海螺沟冰川退缩区原生演替生态系统中铅累积的历史记录
2022-02-19杨丹荔罗辑贾龙玉陈云飞
杨丹荔,罗辑,贾龙玉 ,陈云飞
1.内江师范学院沱江流域高质量发展研究中心,四川 内江 641100;2.中国科学院、水利部成都山地灾害与环境研究所,四川 成都 610041;3.成都理工大学地球科学学院,四川 成都 610059
环境问题由来已久,人们所普遍关注的重金属污染中,铅由于其生物毒性大且可通过大气的远距离运输而影响偏远和相对洁净的地区(Spokes et al.,1995),对全球尺度的生态安全产生危害而备受学者们关注。在欧洲,可检测到的大气铅污染早在6000年前就已开始,主要是由于土壤粉尘和农业活动。自工业革命以来,化石燃料燃烧、金属冶炼以及汽车尾气的排放等人类活动导致大气中的铅不断增加。国外不少学者利用冰芯(Rosman et al.,1993)、湖泊及海洋沉积物(Farmer et al.,1996;Renberg et al.,2002)、泥炭沼泽(Fancois et al.,2007)及树轮(Padilla et al.,2002)等研究大气铅的污染历史,其研究结果均指出在 1950年以后是铅沉降增强的时期,尤其是 1950—1980年是全欧洲最大的铅排放时期。同样在中国,尤其是在偏远且纯净的青藏高原区,已有不少学者监测到外源污染物对该地区生态环境产生的负面影响已经凸显,包括在该地区冰芯(李真等,2006)、湖泊沉积物(Bing et al.,2016)以及树轮(何咏梅等,2021)中均监测到铅污染在近几十年以来其浓度呈现出明显增加的趋势。但常规用以监测和记录重金属污染的手段有其各自的优点,但也有不可避免的缺陷(Alley,2010;Bacon et al.,2017)。在此基础上,不少学者越来越关注生态系统对重金属的富集和监测作用,包括利用苔藓监测欧洲大气铅污染(Stankovic et al.,2018);利用青藏高原东部林线样带中植物和土壤重金属的分布监测到铅浓度由东南向西北递减(Luo et al.,2014,2015;Tang et al.,2014,2015)。因此,生态系统对铅污染的响应,为铅污染的监测提供了方便可靠的条件。而建立在冰川退缩区的植被演替序列,不仅可以较为准确的测定重金属在生态系统中的吸收、迁移、周转和分配特征,进而探索其在原生演替序列逐级传递和富集的生物地球化学过程;还可测定原生演替序列上大气沉降输入的重金属通量,从而获得演替序列时间尺度上的重金属污染记录,反演一定时间范围内生态系统中重金属的输入情况。
位于青藏高原东缘贡嘎山东坡的海螺沟冰川退缩区,自小冰期结束以来发育了典型的土壤年代序列和完整的植被演替序列,其独特的自然条件和演化过程,为探索复杂环境中重金属的污染历史记录提供了天然的实验场所。不少学者已在该演替序列上开展了镉和汞的生物地球化学研究,揭示了镉和汞在该演替序列的富集过程,并反演出近120多年来大气镉和汞的污染历史(Wang et al.,2020a,2020b;佘佳,2014)。而关于铅在该演替序列的研究包括苔藓对大气铅污染的监测(Wang et al.,2019)、表层土壤铅的富集(杨丹荔等,2020)以及利用铅同位素对其来源示踪(Bing et al.,2014)等。但有关整个生态系统铅的富集特征和积累过程的研究还不够完善。因此,本文以海螺沟冰川退缩区为载体,研究各演替阶段生态系统中铅的富集特征并利用 1890—2017年生态系统铅的积累速率来反演该演替序列铅的污染过程。这对贡嘎山东坡生态系统重金属含量与贮量的研究具有重要意义,同时丰富了相关研究的方法和数据,为重建重金属的污染历史提供科学依据。
1 材料与方法
1.1 样地的设置
结合海螺沟冰川最早的科学记载资料(1930年)以及先锋树种定居(即冰川消退到树木幼苗发芽之间的时间)和冰川退缩区最大树木年龄的时间间隔确定各样地的冰川退缩时间,并通过树木年轮进行校正。根据2017年野外实地调查资料,分别在冰川退缩时间的 2017年(S0),2000年(S1),1980年(S2),1970年(S3),1958年(S4),1930年(S5)和1890年(S6)布设7个连续样地(图1)。S0为冰川退缩后形成的裸地。在冰川退缩后的第4年开始有先锋物种入侵,在S1样地,一些灌丛及乔木幼苗如沙棘(Hippophae rhamnoides)、柳树(Salixspp.)和冬瓜杨(Populus purdomii)等进入以黄芪(Astragalus membranaceus(Fisch.) Bung)为主的草本群落,形成演替初期的草灌阶段;S2—S4样地,冬瓜杨凭借较高的光合速率和较快的高生长优势,占据林冠上层,形成演替中期以冬瓜杨为优势种的落叶阔叶林群落;峨眉冷杉(Abies fabri)在冬瓜杨形成的郁闭环境下快速生长逐步成为群落优势种,到S5样地形成以冬瓜杨和峨眉冷杉为主的针阔混交林群落;随着峨眉冷杉进入主林层,喜阳的冬瓜杨生长受限并逐步退出群落,最终在S6样地形成以峨眉冷杉和麦吊云杉(Picea brachytyla)为主的针叶林顶极群落(杨丹丹等,2015)。
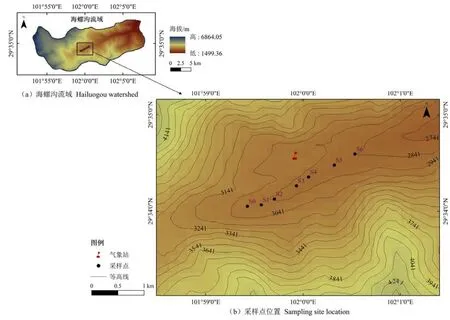
图1 研究区及采样点设置Figure 1 The studied area and sampling sites
1.2 野外取样及样品分析
作者于2017—2018年进行野外取样。由于S0样地为原生裸地,土壤尚未发育,仅取冰川末端堆积物细砂进行元素含量的测定。在 S1—S6各样地随机布设3个10 m×10 m的样方,记录样方内乔木层各优势种的名称、胸径、树高等信息,并采集样地内各优势种干、皮、枝、叶和根各部位的样品。根据实测的胸径和树高估算乔木层各优势种的生物量。有关该地区乔木层的生物量已在相关的研究中发表(杨丹丹等,2015)。在每个10 m×10 m的样方内,设置1 m×1 m的代表性小样方,记录样方内所有灌木、草本和苔藓的名字和数量,采用收获法分别获得样方内所有灌木、草本及苔藓,并将所有灌木、草本及苔藓的样品分别混合后记录其各自的鲜重。随后在设置0.5 m×0.5 m的小样方并挖一个土壤剖面,按照野外土壤发育性状,将土壤分为:Oe层,土壤有机质未完全分解的半分解层;Oa层,土壤有机质完全分解形成腐殖质的全分解层(土壤Oe层和Oa层为土壤O层);C层,母质层。由于该演替序列土壤发育时间较短,未见淀积层和淋溶层的形成。记录每层土壤的厚度,并挑出土壤中的根系和石砾,采用环刀法测定土壤的容重,然后分层采集土壤样品。将收集的所有样品带回实验室,植物样品置于 65 ℃烘干至恒定质量并测定其干质量,将灌木、草本、苔藓的干质量作为其生物量,同时用作样品元素分析。土壤样品经自然风干后,用玛瑙研钵粉碎至≤200目,植物等样品经粉碎或剪碎至长度≤2 mm。将处理好的植物样品和土壤样品分别经盐酸-硝酸-氢氟酸-高氯酸敞开消解和硝酸双氧水氢氟酸微波消解后,其全溶液通过 ICP-MS分别测定铅的含量。
1.3 数据分析
(1)表层土壤铅的富集系数,其计算公式为(Aloupi et al.,2001):
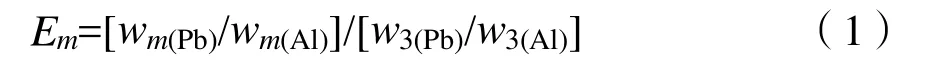
式中:
E——富集系数;
m——土层;
w3(Pb)/w3(Al)——C层铅含量与铝含量之比;
wm(Pb)/wm(Al)——土壤某层中铅含量与铝含量之比。
(2)生物转运系数,为植物某部位重金属含量与该植物细根中元素含量的比值,反映了植物不同部位对重金属的运移能力。其计算公式为:转运系数=地上部分各部位铅含量/细根铅含量(陈文德等,2009)。
(3)生物富集系数,为植物某部位某元素含量与土壤该元素含量之比,反映植物从土壤中摄取该元素的能力。当富集系数>1时,表征该元素在植物体内富集。其计算公式为:生物富集系数=植物各部位铅的含量/土壤铅的含量。(其中土壤铅的含量以各层的厚度为权重通过加权平均得到)(董林林等,2008)。
(4)生态系统各层铅的贮量及积累速率:植被各层铅的贮量通过生物量与其含量的乘积得到,土壤各层铅的贮量通过含量与厚度和容重的乘积得到。各层的积累速率为:

式中:
R——积累速率;
P——各演替阶段活体植被和土壤的铅贮量;
t——冰川退缩的时间;
i=1—6,即S1—S6样地。
本文的研究区位置图采用Arcgis软件绘制,数据分析及研究结果图采用 Origin 2020软件进行绘制,研究区的大气轨迹图通过 HYSPLIT后向轨迹模型进行绘制。
2 研究结果与分析
2.1 土壤铅的富集特征
以 S0样地所采集的冰川细砂中铅的含量作为该地区的背景值,土壤Oe层的铅含量在S4—S6样地高于背景值,土壤Oa层的铅含量在各样地均高于背景值,土壤C层的铅含量除S6样地外均低于背景值,且土壤各层铅的含量在 S2—S6样地主要表现为Oa层>Oe层>C层(图2)。此外,土壤Oe层铅含量为 14.69—52.03 mg·kg−1,Oa层为 33.00—68.47 mg·kg−1,且土壤Oe层和Oa层中铅含量的变化趋势较为一致,均随演替呈先增长后下降的趋势;土壤 C 层铅含量在 15.95—30.16 mg·kg−1,随演替的波动较小。

图2 不同样地土壤各层铅质量分数及表层土壤的富集系数Figure 2 Mass fraction of Pb in each soil layer and enrichment coefficient of surface soil
土壤Oe层和Oa层铅的富集系数随演替呈先增长后下降的趋势,且均在S5样地分别达最大值9.88和5.47,远高于富集系数的临界值1,说明该演替序列土壤中的铅表现出明显的表层富集,尤其是土壤Oe层在S5样地的富集系数接近10,显示出该演替序列土壤O层中的铅并非来源于成土母质,极大可能受凋落物的分解或大气铅沉降的影响导致含量较高。
2.2 活体植被铅的富集特征
2.2.1 优势乔木
从各样地优势乔木不同部位的铅含量来看(图3),沙棘表现为细根>皮>叶>中根>粗根>枝>干,柳树为细根>叶>中根>枝>皮>粗根>干,冬瓜杨为细根>枝>中根>叶>皮>粗根>干,峨眉冷杉为细根>枝>中根>叶>粗根>皮>干。可见,各优势乔木均表现为细根中的铅含量最高,干中的铅含量最低。此外,沙棘地下部分根的平均铅含量(0.32—4.6 mg·kg−1)比地上部分各部位平均铅含量(0.45—3.31 mg·kg−1)高 1.47倍,柳树地下部分(0.49—21.1 mg·kg−1)比地上部分(0.34—3.12 mg·kg−1)高 3.45倍,冬瓜杨地下部分(0.57—16.2 mg·kg−1)比地上部分(0.35—7.08 mg·kg−1)高 1.71倍,峨眉冷杉地下部分(0.60—7.4 mg·kg−1)比地上部分高(0.25—7.43 mg·kg−1)高 1.55倍。可见,各优势乔木不同部位的铅含量均表现为地下部分>地上部分。
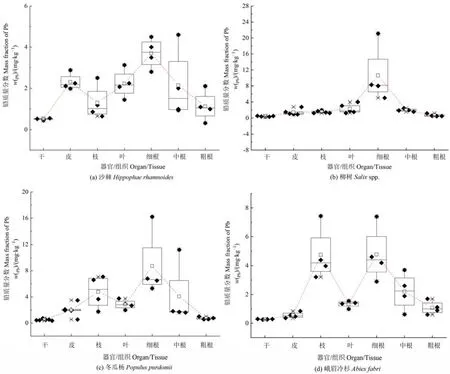
图3 各优势乔木不同部位的铅质量分数Figure 3 Mass fraction of Pb in different parts of dominant trees
为进一步说明铅在乔木各部位的迁移与富集,本文引入生物转运系数和生物富集系数反映植物各部位对铅的运移能力和富集程度。从乔木地上各部位的生物转运系数来看(图 4),沙棘皮的运移能力最高,柳树叶的运移能力最高,而冬瓜杨和峨眉冷杉枝的运移能力最高。此外,沙棘地上各部位的转运系数最高,其次为峨眉冷杉、冬瓜杨和柳树。但除峨眉冷杉枝的转运系数 (1.09)>1以外,其余优势乔木各部位的转运系数均<1。从乔木各部位的生物富集系数来看,除峨眉冷杉皮中铅的富集系数较高外,其余均在细根中的富集系数最高。此外,各优势乔木地下部分的富集系数均高于地上部分,且柳树的富集系数最高,其次为冬瓜杨、沙棘和峨眉冷杉。由此可见,各优势乔木地上部分的运移能力较低,而地下部分的富集程度均高于地上部分,根系所吸收的铅可能大部分被存留在根系组织中。
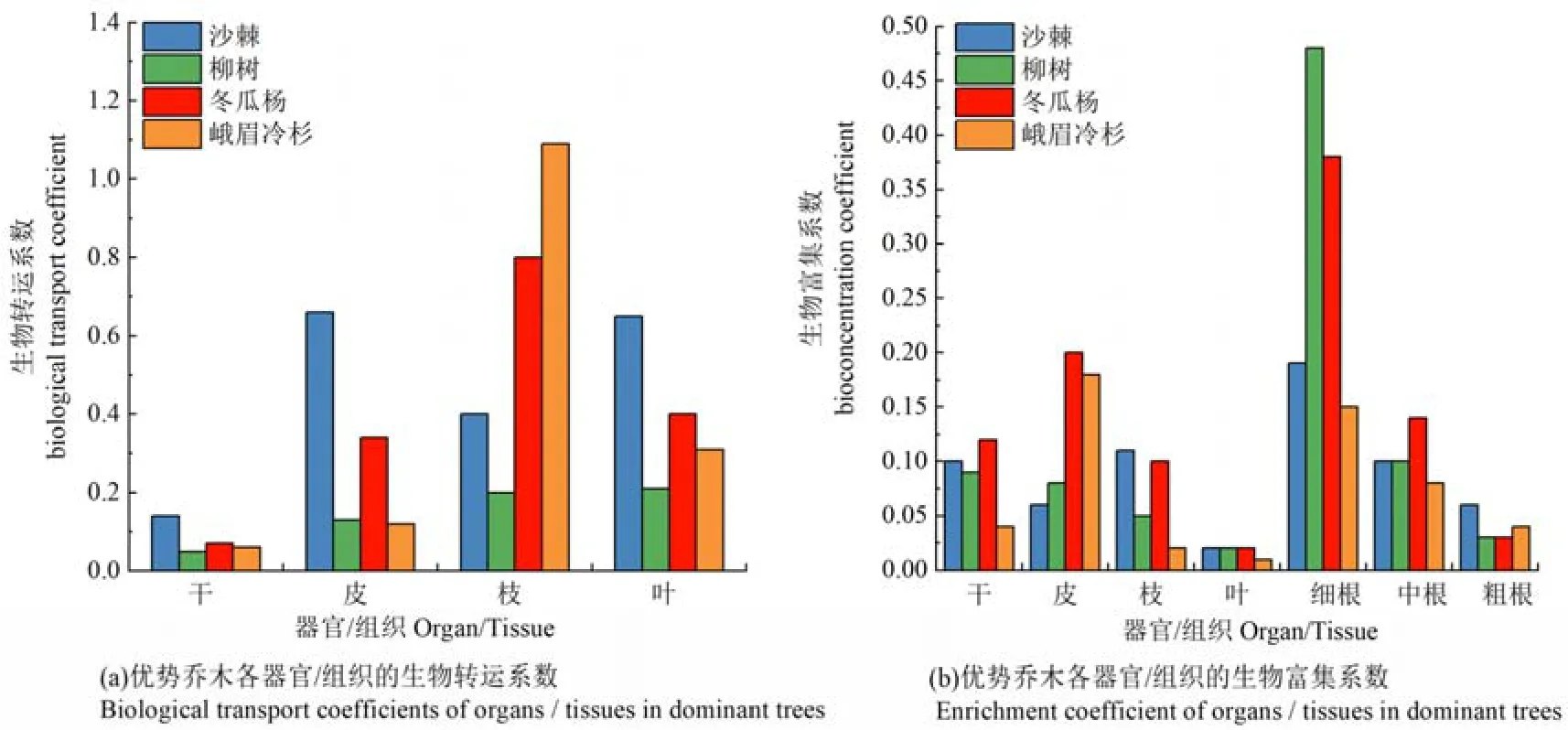
图4 各优势乔木不同部位铅的生物转运系数和生物富集系数Figure 4 Biological transport coefficient and bioaccumulation coefficient of Pb in different parts of dominant trees
2.2.2 林下植被层
灌木层枝铅的含量高于叶,但二者在整个演替序列的变化不大(图5);草本层铅的含量在S1样地最高达8.7 mg·kg−1,且随冰川退缩时间的增加呈先下降后增长的趋势;地被层(苔藓)铅的平均含量为17.23 mg·kg−1,尤其是S1样地地被层的铅含量高达31.62 mg·kg−1,并超过该样地表层土壤中铅的含量,且明显高于林下的其他植被层。可见,林下植被中地被层对铅的富集能力较强,可能会造成生态系统中铅等有毒重金属在其生物量中的积累。
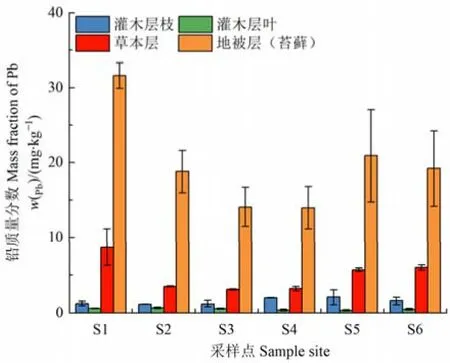
图5 林下各植被层铅质量分数Figure 5 Mass fraction of Pb in each undergrowth vegetation layer
2.3 生态系统铅的积累动态
生态系统铅的总贮量随演替呈波动增长的趋势(图6),从S1样地的64.64 kg·hm−2增长到S6样地的 227.16 kg·hm−2。其中,活体植被的铅贮量随演替从S1样地的0.01 kg·hm−2增长到S5样地的0.58 kg·hm−2,到S6样地略有下降,而土壤铅贮量从S1样地的64.83 kg·hm−2增长到S6样地的226.68 kg·hm−2。可见,各样地土壤铅贮量占生态系统总贮量的99%以上,是生态系统铅的最大储存库。

图6 生态系统活体植被与土壤铅的贮量Figure 6 Pb storage in living vegetation and soil of ecosystem
在活体植被中,乔木层铅贮量随演替逐渐增加,到S5样地达最大值0.35 kg·hm−2,其所占比例在各样地均达60%以上,尤其是S3样地所占比例达90%。其中,S1样地乔木层中沙棘的铅贮量最高,但随植被的演替,到S4样地冬瓜杨的铅贮量达0.27 kg·hm−2,约占整个活体植被铅贮量的76%;到演替末期形成以峨眉冷杉为主的针叶林后,峨眉冷杉的铅贮量为0.26 kg·hm−2,约占活体植被铅贮量的50%。可见,各优势乔木铅贮量的动态变化与其演替趋势是一致的。灌木层的铅贮量随演替从S1样地的0.02 g·hm−2增长到 S6 样地的 81.02 g·hm−2,但在整个演替序列中其铅贮量仅占活体植被铅贮量的5.35%。草本层铅贮量呈先增长后下降的趋势,并在S4样地达最大值 6.02 g·hm−2,其贮量占活体植被铅贮量的比重也非常小,仅为2.38%。地被层(苔藓)铅贮量的变化趋势与乔木层一致,均在S5样地达最大值0.18 kg·hm−2,其占活体植被铅贮量的比重最大可达36%。因此,乔木层和地被层是活体植被中铅的主要存贮单元,而灌木层和草本层的铅贮量较低且所占比重小,对铅在活体植被中的分配格局影响较小。
土壤铅总贮量随演替呈波动的增长趋势,在S2样地略有下降。土壤Oe层和Oa层的铅贮量随演替呈先增长后下降的趋势,并在 S5样地分别达最大值 7.50 kg·hm−2和 54.35 kg·hm−2,土壤 C 层的铅贮量在整个演替序列呈波动增长的趋势。
此外,生态系统铅的积累速率不仅能够量化整个演替序列对铅的积累过程,还能反演一定时间范围内铅的污染历史。本文根据贡嘎山东坡树木年轮(距离海螺沟冰川退缩区峨眉冷杉顶极群落直线距离约200 m)反演该地区百年来重金属污染历史(何咏梅等,2021),并结合海螺沟冰川退缩区原生演替序列120多年来生态系统铅的积累过程,进一步说明生态系统对铅污染的监测作用。从图7可以看出,1860—1920年间,树轮中铅的含量较低且变化较小,1920年以后树轮中的铅含量呈波动性增长,尤其是1905—1955年以及1970—1995年这两个时间范围内,树轮中铅的含量增长较为明显。从海螺沟冰川退缩区原生演替生态系统铅的积累速率来看,1890—1930年生态系统铅的积累速率变化平稳,1930年以后则呈现出波动性增长,尤其是在1958—1970年和1980—2000年这两个时间范围内生态系统对铅的积累较为明显,并且能很好的与树轮中铅含量的变化趋势相对应。这说明生态系统对铅的反应较为敏感,其植被和土壤对铅污染的响应能够反演近百余年时间尺度上铅的污染记录。
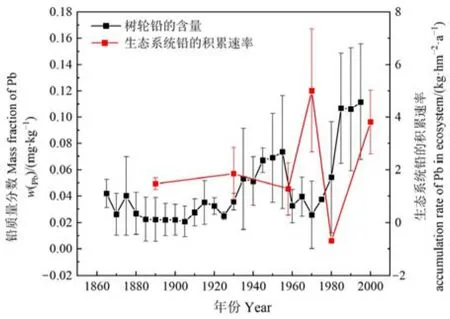
图7 生态系统铅的贮量与树轮铅质量分数Figure 7 Pb storage in ecosystem and mass fraction of Pb in tree rings
3 讨论
3.1 原生演替生态系统对铅的富集作用
不同演替阶段各优势乔木地上部分铅含量及其转运能力和富集程度均较低,而细根中铅的含量和富集程度明显高于其他部位。可见植物根部所吸收的铅绝大部分被存留在根系组织,只有很少一部分铅借助共质体被转移到地上部分(夏时雨等,1994)。这主要是由于土壤中的铅大部分以Pb(OH)2、PbCO3等难溶性盐及有机络合态存在,导致土壤中铅的移动性较差且植物有效性低(唐荣贵,2015)。此外,植物根系所吸收的铅在向地上部分转运的过程中还会受到内皮层组织中凯氏带屏障作用的影响,进一步阻挡铅的输送。而土壤中的铅能被根毛吸附并保留在细胞壁上从而促进细根对铅的吸收(Kabata-Pendias,2000;Uzu et al.,2010)。因此,铅经过根系吸收再向植物地上部位运移较为困难。虽然叶片能够吸附大气中的铅而导致其含量较高(陈英旭等,2000),但该植被演替序列各优势乔木叶片的铅含量并没有明显高于其他地上部分。因此,受土壤铅的存在形态以及植物根系不同部位和组织差异化分布的影响,导致该演替序列各优势乔木地下根系铅的含量明显高于地上部分。值得注意的是,峨眉冷杉枝铅的含量与细根铅的含量相当,且枝中铅的转运系数也明显高于峨眉冷杉及其他优势乔木不同部位的转运系数。这可能是由于针叶根系较强的有机酸分泌能力使得土壤 pH值下降从而增加土壤溶液中铅的溶解度(夏时雨等,1994)。此外,有机酸还可以促进铅从植物根系向地上部分转移(Tyler,1990),而导致峨眉冷杉枝的铅含量较高。林下植被中,地被层(苔藓)铅的平均含量高达 17.23 mg·kg−1,明显高于乔木、灌木各部位以及草本层的铅含量,且苔藓生物量对该演替序列生态系统活体植被总生物量的贡献不足 5%(杨丹丹等,2015),但其铅的积累贡献最大可达36%,表现出对铅的明显富集。在早期的研究中,已有不少学者利用苔藓生物监测大气有毒金属污染水平(Boutron et al.,1991;Stankovic et al.,2018),并指出苔藓具有较强的铅吸收能力(Wang et al.,2019)。而苔藓植物主要分布于高山高寒地区。因此,基于苔藓在高山生态系统中的分布范围较广且对铅的吸收能力较强,可能会造成这些地区对铅的富集。
表层土壤Oe层和Oa层的铅含量在整个演替序列的变化趋势一致,表现出随演替先增长后下降的趋势。从土壤O层铅的富集系数来看,其含量的变化并非受成土母质的影响,而可能受到来自凋落物归还、根系以及大气沉降等因素的影响(杨丹荔等,2020)。该演替序列中期形成以冬瓜杨为主的落叶阔叶林群落,其凋落量达整个演替阶段的最大值(杨丹丹等,2015),枝和叶等吸收的铅通过凋落物分解归还于土壤导致表层土壤铅含量增加;根系作为该演替序列优势种铅富集的主要部位,同样会导致土壤中的铅含量增加。此外,到 S6样地,土壤O层中铅的含量有所下降,这可能与该演替序列表层土壤pH值的变化有关。S1样地表层土壤的pH值高达8.54(杨丹荔等,2020),随着植被的演替尤其是 S6样地形成以峨眉冷杉为主的针叶林后,一方面针叶树种根系的外生菌根分泌有机酸的能力较强;另一方面针叶树种凋落物的酸性较低,这导致S6样地表层土壤的pH值仅为4.3,表现出明显的酸化。而土壤的pH值越低,土壤对铅的“固定”能力和吸持能力也越弱(王春苗等,2009)。因此,在酸性土壤条件下,铅的淋溶迁移能力较强,而滞留在土壤中的铅相对减少(Glover et al.,2002;潘胜强等,2014),这也导致该演替序列表层土壤铅含量在演替末期略有下降。
3.2 原生演替生态系统铅反演的污染历史
常用的元素地球化学记录的自然档案有冰岩芯、树木年轮、湖泊沉积物以及黄土剖面等。通过对比分析冰芯、湖泊沉积等对重金属的污染历史记录可知,在格陵兰(Boutron et al.,1991)和阿尔卑斯山 Monta Rosa地区(Schwikowski et al.,2004)的冰芯中发现在 20世纪 70年代铅的含量达到峰值;在加拿大Clair湖芯(Ndzangou et al.,2005)中也发现 1975年沉积物中的铅浓度明显增高。而在研究青藏高原地区重金属污染历史的结论中发现,帕米尔高原的慕士塔格冰芯(李真等,2006)中的铅浓度在1970—1990年间出现两个峰值,随后逐渐下降;在喜马拉雅山系中段的希夏邦马峰达索普冰芯(霍文冕等,1999)中发现 20世纪 70年代以后的铅浓度增长非常明显;在贡嘎山草海子湖泊沉积中铅的人为通量在 1950年以后显著增加,并在20世纪90年代中期达到峰值,随后逐渐减小(Bing et al.,2016);贡嘎山东坡峨眉冷杉树轮中铅的含量在1905—1955年以及1970—1995年呈上升趋势。由此可见,无论从全球还是青藏高原地区铅的污染历史记录来看,工业革命以来冰芯、湖泊沉积以及树轮所记录的铅含量总体呈波动增长的趋势并在1970年左右达到峰值,随后略有下降。
已有学者利用海螺沟冰川退缩区原生演替生态系统反演近百年来镉的污染历史(Wang et al.,2020a)。同样,该原生演替序列生态系统中的铅经历了从无到有的积累过程,其积累速率在冰川退缩1890—2000年间呈波动变化,尤其是在1958—1970年间积累较快,并在1970年达最大值,随后略有下降到 1980以后又有所回升。可见,该演替序列生态系统铅的积累过程能够很好的与青藏高原乃至全球冰芯、湖泊沉积以及树轮等对铅的污染历史记录相对应,反映同一时期铅污染的历史状况。贡嘎山地区的大气运输在冬季和春季主要受到西风带的影响,在夏季主要受到印度季风的影响(Fu et al.,2009)。通过 HYSPLIT后向轨迹模型计算出2021年海螺沟冰川退缩区的大气轨迹图(图8),可知该地区的大气污染物的输入可能主要来自于中亚和南亚地区。这些地区对 1958—1970年大气中铅沉降的上升均有影响,例如,印度在 20世纪50—80年代末,矿物燃料排放的废气位居世界第9(Pan et al.,2015);中亚5国作为前苏联主要的军事工业基地,在该时期美苏的两次大规模军备竞赛中可能造成污染物的大量排放(Golomb et al.,1997)。贡嘎山位于中国西南地区,已有研究表明该地区受到来自四川东北部和云南西北部等地区铅污染长距离传输的气团影响(Golomb et al.,1997),而中国直到 1970年以后大气污染防治才开始起步。此外,在过去120多年里,该演替序列生态系统铅的平均积累速率为 2.12 mg·m−2·a−1,但受输出通量的影响,大气平均积累速率通常要低于大气总的沉降通量。例如,中国北方大气铅总沉降通量(14.1 mg·m−2·a−1)比平均铅积累速率高 3 倍(Luo et al.,2015);而在欧洲和北美地区则比铅的积累速率高1—3倍(Kyllönen et al.,2009)。贡嘎山地区属于偏远地区,当地没有明显的工业污染排放和人类活动,生态系统中重金属多源于外来输入。若未来大气铅的沉积通量仍然升高,这将导致贡嘎山地区存在潜在的铅污染风险。
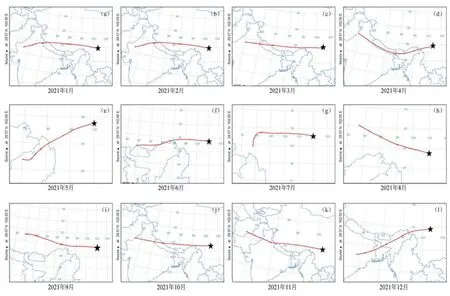
图8 海螺沟冰川退缩区大气轨迹图Figure 8 Atmospheric trajectory of Hailuogou Glacier Retreat area
4 结论
海螺沟冰川退缩区原生演替生态系统对铅的富集主要集中于土壤表层、优势乔木的细根以及地被层的苔藓中,且随冰川退缩时间的增加,生态系统对铅的贮存也随之增加并主要贮存于土壤之中。但值得注意的是,苔藓的生物量对总生物量的贡献不足5%,但其对植被铅的积累贡献最大可达36%,表现出明显的富集能力,这对高山高寒等地区的生态系统可能造成潜在的铅污染。
通过原生演替生态系统对铅的积累过程反演出近百年来铅污染在 1958—1970年和 1980—2000年这两个时间范围内生态系统积累较快,尤其是在1970年出现明显的峰值。这与青藏高原冰芯、贡嘎山东坡湖泊沉积和树轮中铅含量的变化趋势相对应,说明原生演替序列提供了铅在百年时间尺度的生物地球化学记录,也进一步证明铅污染通过远距离的大气传输已影响到相对偏远且洁净的地区。
