西部煤矿区深色有隔内生真菌修复机理与生态应用模式
2022-02-25毕银丽彭苏萍王淑惠
毕银丽,彭苏萍,王淑惠
(1.西安科技大学 西部矿山生态环境修复研究院 陕西 西安 710054;2.中国矿业大学(北京) 煤炭资源与安全开采国家重点实验室 北京 100083)
西部煤矿区是我国矿产资源开发的主产区。该区域属干旱半干旱气候,年降雨量少,蒸发量大,土壤贫瘠,生物种群单一。煤炭开采是导致矿区土地退化的直接驱动力,不同煤炭开采工艺对生态环境的损伤特征存在差异,如井工开采致使矿区采煤沉陷地频发,沉陷裂缝发育损伤植物根系,降低土壤肥力,植被成活率降低,矿区生态环境更加脆弱,生态修复难度增加;露天开采剥离表土,土层搬运堆放致使排土场土壤无结构性,土壤养分低,扰动土壤生物种群锐减,生态重构效果差。为了解决西部矿区土壤生产力的恢复和生态群落重建问题,采用物理、化学和生物修复技术均可为矿区土地复垦提供有力的支持和保障,生物修复技术更具高效与可持续性。以微生物作为提升土壤功能的重要手段,可从根本上系统修复矿区土壤生态系统的功能,挖掘和激发土壤潜在肥力,对加快土壤改良、植被重建,促进矿区生态系统的稳定和可持续发展具有重要作用。
目前微生物复垦技术在煤矿区生态修复中的作用与应用受到越来越多专家学者关注,其中应用最多的微生物是丛枝菌根真菌(Arbuscular Mycorrhiza Fungi,AMF),该菌已在西部煤矿区取得了良好生态修复效应,技术应用趋于成熟。随着西部煤矿区微生物生态修复规模化应用,对菌剂的需求量更大。AMF无法纯培养,生产周期较长,对宿主植物有一定选择性,施用范围受植物种类一定限制。矿区生态修复对菌剂普适性与快速培养的需求使研究者开始关注另外一种微生物,深色有隔内生真菌(dark septate endophytes,DSE)。由于DSE具有广泛的宿主适应性,作为土壤微生物的重要组成部分,其发挥的生态学功能也引起了学者们广泛兴趣。
DSE泛指一类定殖于宿主植物根表皮、皮层,甚至维管束组织细胞内和细胞间隙,形成深色有隔菌丝和微菌核特征结构的内生真菌。研究表明,DSE具有与AMF相似的生态学功能,能促进宿主生长,提高宿主抗逆性。DSE可以纯培养,菌剂可以规模化生产,可为煤矿区大面积退化土地的修复快速提供充足微生物菌剂来源。西部煤矿区扰动土壤养分贫瘠,逆境胁迫(如高温、干旱、盐碱化、重金属污染、酷寒等)致使生态种群单一,生态系统稳定性差,是西部矿区土地复垦与生态修复的主要障碍因子。利用DSE在土壤和植物根系中的生态位优势,探究其在西部矿区生态修复的应用潜力。笔者主要从DSE的生态功能和矿区生态修复机理方面进行梳理,探索DSE菌剂在西部矿区生态修复的应用模式,为矿区生态修复提供新的技术和应用方向。
1 DSE生态特征
植物根际中存在大量DSE,DSE在空间和宿主植物中均未表现出特异性。有文献表明在我国西部陕西、内蒙古、新疆和宁夏煤矿区植物根际均发现有DSE定殖,阐明DSE生态特征可为其修复机理奠定基础。
1.1 DSE分类特征
在显微镜下观察植物根内DSE的形态学特征,深色有隔菌丝和微菌核是DSE的主要特征结构(图1)。单个菌丝沿根的主轴生长,可以在皮层细胞之间生长,也可以在表皮细胞之间的凹陷处生长。菌丝在皮层细胞间隙延伸并形成菌丝网,有时可延伸至细胞内形成胞内菌丝。菌丝颜色深浅不一,常见为深棕色至浅棕色,且随着菌丝的延伸,颜色可逐渐变浅。菌丝壁较厚,具明显的横隔。菌丝直或曲,常伴有树枝状随意隆起的分枝,并存在菌丝特化结构,如沿植物根细胞内壁盘曲的菌丝圈等。微菌核是由细胞壁膨大加厚的细胞紧密堆积形成,形状大小不一。颜色多为深棕色,偶见较浅的颜色,分布于表皮、皮层细胞中或细胞间隙,少见于维管组织内,有时可见其与菌丝相连。研究学者根据形态将微菌核定殖结构分为聚合型(脑状、哑铃状)和离散型(带状、链状、星点状等)。

图1 植物根内DSE定殖结构Fig.1 DSE colonization structure in roots
DSE可进行分离培养。培养基中的DSE菌落多呈圆形或椭圆形,颜色以灰色、黑色和褐色为主,表面平整或隆起具有褶皱,多有绒毛状覆盖,有些DSE产球形或椭圆形孢子,有些DSE不产孢。早期使用的长rRNA基因序列分析支持了DSE的多系统起源,DSE分类研究运用核糖体DNA-ITS序列分析技术进行基因序列检测。分子生物学分类可用于研究形态类群之间的系统发育关系,为更好地了解其与宿主的相互作用奠定了基础。通过对DSE真菌的分子结合形态系统研究和分类单元的重新评价,将为在形态相似、遗传序列不同的类群中分离出具有不同生态功能的DSE开辟道路。
1.2 不同宿主植物DSE定殖特点
DSE作为内生真菌,其与宿主植物间的互作关系以及定殖情况均会随着外界环境条件的不断变化而变化。植物物候是影响内生真菌在宿主植物中定殖的关键因素。研究学者发现最高水平的内生真菌定殖通常发生在宿主植物生长季节的高峰或后期,内生真菌数量的增减取决于植物的营养需求和植物向真菌提供的光合产物输出量,以及真菌对营养的需求和真菌向植物提供的矿物质营养的数量。这一相互作用反映了随宿主植物生长季节的变化,植物根系向内生真菌提供光合产物的输出量不同,2者之间的营养交换发生变化,进而影响内生真菌的定殖情况。DSE定殖与植物物候有关,但目前对于DSE与植物物候之间的关系仍不明确。MULLEN等发现,在生长季节开始时,毛茛属根内DSE定殖率非常高,促进了植物对氮的吸收。LI Han等研究发现夏季黑果枸杞根内DSE菌丝定殖率高,促进寄主植物对营养物质的吸收和利用;冬季微菌核定殖率高,有助于宿主植物抵御寒冷环境。然而,也有研究学者在高山苔原湿草甸植物群落中对4种多年生草本植物在萌芽期、盛花期、幼果期、果实成熟期和落叶期5个物候期进行采样研究,发现DSE菌丝定殖率随着时间的推移和宿主植物的不同均未发生显著变化。这可能由于不同种类宿主植物的物候期对DSE定殖的影响程度不同。
DSE的宿主范围较大,且缺乏宿主特异性,同种DSE可以定殖于多种宿主植物,例如,DSE ()能够定殖大于20个物种,但不同宿主植物根系中DSE定殖率存在差异。植物种类对DSE菌丝定殖率和总定殖率具有显著直接影响。在湿地生态系统中,DSE的定殖在单子叶植物中比双子叶植物中更普遍。此外,研究表明DSE在根系较老的部分能更频繁地定殖,表明DSE更喜欢衰老的根系组织。
1.3 不同生境中DSE生态型
在一个寄主物种中,微生物的生存环境相比宿主的基因型对其影响更大。研究表明从南非沿海平原和低地到热带、温带、亚高山、高山、海洋南极和北极地区的各种生境的宿主植物根系中均可观察到DSE的定殖结构,未表现出明显的区域或环境特异性。随海拔的升高,DSE菌丝变粗,颜色逐渐变深,横隔膜缩短;微菌核由分散颗粒状聚集成簇、成团,并出现迷宫状,颜色也显现不同变化规律,且菌丝定殖率、微菌核和总定殖率逐渐增大。
DSE分布和定殖具有明显的空间异质性,与土壤因子密切相关。一些研究者认为,土壤温度和土壤湿度在定殖水平中发挥作用。土壤有机质、全氮、全磷、速效钾、磷酸酶等土壤因子与DSE定殖率密切相关,但不同研究学者对于土壤因子与DSE定殖情况存在正/负相关性差异。土壤重金属含量也会影响DSE在宿主植物根内的定殖。DSE定殖情况与土层深度也存在相关关系,DSE总侵染率和定殖率随土层加深而减小。此外,有研究发现根际DSE菌丝定殖率高于根围。
可见,DSE在不同宿主植物和生境中存在不同的生态特征,但对于宿主植物种类和生境具有广泛适应性,且对环境的适应性较强,具有普适性。DSE可作为矿区生态修复的菌剂材料。
2 DSE在西部煤矿区生态修复作用机理
西部煤矿区气候干旱、土壤贫瘠、采煤扰动致使生物种群减少,生态系统脆弱。以微生物作为受损生态系统修复的切入点,可从根本上改善土壤的生物组成结构,提升土壤质量,恢复土地生态系统。矿区修复植被的健康与适应性不仅与植物基因组有关,还与环境中的多种生物因子有着错综复杂的联系。DSE对宿主植物的生态作用是多方面的,其促进宿主矿质营养的吸收,保护宿主免受非生物胁迫(水分胁迫、盐胁迫、重金属胁迫、寒冷胁迫等)和生物胁迫(病原菌伤害)都发挥重要作用,可增强生态系统稳定性。
2.1 DSE对宿主植物矿质营养吸收的作用机理
西部矿区土壤贫瘠,磷是关键的限制因子。而磷是植物生长发育所必需的大量营养元素之一,在植物体内不仅参与细胞的结构组成,而且在新陈代谢和遗传信息传递等方面都起着重要作用。HASELWANDTER和READ给接种DSE后,发现其能增加植物的生物量和茎秆中的磷含量。JUMPPONEN和TRAPPE研究发现,DSE可以分解不溶性磷酸盐供宿主植物利用。徐润冰将DSE菌片分别接种于有机磷和无机磷液体培养基中,发现DSE能分泌酸性磷酸酶和质子,将难溶性磷转换成可溶性磷酸根。通过对不同磷水平下接种DSE()对玉米磷吸收的影响发现DSE对不同形式的磷酸根吸收功能不同,DSE通过增强植物磷吸收相关基因的表达,促进植物分泌质子,增加细胞内可溶性磷酸根;还可以通过分泌大量酸性和碱性磷酸酶来分解难溶性磷,促进玉米磷元素吸收。

2.2 DSE对宿主植物抗逆性的作用机理
西部矿区生境差,植物的适应性弱而导致生态修复难。DSE对各种逆境,如干旱、低温、盐度、重金属等表现出很强的耐受力。植物可与内生真菌结合以提高其对环境胁迫的耐受性,提高生态修复效率。
..DSE对植物对水分胁迫的抗性作用
在干旱胁迫下,DSE对植物的定殖可以增加植物生物量,调节渗透物质,稳定光合作用参数,降低气孔导度,减少整体水分损失。LIU YAN等发现接种DSE()可有效降低干旱胁迫对细胞超微结构的损伤,帮助宿主改善根系的形态和构型,扩大根系的吸收范围,增加根系与土壤的接触,提高植物适应胁迫环境的能力。此外,接种DSE可以通过调节根系中激素含量水平和比例来调节根系形态和构型,促进根系生长,抵抗干旱胁迫环境,从而提高宿主的抗旱性。在干旱条件下,ZHANG Qiaoming等接种DSE(GM25)促进高粱(L. Moench)生长,加快植株光合速率和蒸腾速率,提高次生代谢产物花青素含量,改善次生代谢相关的酶活性。
..DSE对植物盐胁迫的抗性机理
邓勋等在PDA培养基中培养DSE(),发现DSE对NaCl和NaSO耐受性较好。之后,进一步将DSE接种到樟子松苗木中,通过提高宿主植物的抗氧化酶活性,同时降低丙二醛(MDA)和游离脯氨酸含量来缓解盐胁迫对樟子松苗木的伤害。MATEU等研究发现芦苇根内的DSE()增强了芦苇的耐盐性,提高了在易受海水入侵地区的存活率。此外,DSE自身产生的黑色素、甘露醇和海藻糖等代谢产物可以帮助宿主提高对盐胁迫的抗性。ZHAN Fangdong等发现黑色素可以消除盐胁迫产生的自由基,DSE中分离出的黑色素被报道具有很高的抗氧化活性。甘露醇和海藻糖等被认为是保护植物免受非生物胁迫引起的细胞损伤的重要渗透物质,DSE细胞可以吸收来自宿主的蔗糖,最终将其转化为甘露醇和海藻糖。值得思考的是,干旱和盐胁迫诱发了类似的整体胁迫(水分亏缺状态),触发了类似的植物早期反应;因此,帮助植物抵御盐度的微生物也可能使植物耐受干旱胁迫。
..DSE对植物重金属胁迫的抗性机理
DSE可以将金属离子或其络合物直接泵出宿主植物细胞外,通过细胞外螯合防止重金属元素渗透到细胞内,并在细胞间隔中络合和隔离重金属元素以抑制其毒性。例如,DSE可以通过转运以PCs络合物或谷胱甘肽络合物形式存在的重金属至宿主植物细胞液泡或其他位于细胞质的泡囊中区隔储存,将金属离子FeS簇结合物从宿主植物线粒体膜转运至细胞质中,并区隔储存。
在蛋白和转录水平上,DSE可以通过参与调控宿主植物根系细胞壁合成通路相关基因的表达,激活细胞壁合成关键酶的活性,强化细胞壁对根系吸收的游离重金属离子的固持与区室化作用,进而改变重金属离子在植物根组织、细胞和亚细胞水平上的分布,限制其向原生质体和地上部分的运输,以减轻重金属毒害。此外,利用微阵列或RNA-seq技术进行了DSE和转录组研究,以确定其涉及金属耐受性的基因。这2项研究报告了中谷胱甘肽s -转移酶和Nramp金属转运蛋白的功能特征,这是DSE中惟一可用的功能基因组数据。
..DSE对植物抗寒的作用机理
低温可引起生物分子间的物理和化学变化,低温(<15 ℃)和冰冻(<0 ℃)条件对植物的发育有重要的限制。在极地和高山生态系统中,DSE可能对植物养分获取有积极影响,从而提高植物对寒冷条件的耐受力。UPSON 等研究发现,与对照组相比,在5 ℃条件下,接种6种DSE的可以显著提高宿主植物的地上和地下生物量。同样,养分有效性也改变了DSE()和大麦()之间相互作用的结果,在8 ℃条件下,无论土壤养分有效性高或低,植株生物量和粒重均得到提高。
..DSE对植物抗病的作用机理
微生物可以通过增强植物的抵抗力或耐受性,或通过它们对植物病原体的直接拮抗来保护植物免受病原体的伤害。一些研究表明,DSE具有控制植物病害的潜力。DSE()通过积累HO和诱导植物系统抗性保护水稻植株不受稻瘟病病原菌()侵染,DSE()降低了离体卷心菜中黄萎病病原菌()的症状,DSE()降低了感染病原菌()的番茄植株的萎蔫病,DSE(sp.)在提供有机氮源的培养基中增强了甜瓜幼苗对病原菌(f. sp.)的抗性。可见,DSE可以通过改变植物的生物化学特性进而提高植物的抗病性或耐受性。DSE可以通过增强植物抗氧化系统,调节抗病相关酶活性和抗病相关物质的含量,提高植物抗病性。此外,还可通过改善植物根际微环境,增加微生物群落稳定性,从而提高植物的存活能力,增强植物抗病性。
2.3 DSE代谢物对宿主植物的刺激效应
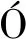
3 DSE在西部煤矿区生态修复应用模式
微生物菌剂在矿区生态环境修复领域的应用被认为是可持续技术,具有经济高效、低能耗、不产生二次污染等特点,矿区微生物修复具有较大的应用潜力和市场。目前关于DSE的研究大多限于DSE生态多样性调查和室内抗逆机理,对DSE菌剂的开发及生态修复应用仍处于起步阶段。利用DSE特性开发出液体应用方法和固体菌剂产品,规模化应用于矿区生态修复将是微生物复垦新的思路和方向。
3.1 DSE菌液浸种搭载无人机飞播生态修复模式
常规液体菌剂野外规模化应用是将液体发酵菌液直接施用于根际土壤,菌剂的需求量大,且液体储存、运输成本高,大面积矿区复垦实践很难推广。考虑矿区生态修复普遍采用大面积撒播灌草种子,且地形复杂多变,采用低成本高效的菌液浸种搭载无人机飞播技术可实现多物种组合的立体快速生态修复效果(图2)。
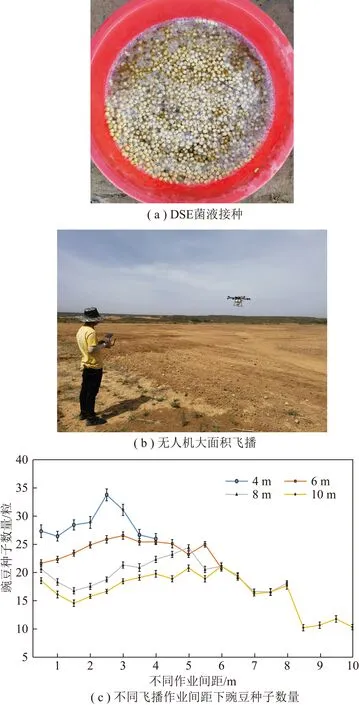
图2 无人机飞播作业及飞播参数试验Fig.2 Uar broadcast operation and broadcast parameters test
微生物菌液浸种可以促进植物健康生长,提高植物抗逆性。采用浓度为20%的DSE(sp.)针A2-7的菌液浸泡玉米种子12 h可以促进种子快速萌发,并显著促进玉米幼苗生长,对于在西部煤矿区植被重建具有重大的应用推广价值。
无人机飞播具有作业速度快、工作效率高、适用范围广、操作便利等优点。在西部采煤沉陷区,可以通过对采煤沉陷区地形图的坡度分级和地面覆盖率,确定无人机飞播过程中的撒播系数和飞行参数,如在陕北张家峁矿区飞播豌豆的无人机飞行高度为2 m、飞行速度为4 m/s、材料漏速为8.22 kg/min,作业间距为6 m。通过无人机飞播遥控式作业,不仅可以提高种子撒播均匀度、减少劳动力成本,还可以使地形复杂的西部矿区实现“全覆盖”生态修复。
3.2 DSE固体菌剂生态修复模式
生长活跃的DSE菌丝体相比微菌核在土壤中生长更快、更活跃。DSE菌液可以浓缩菌丝体干燥后制成DSE固体菌剂胶囊,也可采用矿区本地采矿伴生黏土制丸粒,具有广阔的应用前景(图3)。胶囊和丸粒固体菌剂可常温保存,解决液体菌剂不宜长期储存且运输不便等问题。DSE(sp.)针A2-1在60 ℃下干燥,常温保存20 d仍可保持DSE生物活性,固体菌剂研发可延长DSE菌存活时间,更大程度发挥其促进矿区生态恢复的价值。菌剂接种到根际土壤后繁殖迅速,侵染率高。培养10 d的DSE固体菌剂对紫花苜蓿的促生作用显著,能够提高紫花苜蓿的养分吸收能力。
图3 固体菌剂研制Fig.3 Development of solid bactericide
3.3 DSE菌液叶面肥涂抹生态修复模式
DSE菌液进入土壤后容易被土壤吸附或者固定,影响菌肥效应的发挥,所需菌液量较大,费工费力。将叶面肥DSE菌液以叶片涂抹方式接入,可以最大效率提高菌肥的利用率,产生较好的接菌生态效应,可以提高叶片对菌液有效养分的吸收。DSE菌液代谢物是影响宿主植物和土壤环境的重要化合物,影响植物在分子水平上的反应,促进色素、次生代谢物、激素、抗氧化剂和生物碱的生物合成,这对宿主植物促生等也发挥着重要作用。
盆栽实验发现,在叶面直接涂抹DSE菌液可以明显促进蛋白桑生长,提高蛋白桑营养品质(图4),对蛋白桑生物量的贡献率高达226%。采用液相色谱质谱联用分析(UPLC-MS)检测DSE菌液代谢产物,含有137余种化合物,包括氨基酸和肽类、有机氧化合物、脂质和类脂质分子、有机杂环化合物、苯丙烷和聚酮化合物等11种类型,其中以氨基酸和肽类为主,占总化合物的36.45%。
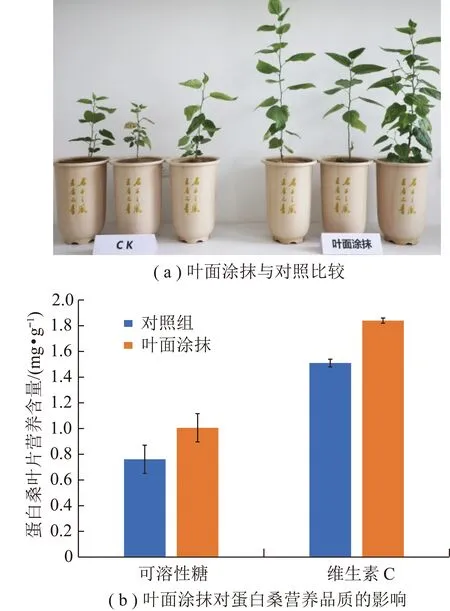
图4 DSE代谢物叶面涂抹效果Fig.4 Foliar coating effect of DSE metabolites
氨基酸和肽类化合物作为叶面肥可以促进植物生长,提高抗冻、抗旱、耐盐、光合活性和对真菌、细菌和病毒的抵抗力。相比直接施入土壤方式,叶面肥涂抹更为高效。后期需要考虑环境参数的影响(如日照、降水),需对植物吸收、运输和利用叶面肥养分的物理、化学、生物和环境等机理进一步探索,进一步提升DSE菌液作为叶面肥的功效。
综上所述,DSE菌剂产品可对西部煤矿区生态修复产生积极促进作用,有潜在的生态修复利用价值,但DSE生态修复的长期生态效应仍有许多未知需要探索,各种非生物因素(植物种类、土壤pH值、水分、质地、养分和盐度等)均可能会影响DSE菌剂的定殖和生态效应,DSE对西部矿区生态修复长期作用效果仍有待进一步深入研究。
4 结 语
DSE作为自然界土壤中的一类广谱真菌,对于促进土壤养分活化,增加植物生物量,抵抗逆境具有重要的作用,可以减缓煤炭开采对土壤性状和生物种群的损伤,对于煤矿区受损生态修复具有积极促进意义。DSE浸种搭载无人机飞播、固体菌剂研发、DSE叶面肥的开发等多样化生态修复模式将会为西部煤矿区土地复垦与生态修复提供多样化的技术支撑,具有重要的现实生态意义。预计在未来,多样化的DSE菌剂材料将应用矿区生态修复,以加速实现西部矿区的生态、社会和经济效益。
[1] 彭苏萍,毕银丽. 黄河流域煤矿区生态环境修复关键技术与战略思考 [J]. 煤炭学报,2020,45(4):1211-1221.
PENG Suping,BI Yinli. Key technology and strategic thinking of ecological environment restoration in coal mine area of Yellow River Basin [J]. Journal of China Coal Society,2020,45(4):1211-1221.
[2] 毕银丽,彭苏萍,杜善周. 西部干旱半干旱露天煤矿生态重构技术难点及发展方向[J]. 煤炭学报,2021,46(5):1355-1364.
BI Yinli,PENG Suping,DU Shanzhou. Technical difficulties and development direction of ecological reconstruction in arid and semi-arid open-pit coal mines in western China[J]. Journal of China Coal Society,2021,46(5):1355-1364.
[3] FENG Y,WANG J M,BAI Z K,et al. Effects of surface coal mining and land reclamation on soil properties:A review[J]. Earth-Science Reviews,2019,191:12-25.
[4] 毕银丽. 丛枝菌根真菌在煤矿区沉陷地生态修复应用研究进展[J]. 菌物学报,2017,36(7):800-806.
BI Yinli. Research progress of arbuscular mycorrhizal fungi in ecological restoration of coal mine subsidence[J]. Mycosystema,2017,36(7):800-806.
[5] 毕银丽,解琳琳. 丛枝菌根真菌与深色有隔内生真菌生态修复功能与作用[J]. 微生物学报,2021,61(1):58-67.
BI Yinli,XIE Linlin. Ecological restoration of arbuscular mycorrhizal fungi and dark septum endophytic fungi[J]. Acta microbiologica Sinica,2021,61(1):58-67.
[6] 罗园园,郝鲜俊,张铠珏. 接种AM真菌对煤矿区废弃土壤不同磷素形态的影响[J]. 西南农业学报,2019,32(2):381-388.
LUO Yuanyuan,HAO Xianjun,ZHANG Kaijue. Effects of inoculated AM fungi on different phosphorus forms in waste soil of coal mine area[J]. Southwest China Journal of Agricultural Sciences,2019,32(2):381-388.
[7] RUOTSALAINEN A L,KAUPPINEN M,WALI P R,et al. Dark septate endophytes:Mutualism from by-products? [J]. Trends in Plant Science,2021:ISSN 1360-1385,https://doi.org/10.1016/j.tpants.2021.10.001.
[8] JUMPPONEN A,TRAPPE J M. Dark septate endophytes:A review of facultative biotrophic root-colonizing fungi[J]. New Phytologist,1998,140(2):295-310.
[9] 刘茂军,张兴涛,赵之伟. 深色有隔内生真菌(DSE)研究进展[J]. 菌物学报,2009,28(6):888-894.
LIU Maojun,ZHANG Xingtao,ZHAO Zhiwei. Advances in the research of dark septate endophytes[J]. Mycosystema,2009,28(6):888-894.
[10] 毕银丽,任颖,全文智. 一种深色有隔内生真菌菌液高效规模化产运联动生产方法[P]. 中国专利:CN111394256B. 2021-10-15.
[11] CLEMMENSEN K E,FINLAY R D,DAHLBERG A,et al. Carbon sequestration is related to mycorrhizal fungal community shifts
during long-term succession in boreal forests[J]. New Phytologist,2015,205(4):1525-1536.
[12] ZUO Y L,SU F,HE X L,et al. Colonization by dark septate endophytes improves the growth ofmultiple inoculum levels[J]. Symbiosis,2020,82(3):201-214.
[13] 徐风美,王春燕,褚洪龙,等. 松萎蔫病发生区和未发生区油松根部真菌群落研究[J]. 西北植物学报,2014,34(8):1627-1634.
XU Fengmei,WANG Chunyan,CHU Honglong,et al. Study on fungal community ofroot in occurrence and non-occurrence areas of pine wilt disease[J]. Acta Botanica Boreali-Occidentalia Sinica,2014,34(8):1627-1634.
[14] O’DELL T E,MASSICOTTE H B,TRAPPE J M. Root colonization ofAgardh. andDougl. byWang & Wilcox[J].New Phytologist,1993,124(1):93-100.
[15] 王少杰,侯力峰,解琳琳,等. 蒙古沙冬青根系DSE定殖与土壤因子典范对应分析[J]. 河北大学学报(自然科学版),2017,37(5):497-507.
WANG Shaojie,HOU Lifeng,XIE Linlin,et al. Model correspondence analysis of root DSE colonization and soil factors of Ilex mongolica[J]. Journal of Hebei University (Natural Science Edition),2017,37(5):497-507.
[16] 胡丽杰,闫思远,孙牧笛,等. 宁夏枸杞深色有隔内生真菌的侵染特征及多样性[J]. 微生物学通报,2019,46(11):2973-2984.
HU Lijie,YAN Siyuan,SUN Mudi,et al. Characterization and diversity of endophytic fungi from Lycium barbarum in Ningxia[J]. Microbiology China,2019,46(11):2973-2984.
[17] GRUNIG C R,SIEBER T N,HOLDENRIEDER O. Characterisation of dark septate endophytic fungi (DSE) using inter-simple-sequence-repeat-anchored polymerase chain reaction (ISSR-PCR) amplification[J]. Mycological Research,2001,105:24-32.
[18] DE MESQUITA CPB,DEL RIO CMM,SUDING KN,et al. Rapid temporal changes in root colonization by arbuscular mycorrhizal fungi and fine root endophytes,not dark septate endophytes,track plant activity and environment in an alpine ecosystem[J]. Mycorrhiza,2018,28(8):717-726.
[19] MANDYAM K,JUMPPONEN A. Seasonal and temporal dynamics of arbuscular mycorrhizal and dark septate endophytic fungi in a tallgrass prairie ecosystem are minimally affected by nitrogen enrichment[J]. Mycorrhiza,2008,18(3):145-155.
[20] KIERS E T,DUHAMEL M,BEESETTY Y,et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis[J]. Science,2011,333:880-882.
[21] HAN L,SHI J X,HE C,et al. Temporal and spatial dynamics of dark septate endophytes in the roots ofin the desert region of Northwest China[J]. Agronomy-Basel,2021,11(4):648.
[22] MULLEN R B,SCHMIDT S K,III C H J. Nitrogen uptake during snowmelt by the snow buttercup,ranunculus adoneus[J]. Arctic and Alpine Research,1998,30(2):121.
[23] HAN L,ZUO Y L,HE X L,et al. Plant identity and soil variables shift the colonisation and species composition of dark septate endophytes associated with medicinal plants in a northern farmland in China[J]. Applied Soil Ecology,2021,167:104042.
[24] RODRIGUEZ R J,WHITE J Fjr.,ARNOLD A E,et al. Fungal endophytes:Diversity and functional roles[J]. New Phytologist,2009,182(2):314-330.
[25] WEISHAMPEL P A,BEDFORD B L. Wetland dicots and monocots differ in colonization by arbuscular mycorrhizal fungi and dark septate endophytes[J]. Mycorrhiza,2006,16(7):495-502.
[26] PEIFFER J A,SPOR A,KOREN O,et al. Diversity and heritability of the maize rhizosphere microbiome under field conditions[J].Proceeding of the National Academy of Science of the United States of America,2013,110(16):6548-6553.
[27] 李宝库,贺学礼,吴伟家. 不同海拔蒙古沙冬青根系深色有隔内生真菌(DSE)定殖的空间变化[J]. 江苏农业科学,2016,44(5):502-505.
LI Baoku,HE Xueli,WU Weijia. Spatial variation of colonization of dark septate endophytic fungi (DSE) in roots of Ilex mongolica at different elevations[J]. Jiangsu Agricultural Sciences,2016,44(5):502-505.
[28] 葛佳丽,张琳琳,李欣玫,等. 极旱荒漠植物深色有隔内生真菌定殖及其与土壤因子的相关性[J]. 河北大学学报(自然科学版),2018,38(1):73-82.
GE Jiali,ZHANG Linlin,LI Xinmei,et al. Colonization of dark septal endophytic fungi and its correlation with soil factors in desert plants of extreme drought [J]. Journal of Hebei University (Natural Science Edition),2018,38(1):73-82.
[29] 孙茜,贺超,贺学礼,等. 沙冬青与伴生植物深色有隔内生真菌定殖规律及其与土壤因子的相关性[J]. 植物生态学报,2015,39(9):878-889.
SUN Qian,HE Chao,HE Xueli,et al. Colonization patterns of endophytic fungi with dark septum and its correlation with soil factors[J]. Chinese Journal of Plant Ecology,2015,39(9):878-889.
[30] 刘欢. 紫茎泽兰DSE多样性及其重金属耐性初探[D]. 昆明:云南大学,2017:12-19.
LIU Huan. Preliminary study on DSE diversity and heavy metal tolerance of[D]. Kunming:Yunnan University,2017:12-19.
[31] 陈严严,贺学礼,李宝库,等. 蒙古沙冬青根系深色有隔内生真菌共生及分离培养特征研究[J]. 河北农业大学学报,2014,37(5):18-24.
CHEN Yanyan,HE Xueli,LI Baoku,et al. Study on symbiotic and isolated culture characteristics of endophytic fungi with dark septum in root system of Ilex mongolica[J]. Journal of Agricultural University of Hebei,2014,37(5):18-24.
[32] VANNIER N,MONY C,BITTEBIERE A K,et al. Epigenetic me-
chanisms and microbiota as a toolbox for plant phenotypic adjustment to environment[J]. Frontiers in Plant Science,2015,6:1159-1167.
[33] HASELWANDTER K,READ D J. The significance of a root-fun-
gus association in twospecies of high-alpine plant communities[J]. Oecologia,1982,53(3):352-354.
[34] 徐润冰. 深色有隔内生真菌嗜鱼外瓶霉()促进玉米磷吸收的机制研究[D]. 昆明:云南大学,2017:14-31.
XU Runbing. Study on the mechanism of the dark septum endophytic funguspromoting phosphorus uptake in maize[D]. Kunming:Yunnan University,2017:14-31.
[35] WU F L,QU D H,WEI T,et al. Transcriptome analysis for understanding the mechanism of dark septate endophyte S16 in promoting the growth and nitrate uptake of sweet cherry [J]. Journal of Integrative Agriculture,2021,20(7):1819-1831.
[36] LI X,HE X,HOU L,et al. Dark septate endophytes isolated from a xerophyte plant promote the growth of ammopiptanthus mongolicus under drought condition[J]. Scientific Reports,2018,8(1):7896-7907.
[37] MENOYO E,TESTE F P,FERRERO M A,et al. Associations between fungal root endophytes and grass dominance in arid highlands[J]. Fungal Ecology,2020,45:1-8.
[38] CHEN E,BLAZE J A,SMITH R S,et al. Freeze tolerance of poleward-spreading mangrove species weakened by soil properties of resident salt marsh competitor[J]. Journal of Ecology,2020,108(4):1725-1737.
[39] LIU Y,WEI X. Dark Septate endophyte improves drought tolerance ofHemsley & E. H. Wilson by modulating root morphology,ultrastructure,and the ratio of root hormones[J]. Forests,2019,10(10):1-13.
[40] ZHANG Q,GONG M,YUAN J,et al. Dark septate endophyte improves drought tolerance in sorghum[J]. International Journal of Agriculture and Biology,2017,19(1):53-60.
[41] 邓勋,宋小双,尹大川,等. 盐胁迫对2株深色有隔内生真菌(DSE)生长及营养代谢的影响[J]. 中南林业科技大学学报,2015,35(5):1-8.
DENG Xun,SONG Xiaoshuang,YIN Dachuan,et al. Effects of salt stress on growth and nutrient metabolism of two endophytic fungi (DSE) [J]. Journal of Central South University of Forestry & Technology,2015,35(5):1-8.
[42] MATEU M G,BALDWIN A H,MAUL J E,et al. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis[J]. ISME Journal,2020,14(8):1943-1954.
[43] JACOBSON E S,TINNELL S B. Antioxidant function of fungal melanin[J]. Journal of Bacteriology,1993,175(21):7102-7104.
[44] ZHAN F,HE Y,ZU Y,et al. Characterization of melanin isolated from a dark septate endophyte (DSE),[J]. World Journal of Microbiology and Biotechnology,2011,27(10):2483-2489.
[45] USUKI F,NARISAWA H. A mutualistic symbiosis between a dark septate endophytic fungus,,and a nonmycorrhizal plant,Chinese cabbage[J]. Mycologia,2007,99(2):175-184.
[46] 曹冠华. 嗜鱼外瓶霉()ABC转运蛋白基因与宿主重金属耐性的研究[D]. 昆明:云南大学,2017:118-120.
CAO Guanhua. Study on ABC transporter gene ofand heavy metal tolerance of host [D]. Kunming:Yunnan University,2017:118-120.
[47] GORFER M,PERSAK H,BERGER H,et al. Identification of heavy
metal regulated genes from the root associated ascomycete Cadophora finlandica using a genomic microarray[J]. Mycological Research,2009,113:1377-1388.
[48] ZHAO D,LI T,SHEN M,et al. Diverse strategies conferring extreme cadmium (Cd) tolerance in the dark septate endophyte (DSE),:Evidence from RNA-seq data[J]. Microbiological Research,2015,170:27-35.
[49] HILL P W,BROUGHTON R,BOUGOURE J,et al. Angiosperm symbioses with non-mycorrhizal fungal partners enhance N acquisition from ancient organic matter in a warming maritime Antarctic[J]. Ecology Letters,2019,22(12):2111-2119.
[50] UPSON R,READ D J,NEWSHAM K K. Nitrogen form influences the response of deschampsia antarctica to dark septate root endophytes[J]. Mycorrhiza,2009,20(1):1-11.
[51] SONG X H,LI Y K,HU Y,et al. Endophytes from blueberry roots and their antifungal activity and plant growth enhancement effects [J]. Rhizosphere,2021,20:1-8.
[52] SU Z Z,MAO L J,LI N,et al. Evidence for biotrophic lifestyle and biocontrol potential of dark septate endophyte harpophora oryzae to rice blast disease[J]. Plos One,2013,8(4):e61332.
[53] NARISAWA K,USUKI F,HASHIBA T. Control of verticillium yell-
ows in Chinese cabbage by the dark septate endophytic fungus LtVB3[J]. Phytopathology,2004,94(5):412-418.
[54] ANDRADE-LINARES D R,GROSCH R,RESTREPO S,et al. Effects
of dark septate endophytes on tomato plant performance[J]. Mycorrhiza,2011,21(5):413-422.
[55] KHASTINI R O,OGAWARA T,SATO Y,et al. Control ofwilt in melon by the fungal endophyte,sp. [J]. European Journal of Plant Pathology,2014,139(2):333-342.
[56] YAKTI W,KOVACS G M,FRANKEN P. Differential interaction of the dark septate endophytesp. and fungal pathogens in vitro and in planta[J]. FEMS Microbiology Ecology,2019,95(12):1-12.
[57] RINSCHEN M M,IVANISEVIC J,GIERA M,et al. Identification of bioactive metabolites using activity metabolomics[J]. Nature Reviews Molecular Cell Biology,2019,20(6):353-367.
[58] TIENAHO J,KARONEN M,MUILU-MAKELA R,et al. Metabolic profiling of water-soluble compounds from the extracts of dark septate endophytic fungi (DSE) isolated from Scots Pine (L. ) seedlings using UPLC-Orbitrap-MS[J]. Molecules,2019,24(12):2330-2338.
[59] PELLEGRINI M,PAGNANI G,BERNARDI M,et al. Cell-free supernatants of plant growth-promoting bacteria:A review of their use as biostimulant and microbial biocontrol agents in sustainable agriculture[J]. Sustainability,2020,12(23):1-22.
[60] LI X Y,WANG Y H,YANG J,et al. Acaricidal activity of cyclodipeptides fromW1 against[J]. Journal of Agricultural and Food Chemistry,2018,66(39):10163-10168.
[61] SILESS G E,GALLARDO G L,RODRIGUEZ M A,et al. Metabolites from the dark septate endophytesp. Evaluation by LC/MS and principal component analysis of culture extracts with histone deacetylase inhibitors[J]. Chemistry & Biodiversity,2018,15(8):e1800133.
[62] 沙月霞. 嗜碱假单胞菌Ej2浸种对盐胁迫下水稻根系内生菌群落结构的影响[J/OL].微生物学报,https://doi. org/10. 13343/j. cnki. wsxb. 20210656.
SHA Yuexia. Effects ofEj2 seed soaking on endophytic community of rice roots under salt stress. [J/OL]. Acta Microbiologica Sinica,https://doi. org/10. 13343/j. cnki. wsxb. 20210656.
[63] 毕银丽,薛子可,全文智. 一种利用深色有隔内生真菌浸种促进植物幼苗生长发育的方法[P]. 中国专利:CN111373899B. 2020-12-22.
[64] 毕银丽,马少鹏,高雅坤,等. 一种采煤沉陷区飞播生物修复方法[P]. 中国专利:CN112956310A. 2021-06-15.
[65] JOHN R P,TYAGI R D,BRAR S K,et al. Bio-encapsulation of microbial cells for targeted agricultural delivery[J]. Critical Reviews in Biotechnology,2011,31(3):211-226.
[66] 毕银丽,任颖,全文智. 一种深色有隔内生真菌干粉菌剂的制备方法[P]. 中国专利:CN111411047A. 2020-07-14.
[67] 王淑惠,毕银丽,李梦琪. 不同培养期深色有隔内生真菌链格孢菌对紫花苜蓿生长的影响[J]. 菌物学报,2021,40(10):2863-2873.
WANG Shuhui,BI Yinli,LI Mengqi. Effects of dark septate endophytesp. with different culture periods on growth of[J]. Mycosystema,2021,40(10):2863-2873.
[68] COLLA G,HOAGLAND L,RUZZI M,et al. Biostimulant action of protein hydrolysates:Unraveling their effects on plant physiology and microbiome[J]. Frontiers in Plant Science,2017,8:1-14.
[69] SHARMA H S S,FLEMING C,SELBY C,et al. Plant biostimul-ants:A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses[J]. Journal of Applied Phycology,2014,26(1):465-490.
