Morphological and Phylogenetic Studies of a Copepod Species, Irodes parupenei Ho and Lin (2007), Infecting Parupeneus rubescens in Saudi Arabia
2022-02-24DKHILMohamedALHAFIDHWejdanALQURAISHYSalehALOTAIBIMashaelBANAEEMManalALSALEHThekraandABDELGABERRewaida
DKHIL Mohamed A., ALHAFIDH Wejdan, AL-QURAISHY Saleh, ALOTAIBI Mashael,BANAEEM Manal, ALSALEH Thekra, and ABDEL-GABER Rewaida
Morphological and Phylogenetic Studies of a Copepod Species,Ho and Lin (2007), Infectingin Saudi Arabia
DKHIL Mohamed A., ALHAFIDH Wejdan, AL-QURAISHY Saleh, ALOTAIBI Mashael,BANAEEM Manal, ALSALEH Thekra, and ABDEL-GABER Rewaida*
,,,11451,
Dammam City is one of the gorgeous coastal areas in the Arabian Gulf of Saudi Arabia. The present study aimed to ex- amine one of the copepod species infecting the rosy goatfish that represents a highly consumed fish species by the local population in the Arabian Gulf.The copepod species isolated from the infected fish specimens belong to the family Taeniacanthidae and was iden- tified asHo and Lin (2007), primarily based on its morphological, morphometric, and ultrastructural characteris- tics, especially the structures of the dorsal cephalic area, segmentation of the first antenna, the absence of the maxilliped claw in the fe- male specimens, and the setation and spinulation of the legs 2–4 for the adult females are of great significance in the taxonomic iden-tification. Thegene sequence was analyzed to ensure the precise identity and exact taxonomic status of the copepod species.The result showed that thiscopepod species belong to Taenicanthidae and closely related to(gb| JF781550.1) in the same taxon. More details on the specificity of the goatfish forspecies and identifying these parasitic taxa using molecular analysis are given in the present study.
Taenicanthidae;species; goatfish; parasite specificity;gene
1 Introduction
Goatfishes are tropical marine perciform fish belonging to the family Mullidae, including more than 60 species (Ui-blein, 2007). The rosy goatfishis considered one of the essential goatfish species in the Ara- bian Gulf used as seafood at fish markets in Saudi Arabia. Despite its economic and ecologic importance, the ichthy- oparasitological problemsrelated toare in general scarce for this region, particularly those as- sociated with parasitic copepods that may affect them. Someresearch projects have focused on copepod parasites of goatfishes (Izawa, 1976; Dojiri and Cressey, 1987; Ho., 1999; Ho and Lin, 2007; Anh Tuan., 2015; Paschoal., 2016; Cardoso., 2017; Soler-Jiménez., 2019; Abdel-Gaber., 2020a, b, c). Taeniacanthidae Wilson, 1911 is a unique copepod family within the order Poeci- lostomatoida with either parasitic members on marine fish or associated with sea urchins (Dojiri and Humes, 1982; Boxshall and Halsey, 2004). Taeniacanthids exhibit a high degree of host specificity at both the generic and specific levels (Boxshall and Halsey, 2004). Copepods of the genusare usually parasitic on the gills, nostrils, and bran- chial cavities of their fish hosts (Dojiri and Cressy, 1987; Ho and Lin, 2007).was proposed by Wilson (1911) and established as its type-species, whichwas formerly known asHeller, 1865. How- ever, due to his inclusion of other species ofvon Nordmann, 1832, which created confusion in diagno- sing the newly erected genus, Ho (1969) rejected. This decision was followed by Kabata (1979), Balarman (1983), and Pillai (1985). Nevertheless, due to the disco- very of four other taeniacanthids that share certain deri- ved character states with, Dojiri and Cressey (1987) resurrected the genusand redefined it accord- ingly.
The identification and classification of copepods are fun- damentally based on their morphological and anatomical features (Huys and Boxshall, 1991; Ho, 2001; Boxshall and Halsey, 2004), which is very limited due to the relatively low number of the copepodologists (Hamza., 2007; Ramdane, 2009). Therefore, to complement the study of the phylogenetic relationships among copepod families based on the morphological characters, a different approach em- ploying molecular data is necessary (Ferrari and von Vau- pel Klein, 2019). Many other genetic markers complementthose conventional approaches (Baek., 2016). Among various molecular markers, phylogenetic studies based on thegene sequences have proven to be useful for investigating the evolutionary history of the crustaceans (Graybeal, 1994) and other metazoans (Aguinaldo., 1997). The versatile systematic utility ofgene is due to different evolutionary rates among different regions of the gene (Mindell and Honeycutt, 1990) in conjunction with a large size and a conserved function (Olsen and Woese, 1993). Due to such advantages,gene se- quence help to assess the phylogenetic relationships among copepod families containing highly modified morphologi- cal characters. Huys. (2007) found that nuclear ribo- somal genes ofandinclude semi-conserved domains that intersperse with divergent regions, allowing for a wide range of taxonomic levels in phylogenetic re-construction. Few mitochondrial genomes of copepods have been published so far (Wang., 2011; Easton., 2014), although others claim that copepod mitochondrial genomes retained pancrustacean features and maintained calanoid-specific patterns (Kim., 2013), or adapted to harsh environments by mitogenome rearrangements (Ca- meron., 2007; Ki., 2009).
There is, however, little information on the copepods in Saudi Arabia. Therefore, this study was designed to pro- vide the first report for the parasiteHo and Lin, 2007 ininhabiting Dam- mam City, Saudi Arabia. The copepod species was identi- fied and characterized through morphological and mole- cular analyses.
2 Materials and Methods
2.1 Fish Collection
A total of 80 specimens of the rosy goatfish,(Family: Mullidae), were randomly collect- ed during the period of January–October 2020 from the Arabian Gulf, Dammam City, Saudi Arabia. They were tran- sported to Parasitology Research Laboratory at the Zoo-logy Department, College of Science, King Saud University, Riyadh, Saudi Arabia. Fish were thoroughly tested for pa- rasitic species, according to the technique proposed by Ra- vichandran. (2007).
2.2 Parasitological Studies
Recovered parasites were carefully removed from the gill region of infected fish under a stereo-dissecting microscope (Nikon SMZ18, NIS ELEMENTS software), washed in a sterile 0.9% saline solution, and preserved for further de- tailed examination in 70% ethanol. The preserved samples were cleared for 24h with lactophenol,then the different body parts and appendages were examined. All drawings were made with the aid of a camera lucida. Using Olympus ocular micrometer, the different body parts of the selected copepods were measured. The average values with the range were given in parentheses. Morphological terminology fol- lows the guidelines of El-Rashidy and Boxshall (2014).Sewell’s style (1949) was adopted for the armature formu- la of the swimming legs, in which the spines and setae are denoted by Roman and Arabic numerals, respectively.
2.3 Molecular Analysis
2.3.1 DNA extraction and Polymerase chain reaction (PCR)
The genomic DNA was extracted from the preserved copepod samples using DNeasy tissue kit©(Qiagen, Hilden, Germany) following the recommended protocol. The qua- lity and purity of DNA were quantified with a NanoDrop ND-1000 spectrophotometer (Thermo Fischer Scientific, Wilmington, DE, USA). Thegene was targeted and amplifiedpolymerase chain reaction (PCR) and sub- sequently sequenced. PCR amplification was carried out using the same protocol and primers of the previous study of Huys. (2012). PCR amplicons were visualized on 1.5% TBE agarose gel stained with SYBR Safe DNA gel stain (Thermo Fisher Scientific, Ottawa, ON, Canada) by UV transilluminator.
2.3.2 Sequence alignment and Phylogenetic analysis
The amplicons were sequenced bidirectionally with ABI PRISM 310TMDNA Analyzer (Applied Biosystems, Fos- ter City, CA), then were analyzed using BLASTn, and align- ed using ClustalW multiple alignments (Thompson., 1994) implemented in BioEdit Sequence Alignment Editor ver. 5.0.9 (Hall, 1999). Phylogenetic analysis was conduct- ed using the method of Maximum Parsimony (MP) focus- ed on uniform rates among sites of the Tamura-Nei model (Tamura and Nei, 1993) on Molecular Evolutionary Gene- tics Analysis (MEGA) software version 7.0 (Kumar., 2016). This analysis was performed using a heuristic search strategy of Nearest-Neighbor-Interchange (NNI) with ran- dom addition sequences followed by tree-bisection recon- nection branch-swapping (TBR) with 1000 bootstrap rep- licates (Felsenstein, 1985) under appropriate substitution Nucleotide models.
3 Results
Twelve out of eighty (15%) specimens of the examined rosy goatfish,, were naturally infect- ed by a female copepod parasite known asHo and Lin, 2007 in the gills region.
3.1 Microscopic Examination
3.1.1 Adult females (Fig.1)
Body flattened. Prosome longer than Urosome, compri- sing of broad cephalothorax of about 40% of the total bodylength and four well-separated pedigers that distinctly broad. Urosome comprising of genital somite and four free abdo- minal somites without ornamentation. Genital somite with convex lateral margins bearing three long setae. Caudal ra-mus provided with six setae (four short and two long). Egg- sac oblong with large eggs.
3.1.2 Dimensions
Body length 1.81 (1.58–2.1) (excluding setae on caudal rami); cephalothorax 0.58 (0.56–0.63)×0.75 (0.71–0.78); urosome length 0.45 (0.39–0.49); genital somite wider than long 0.163 (0.150–0.172)×0.236 (0.197–0.243); fourabdominal somites 0.092 (0.089–0.095)×0.140 (0.132–0.145), 0.085 (0.080–0.089)×0.134 (0.127–0.137), 0.080(0.075–0.082)×0.107 (0.097–0.111), and 0.055 (0.046–0.059)×0.113 (0.110–0.118), respectively, from anterior to posterior; caudal rami length 0.070 (0.065–0.078), width 0.037 (0.031–0.039); and egg sac 0.632 (0.618–0.651)×0.341 (0.335–0.367).
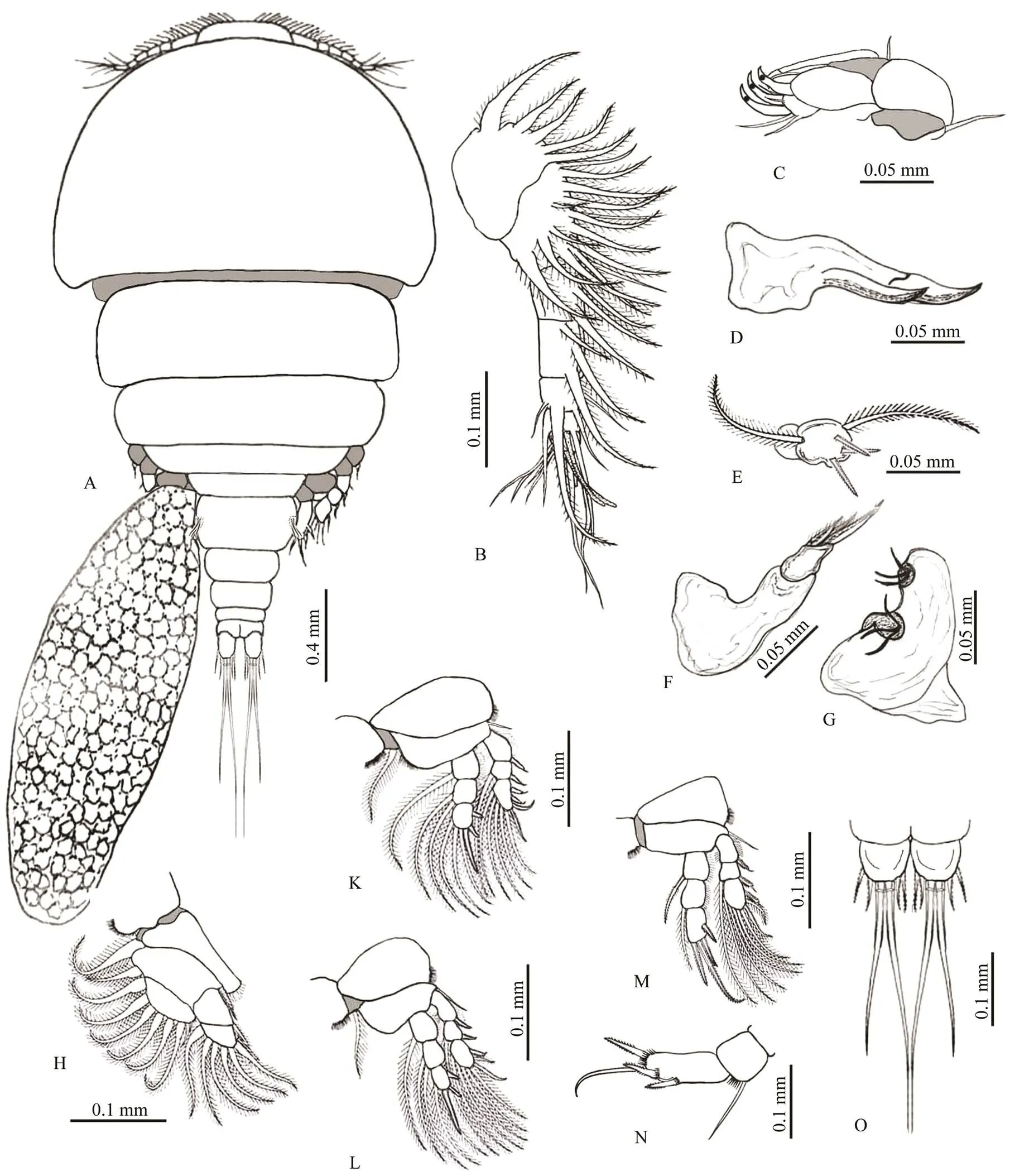
Fig.1 Irodes parupenei. A, the whole body of an adult female; B, antennule; C, antenna; D, mandible; E, maxillue; F, maxilla; G, maxilliped; H, leg 1; K, leg 2; L, leg 3; M, leg 4; N, leg 5; O, caudal rami.
3.1.3 Description
Rostral area broadly protruded anteriorly on the apex of the cephalothorax. First antenna (antennule) 6-segmented, with an armature formula of 20, 4, 3, 4, 2+1 aesthetasc, and 7+1 aesthetasc. Second antenna (antenna) uniramous, 4-segmented, proximal segment (coxobasis) largest and car-rying one seta, then strongly flexed outwardly at the junc- tion between the second and third segment. The second one carries a simple seta and the third one has one process and one curved claw, while the terminal segment consists of two claws and four setae. Mandible with two blades of serrate anterior edge. First maxilla (maxillule) lobate with five setae. Second maxilla (maxilla) 2-segmented, and the distal segment provided with three spines. Maxilliped 2- segmented with distal one carrying six setae. Legs 1–4 bi- ramous, Leg 1 with 2-segmented exopod and endopod. Legs2–4 with 3-segmented exopod and endopod. Armature for- mulae of legs 1–4 as:
Leg 1:exopod I-0;0–9 endopod 0–1;0–7
Leg 2:exopod I-0;I-1;III,I,5 endopod 0–1;0–2;II,I,3
Leg 3:exopod I-0;I-1;II,I,5 endopod 0–1;0–2;II,1,2
Leg 4:exopod I-0;I-1;II,I,5 endopod 0–1;0–1;I,III
Leg 5 segmented in two; the proximal segment with one seta, and the distal segment with three spines and one seta. Leg 6 with three setae located in the area of the egg sac attachment area.
3.1.4 Adult males
Adult males were not observed in this study.
3.2 Molecular Analysis
A total of 1442bp with 49.4% GC content was evalu-ated and deposited in GenBank under the accession number MW148591.1 for thegene of the existing taenia- canthid Copepoda species. Phylogenetic analysis was performed by aligning the partial and complete sequences of thegene with 29 taxa representing six copepod orders using a maximum likelihood method (Table 1, Fig.2).
The results showed that thegene sequences of this taeniacanthid species revealed 90.12%–91.18% identity with Harpacticoida, 89.21% with Siphonostoma- toida, 89.92% with Misophrioida, 90.21%–90.95% with Cy- clopoida, 87.51%–87.85% with Calanoida, and 96.38%–98.75% with Poecilostomatoida (Table 1). Among Poeci- lostomatoida, the present species bears 96.38%–98.75% similarity with taxa of Taenicanthidae, 97.36%–98.34% with Bomolochidae, and 97.78% with Umazuracolidae.
Among Poecilostomatoida, the maximum identity (98.75%) with the lowest divergent value was recorded between the present taeniacanthid species and(gb| JF 781550.1), followed by(97.92%,gb| KR048748.1),(97.78%, gb|JF781548.1),(97.71%, gb| JF78 1549.1),(97.23%, gb| JF781547.1),(97.16%,gb| KT030267.1), and(96.38%, KR048746.1) (Table 1). The overall mean distance between the studied species and other taeniacanthids was 0.02.
The ME tree showed that the cluster containing all of the copepod species was divided into two separate and dis-tinct clades (Fig.2). The first clade contains some copepod species belonging to Harpacticoida (representing by Arges- tidae, Dactylopusiidae, Pseudotachidiidae, Ectinosomati- dae, Zosimeidae, and Aegisthidae), Siphonostomatoida (re- presented by Kroyeriidae), Misophrioida (represented by Speleophriidae), Oithonidae, Schminlapinellidae, and My- tilicolidae of the order Cyclopoida, and Calanoida (repre- sented by Arietellidae, Clausocalanidae, and Kyphocalani-dae). The second clade comprises the other copepod spe- cies belonging to the Taeniacanthidae, Bomolochidae, and Umazuracolidae families of the order Poecilostomatoida. The ME tree revealed the studied species is closely related to(gb| JF781550.1) in the same taxon.
4 Discussion
Parasitic copepods utilize an extraordinary range of hosts, occurring on virtually every available phylum in the ma- rine environment (Boxshall and Halsey, 2004). Little in- formation is known about the parasitic fish copepods in the Arabian Gulf (Ho, 2001; Hamza., 2007; Ramdane, 2009; Abdel-Gaber., 2020a, c). Thus, the current study focused on investigating copepod parasites infecting the goatfish(Mullidae) from the coasts of the Arabian Gulf in Dammam City, Saudi Arabia. Mem- bers of thegenus have high host specificity to va- rious goatfish species that inhabited different geographical locations of India, Kenya, Kuwait, China, and Vietnam (Uma Devi and Shyamasundari, 1980; Dojiri and Cressey, 1987; Ho and Lin, 2007).

Table 1 Copepoda species used in the phylogenetic analysis of Irodes parupenei for their corresponding 18S rRNA gene region
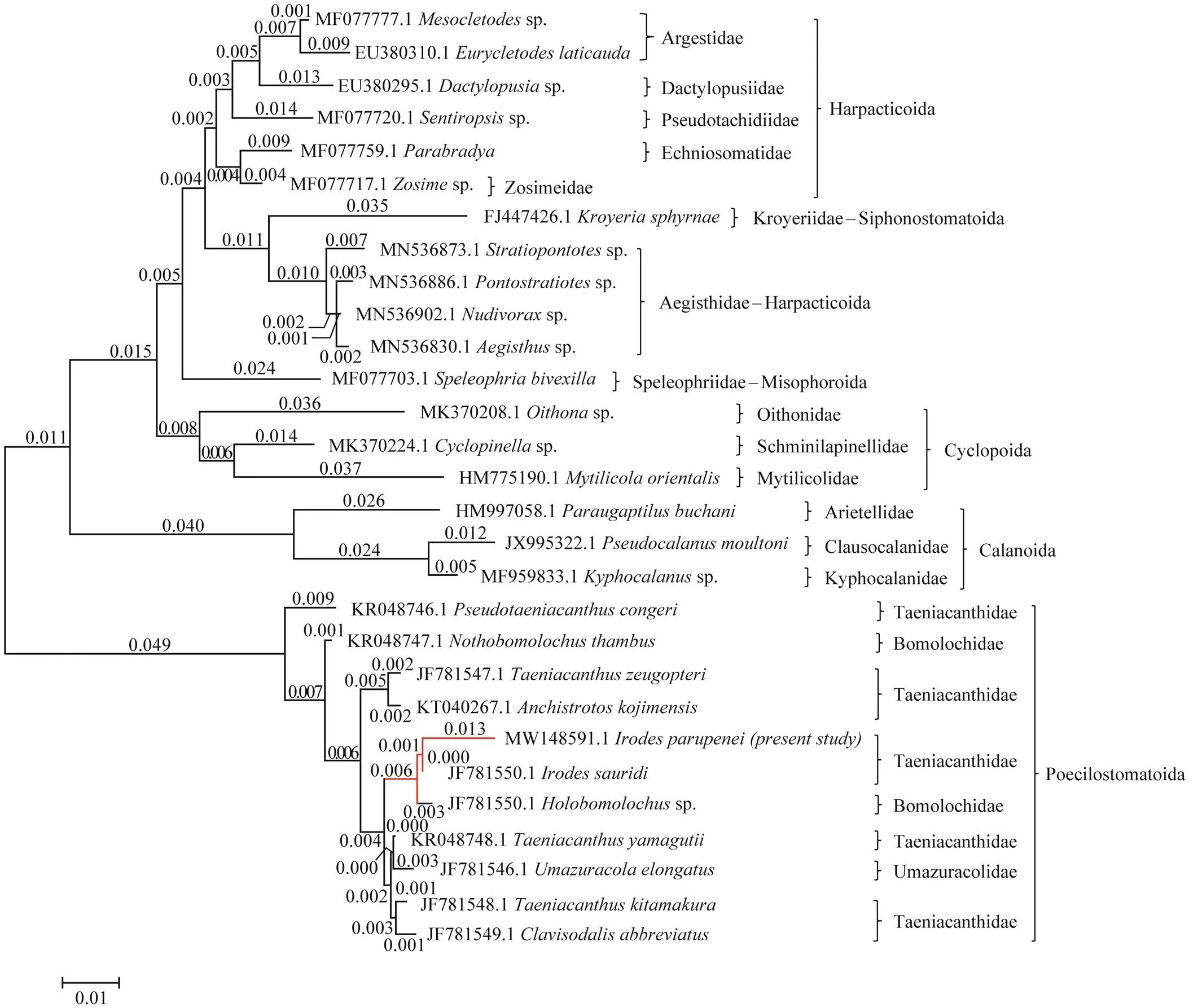
Fig.2 Molecular phylogenetic analysis by maximum likelihood method based on the Tamura-Nei model. The tree with the highest log likelihood (−5881.50) is shown. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach and then selecting the topology with a superior log-likelihood value. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site (next to the branches).
The taeniacanthid copepods species described herein agree with the previous illustrations ofgiven by Ho and Lin (2007) from the marine goatfish’ no- strilsandlanded at Ma-gong Fishing Port in Penghu County of Taiwan, Chi- na by their external morphology and host preferences. It differs from the previously described species in the type of host species and its geographical distribution. This study indicates that those species are synonymous.
The currentspecies can be differentiated from the other congeners of thegenus with the most related host species, such asYamaguti, 1954 isolated from the gills of,,, and, in the presence of nine elements on the terminal ex- opodal segment for the second leg (eight in), the presence of three spines and one setae on the distal segment of the fifth leg (four short setae in), and the anal segment without ornamentation (the short row of spinules near insertion of each caudal ramus in); and differed fromPillai, 1963 collected fromandby hav-ing minute outermost spine of terminal endopod segment of leg 4 (. conspicuous in), and the ventral surface of the anal segment with four rows of spinules in the anterior portion and a row of spinules near insertion of each caudal ramus (. absent in).
In addition, there are some differences between the pre- sentspecies and the other congeners infecting dif- ferent host types, such asDojiri and Cressey (1987) collected fromandby the presence of two sclerotized ridges forming V-shaped structure in the rostral area (. absent in), the second segment of antennule with 5 spi- nes (. 4 in), the appearance of strongly curved spine (resembling hooks) of the second segment and out- ermost spine of third exopodal segment of leg 3 (. notcurved in), almost equal width of thoracic seg-ments bearing legs 2 and 3 (. distinct varying size in), and the presence of a single row of minute spi- nules near insertion of each caudal ramus (. unornament- ed in); fromUma Devi and Shya- masundari (1980) infectedby hav- ing three-segmented antenna with four spiniform setae and one slender setae (that observed in); fromYamaguti, 1939 in the presence of 4-segment- ed abdomen (3-segments in), and the pre- sence of nine setae in the terminal exopodal segment of leg 1 (7 in); fromHeller, 1865 that isolated from,,,,,,,, and; by having two small rounded pro- cesses at antero-ventral region and a large bifid sclerotized part bearing transverse shelf-like protuberance (. absent in), and the presence of 5 spines on the second segment of antennule (. 4 in).
Until now, all valid species ofcan be identifiedfrom their congeners using the generic features in the avail- able key of Ho. (1999). However, mostspecies are challenging to identify because of the morphological similarities and the lack of detailed descriptions. Thus more detailed researches on both morphological and molecular analyses are needed for accurate identification. Copepods are exceptional arthropods because, with only a few com- plete mitogenomes published, they already present many deviations from the reference gene order (Feng., 2016; Su., 2016). The current study focused on the genetic markerfor precise identification of the recover- ed taeniacanthid species, which is consistent with Dippenaar (2009), Huys. (2009), Song. (2009), and Cepeda. (2012). The results indicated that the analysis of the DNA sequence for the target genes offers invaluable in- formation on the taxonomic position of copepod species.
According to Huys. (2002) and Ho. (2003), copepods are classified into three infraorders, Progymno- plea Lang, 1948, Gymnoplea Giesbrecht, 1892, and Podo- plea Giesbrecht, 1892. The current dendrogram was split into two clades representing copepod species by the last two infraorders of Gymnoplea and Podoplea. Herein, the basal taxon of Misophrioida and Calanoida in Neocopepods wasobserved. In addition to being a sister-group to the mem- bers of Podoplea, these results agreed with the previous data of Ho (1990), Huys and Boxshall (1991), Ho (1994), Ho. (2003), Blanco-Bercial. (2011), and Huys. (2012) indicating that these orders are the most primitive ones within Neocopepoda and Podoplea.
Previously, Dahms (2004), Jenner (2009), and Schizas. (2015) reported that the taxa of copepods within the Podoplea infraorder diversified into two clades: MHPSM- clade including Monstrilloida, Harpacticoida, Poecilosto- matoida, Siphonostomatoida, Mormonilloida; and MCG- clade was including Misophrioida, Cyclopoida, and Gelyel-loida. The current phylogeny represented those two clades and Monstrilloida, Mormonilloida, and Gelyelloida were not considered. Additionally, Walter and Boxshall (2019) re-ported that Poecilostomatoida comprises more than sixty families. Our molecular study provided nodal support for the monophyly of the bomolochiform complex. In 1987, Dojiri and Cressey mentioned the phylogenetic relationship within the bomolochiform complex consists of three close-ly related families including Bomolochidae, Taenicanthidae, and Tuccidae, while there is a close relationship between the former two families. Huys. (2012) assumed that Umazuracolidae belongs to this group of families. Herein, Umazuracolidae is related to the fish-parasitizing families of Bomolochidae and Taeniacanthidae that referred to a close similarity in the morphology of the antenna and mou- thparts as mentioned by Dojiri and Cressey (1987).
There is a sister-group of the Bomolochidae+Taeniacan- thidae in the current study, which agreed with Kabata (1979), Dojiri and Humes (1982), and Humes and Dojiri (1984). Both of these families have a mosaic of plesiomorphic and apomorphic characters based on the setation of the first an- tenna, the structure and shape of the rostral area, the an- tenna of the female specimens, the structure of prosome, the location and shape of maxilliped, and the spinulation oflegs 2–4. In the present study, the monophyly of Taeniacan-thidae was not supported by strict consensus due to thespecies that appeared to be closely related tosp., which is consistent with the observation of Huys. (2012). Herein, Taeniacanthidae includes five genera,,,,, and. By comparing thegene query, the current copepod parasite was detected to be a dis-tinct species related closely to(JF781550.1).
It can be concluded that the present study provided va- luable information about a taeniacanthid copepod speciesHo and Lin, 2007, which can infectrosy goatfish. Future studies on the func-tions of other genes are necessary to understand better about this species.
Acknowledgement
This study was supported by the Researchers Support- ing Project of King Saud University, Riyadh, Saudi Arabia (No. RSP-2021/25).
Abdel-Gaber, R., Alghamdi, M., Abu Hawsah, M., Ali, S., and Yehia, R.S., 2020a. Light and scanning electron microscopic studies of(Siphonostomatoida: Ca- ligidae) infecting the rosy goatfish., 12 (2): 282-287.
Abdel-Gaber, R., Al-Quraishy, S., Dkhil, M.A., Alghamdi, M., Alghamdi, J., and Kadry, M., 2020b. Morphology and phylo- geny ofShiino, 1956 (Hexanauplia: Taeniacanthidae), a copepod infecting the gills of rosy goatfish(Mullidae) in the Arabian Gulf., 19: 1409-1420.
Abdel-Gaber, R., Al-Quraishy, S., Dkhil, M.A., Elamin, M., and Alghamdi, M., 2020c. Morphological analysis ofvon Nordmann, 1832 (Copepoda: Caligidae) from the rosygoatfish(Mullidae)., 83 (11): 1369-1380.
Aguinaldo, A.M., Turbeville, J.M., Linford, L.S., Rivera, M.C., Garey, J.R., Raff, R.A.,., 1997. Evidence for a clade of nematodes, arthropods, and other moulting animals.,387: 489-493.
Anh Tuan, D.N., Sang, T.Q., and Binh, D.T., 2015. Parasites of goatfishes (spp.) in Khanh Hoa Province, Vietnam,preliminary results., 2015(Special issue):10-15.
Baek, S.Y., Jang, K.H., Choi, E.H., Ryu, S.H., Kim, S.K., Lee, J.H.,., 2016. DNA barcoding of metazoan zooplanktoncopepods from South Korea., 11(7): e0157307.
Balarman, S., 1983. Discussion on the validity of the genera of Taeniacanthidae (Crustacea, Copepoda). In:Rabindranath, P., ed., The Aquarium, Trivan-drum, 207pp.
Blanco-Bercial, L., Bradford-Grieve, J., and Bucklin, A., 2011. Molecular phylogeny of the Calanoida (Crustacea: Copepoda)., 59: 103-113.
Boxshall, G. A., and Halsey, S. H., 2004.. The Ray Society, London, 966pp.
Cameron, S.L., Johnson, K.P., and Whiting, M.F., 2007. The mitochondrial genome of the screamer louse(Phthiraptera: Ischnocera): Effects of extensive gene rearrange- ments on the evolution of the genome., 65: 589-604.
Cardoso, L., Martins, M.L., Cadorin, D.I., Bonfim, C.N.C., and de Oliveira, R.L.M., 2017. First record of(Copepoda: Bomolochidae) in Brazil, ectoparasite of the spotted goatfish(Actinopterygii: Mullidae)., 39 (2): 251-258.
Cepeda, G. D., Blanco-Bercial, L., Bucklin, A., Berón, C. M., and Viñas, M. D., 2012. Molecular systematic of three species of(Copepoda, Cyclopoida) from the Atlantic Ocean: Com- parative analysis using 28S rDNA., 7 (4): e35861.
Dahms, H. U., 2004. Exclusion of the Polyarthra from Harpacticoid and its reallocation as an underived branch of the Co- pepoda (Arthropoda, Crustacea)., 1 (1): 29-51.
Dippenaar, S.M., 2009. Estimated molecular phylogenetic rela- tionships of six siphonostomatoid families (Copepoda) symbiotic on elasmobranchs., 82: 1547-1567.
Dojiri, M., and Cressey, R. F., 1987. Revision of the Taeniacan- thidae (Copepoda: Poecilostomatoida) parasitic on fishes and sea urchins., 447: 1-250.
Dojiri, M., and Humes, A. G., 1982. Copepods (Poecilostomata: Taeniacanthidae) from sea urchins (Echinoidea) in the Southwest Pacific., 74: 381-436.
Easton, E.E., Darrow, E.M., Spears, T., and Thistle, D., 2014. The mitochondrial genomes ofand(Harpacticoida: Miraciidae) reveal similarities between the copepod orders Harpacticoida and Poecilostoma- toida.,538: 123-137.
El-Rashidy, H.H., and Boxshall, G.A., 2014. A new parasitic co- pepod (Cyclopoida: Bomolochidae) from a ponyfish (Leiogna-thidae) caught in Egyptian Mediterranean waters, with a re- view of hosts and key to species of., 87: 111-126.
Felsenstein, J., 1985. Confidence limits on phylogenies: An ap- proach using the bootstrap.,39: 783-791.
Feng, H., Wang, L. X., Huang, J., Jiang, J., Tang, D., Fang, R.,., 2016. Complete mitochondrial genome of(Copepoda: Poecilostomatoida)., 27: 2960-2962.
Ferrari, F. D., and von Vaupel Klein, J. C., 2019. Rhabdomoplea, a new superorder for the thaumatopsylloid copepods: The con- sequence of an alternative hypothesis of copepod phylogeny., 92: 177-188.
Giesbrecht, W., 1892. Systematik und Faunistik der pelagischen Copepoden des Golfes von Neapel., 19: 1-831.
Graybeal, A., 1994. Evaluating the phylogenetic utility of genes–A search for genes informative about deep divergences among vertebrates., 43: 174-193.
Hall, T.A., 1999. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT., 41: 95-98.
Hamza, F., Boxshall, G., and Kechemir-Issad, N., 2007. A new species ofNune-Ruivo, 1954 (Copepoda: Hat- schekiidae) parasitic on(Cadenat) of Al- geria., 67: 119-124.
Heller, C., 1865. Crustaceen. In:.Karl Gerold’s Sohn Wien, Vienna, 388 (in German).
Ho, J. S., 1969. Copepods of the family Taeniacanthidae (Cyclo- poida) parasitic on fishes in the Gulf of Mexico., 19: 111-130.
Ho, J. S., 1990. Phylogenetic analysis of copepod orders., 10: 528-536.
Ho, J.S., 1994. Copepod phylogeny: A reconsideration of Huys & Boxshall’s ‘parsimony versus homology’., 292 (293): 31-39.
Ho, J.S., 2001. Why do symbiotic copepods matter?,453/454: 1-7.
Ho, J.S., and Lin, C.L., 2007. Two new species of taeniacan- thid copepods (Poecilostomatoida) parasitic on marine fishes of Taiwan., 67: 73-80.
Ho, J. S., Dojiri, M., Gordon, H., and Deets, G. B., 2003. A new species of Copepoda (Thaumatopsyllidae) symbiotic with a brittle star from California, USA, and designation of a new or- der Thaumatopsylloida., 23: 582- 594.
Ho, J.S., Kim, I.H., and Sey, O., 1999. New species of(Copepoda, Taeniacanthidae) parasitic on the goatfish from Ku- wait, with a key to the species of., 8: 123-129.
Humes, A.G., and Dojiri, M., 1984. Current concepts of taenia- canthid copepods associated with Echinoidea., 7: 258-266.
Huys, R., and Boxshall, G. A., 1991.. The Ray Society, London,468pp.
Huys, R., Fatih, F., Ohtsuka, S., and Llewellyn-Hughes, J., 2012. Evolution of the bomolochiform superfamily complex (Cope- poda: Cyclopoida): New insights fromand morphol- ogy, and origin of umazuracolids from polychaete-infesting ancestors rejected., 42: 71-92.
Huys, R., Llewellyn-Hughes, J., Conroy-Dalton, S., Olson, P. D., Spinks, J. N., and Johnston, D. A., 2007. Extraordinary host switching in siphonostomatoid copepods and the demise of theMonstrilloida: Integrating molecular data, ontogeny and anten- nulary morphology., 43: 368-378.
Huys, R., Lopez-Gonzalez, P. J., Roldan, E., and Luque, A. A., 2002. Brooding in cocculiniform limpets (Gastropoda) and fa- milial distinctiveness of the Nucellicolidae (Copepoda): Mis- conceptions reviewed from a chitonophilid perspective., 75 (2): 187-217.
Huys, R., MacKenzie-Dodds, J., and Llewellyn-Hughes, J., 2009. Cancrincolidae (Copepoda, Harpacticoida) associated with land crabs: A semiterrestrial leaf of the ameirid tree., 51: 143-156.
Izawa, K., 1976. A new parasitic copepod,gen. et sp. nov. (Cyclopoida: Bomolochidae), from a Ja- panese goatfish., 23: 289-298.
Jenner, R. A., 2009. Higher-level crustacean phylogeny: Consen- sus and conflicting hypotheses., 39 (2): 143-153.
Kabata, Z., 1979.. The Ray Society, London, 468pp.
Ki, J.S., Park, H.G., and Lee, J.S., 2009. The complete mito- chondrial genome of the cyclopoid copepod: A highly divergent genome with novel gene order and aty- pical gene numbers., 435: 13-22.
Kim, S., Lim, B.J., Min, G.S., and Choi, H.G., 2013. The com- plete mitochondrial genome of Arctic(Co-pepoda, Calanoida) reveals characteristic patterns in calanoid mitochondrial genome., 520: 64-72.
Kumar, S., Stecher, G., and Tamura, K., 2016. MEGA7: Molecu- lar Evolutionary Genetics Analysis version 7.0 for bigger data- sets., 33: 1870-1874.
Lang, K., 1948. Copepoda ‘Notodelphyoida’ from the Swedish west-coast with an outline on the systematics of the Copepods.14): 1-36.
Mindell, D., and Honeycutt, R.L., 1990. Ribosomal RNA in ver-tebrates: Evolution and phylogenetic applications., 21: 541-566.
Olsen, G.J., and Woese, C.R., 1993. Ribosomal RNA: A key to phylogeny.,7:113-123.
Paschoal, F., Pereira, A.N., and Luque, J.L., 2016.sp. nov. (Copepoda: Philichthyidae) parasitic in the dwarf goatfishPoey, 1852 (Perciformes: Mul- lidae) in the South Atlantic Ocean., 4174 (1): 176-191.
Pillai, N. K., 1963. Copepods of the family Taeniacanthidae pa- rasitic on South Indian fishes., 6: 110-128.
Pillai, N. K., 1985.:. Zoological Society of India, Calcutta,900pp.
Pritchard, M. H., and Kruse, G. O. W., 1982.. The Harold W. Manter Laboratory. University of Nebraska Press, Lincoln, 141pp.
Ramdane, Z., 2009. Identification et écologie des ectoparasites Crustacés des poissons Téléostéens de la côte Est algérienne. Thèse de doctorat de l’Université Badji Moktar Annaba, 235pp.
Ravichandran, S., Ajith Kumar, T. T., Ronald Ross, P., and Mu- thulingam, M., 2007. Histopathology of the infestation of pa- rasitic isopodof the host fish., 2 (1): 68-71.
Schizas, N. V., Dahms, H. U., Kangtia, P., Corgosinho, P., and Ga- lindo, A. M., 2015. A new species ofClaus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Caribbean me- sophotic reefs with remarks on the phylogenetic affinities of Polyarthra., 11 (8): 789-803.
Sewell, R. B. S., 1949. The littoral and semi-parasitic Cyclopoida, Monstrilloida and Notodelphyoida., 9 (2): 17-199.
Soler-Jimenez, L.C., Morales-Serna, F.N., Aguirre-Macedo, M.L., McLaughlin, J.P., and Jaramillo, A.G., 2019. Parasitic co- pepods (Crustacea, Hexanauplia) on fishes from the lagoon flats of Palmyra Atoll, Central Pacific., 833: 85-106.
Song, C., Fang, J., Li, X., Liu, H., and Chao, C. T., 2009. Identi- fication and characterization of 27 conserved microRNAs in citrus., 230: 671-685.
Su, Y. B., Wang, L. X., Kong, S. C., Chen, L., and Fang, R., 2016.Complete mitochpndrial genome of(Cope- poda: Cyclopoida)., 27: 1503-1504.
Tamura, K., and Nei, M., 1993. Estimation of the number of nu- cleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees., 10: 512-526.
Thompson, J.D., Higgins, D.G., and Gibson, T.J., 1994. Clustal W: Improving the sensitivity of progressive multiple alignment through sequence weighting, position-specific gap penalties and weight matrix choice.,22: 4673-4680.
Uiblein, F., 2007. Goatfishes (Mullidae) as indicators in tropical and temperate coastal habitat monitoring and management., 3: 275-288.
Uma, D.V., and Shyamasundari, K., 1980. Studies on the cope- pod parasites of fishes of the Walter Coast: Family Taeniacan- thidae.,39 (2): 197-198.
von Nordmann, A., 1832.. G. Reimer, Berlin, 150pp.
Walter, T. C., and Boxshall, G., 2019. World of Copepods data- base.Vervoort, 1962. Accessed through: World Register of Marine Species. http://www.marinespecies. org/aphia.php?p=taxdetails&id=128611.
Wang, M. X., Sun, S., Li, C. L., and Shen, X., 2011. Distinc- tive mitochondrial genome of Calanoid copepodwith multiple large non-coding regions and reshuffled gene order: Useful molecular markers for phylogenetic and po-pulation studies., 12: 73.
Wilson, C. B., 1911. North American parasitic copepods belong- ing to the family Frgasilidae., 39: 263-400.
Yamaguti, S., 1939. Parasitic copepods from fishes of Japan. Part 5. Caligoida, III., 2: 443-487.
Yamaguti, S., 1954. Parasitic copepods from fishes of Celebes and Borneo., 3 (3): 137-160.
December 23, 2020;
February 10, 2021;
June 15, 2021
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2022
. E-mail: rewaida@sci.cu.edu.eg
(Edited by Qiu Yantao)
杂志排行

Journal of Ocean University of China的其它文章
- Probabilistic Methods for Reliability Assessment of Floating Structures Based on Extreme Wave Simulation
- Structure of the Potential Vorticity of an Explosive Cyclone over the Eastern Asian Region in Late November 2013
- Enzyme-Induced Carbonate Precipitation for the Protection of Earthen Dikes and Embankments Under Surface Runoff: Laboratory Investigations
- Isolation, Identification, and Quantitative Determination of Saponin in Apostichopus japonicus by HPLC-DAD
- Transcriptome Analysis of Pacific White Shrimp (Litopenaeus vannamei) Under Prolonged High-Salinity Stress
- Evaluation of the Bioavailability of Metals in Sediment from the Southern Coastal Wetland of the Qiantang Estuary by Using Diffusive Gradients in Thin Films Technique