菌科R7群、瘤胃菌科_UCG-014、瘤胃球菌科NK4A214群等菌属的相对丰度,降低了链球菌属和埃希氏-志贺菌属等菌属的相对丰度。此外,在低蛋白水平日粮中添加CA显著影响了后肠中代谢物组成结构。差异代谢物分析表明,低蛋白水平日粮添加CA主要影响了核黄素代谢通路。
