氮磷添加对高寒草地土壤养分和相关酶活性的影响
2022-02-19盛基峰李垚于美佳韩艳英叶彦辉
盛基峰 ,李垚, ,于美佳, ,韩艳英 , ,叶彦辉 ,
1.西藏农牧学院高原生态研究所,西藏 林芝 860000;2.西藏高原森林生态教育部重点实验室;西藏 林芝 860000;3.西藏高寒植被生态安全重点实验室,西藏 林芝 860000;4.西藏农牧学院资源与环境学院,西藏 林芝 860000
在陆地生态系统中,氮和磷是限制植物生长的两种主要元素(Harpole et al.,2011)。在过去的一个世纪里,由于化石燃料燃烧、农业施肥和粉尘或灰烬的产生等人类活动,全球生态系统的氮和磷输入大幅增加(Wang et al.,2015)。据预测,到2050年,全球大气氮沉降将增至 200 Tg(Moorhead et al.,2013)。过量的氮沉降会对氮循环和平衡产生负面影响,影响土壤养分状况、土壤微生物多样性和植被生产力(Song et al.,2020)。适量的氮添加可以改善土壤环境,使土壤肥力增强,方运霆等(2004)通过模拟大气氮沉降发现,大气中的氮沉降不仅能够显著提高土壤中无机氮的含量,而且伴随氮沉降时间的延长,土壤中无机氮含量也会呈现出显著增加的趋势。张进霞(2014)对紫花苜蓿(Medicago sativa)的NP添加实验有相同结论,养分添加显著提高土壤碱解氮含量。磷是自然界中最重要的元素之一,参与生物细胞膜的合成和酶的活化(Vance et al.,2003)。土壤中的磷很容易与土壤中的铝和铁的氧化物以及氢氧化物进行化学反应,强烈抑制了其从土壤中的释放,导致磷利用率低(Li et al.,2021)。以往的研究表明,土壤的氮、磷含量在自然界中处于一种动态平衡过程,大气氮沉降使土壤中氮元素含量增加,使影响植物生长发育的限制因素由氮元素转换为磷元素;反之,当土壤中磷含量较高时,土壤中就会出现氮限制(Berendse et al.,2001)。近些年来,通过对不同草地生态系统的研究发现,施肥处理对磷元素的影响结果并不一致。Widdig et al.(2020)报告称,在温带草原,氮和磷的添加对土壤全磷浓度没有显著影响,而其他研究报告称,氮的添加显著降低了温带草原的土壤全磷和有效磷(Gong et al.,2020)。
土壤酶作为土壤生物各种生命化学反应的催化物质,维持着土壤生态系统营养物质的正常循环和平衡(Yu et al.,2016)。土壤酶是土壤营养物质循环的主要驱动因子,其活力是衡量土壤肥力和评价土壤生态系统稳定性的重要指标(Yang et al.,2017)。一些研究表明,添加氮对细胞胞外酶活性具有中性影响,但P添加对胞外酶活性有负面影响(Jing et al.,2016)。Wu et al.(2022)对高寒草地养分添加的研究表明,氮和磷添加显著降低了葡萄糖苷酶催化效率。吴建波等(2021)对藏北高寒草原土壤氮添加发现,土壤pH对土壤酶不存在显著影响,且氮添加降低了碳相关水解酶活性,提高脲酶活性。
土壤有机碳是指土壤中各种正价态的含碳有机化合物,是土壤极其重要的组成部分,不仅与土壤肥力密切相关,而且对地球碳循环有巨大的影响(曹石榴,2019),既是温室气体“源”,也是其重要的“汇”。高寒草地是青藏高原主要分布广泛的草地类型之一,其有机碳储量约为3.02 Pg。受青藏高原恶劣的气候条件和营养物质(主要是氮和磷)限制,高寒草地对环境变化非常敏感,特别是全球变暖和氮磷的增加(Fayiah et al.,2020)。例如,气候变暖导致冰川急剧退缩和多年冻土快速融化,直接导致高寒草地退化和有机碳储量减少(Liu et al.,2018)。NP添加显著影响高寒草地碳循坏和有机碳储量,与添加N和NP相比,P添加土壤有机碳存储最低(Luo et al.,2020)。为研究高寒草地土壤有机碳固存的关键驱动因素及其影响因素,从而防止草地退化或维持高寒草地的稳定。本研究通过以藏东南色季拉山高寒草地土壤为研究对象,设置长期外源养分添加样地,分析土壤养分含量和酶活性对NP添加的响应,同时分析土壤碳、氮、磷与蔗糖酶、葡萄糖苷酶、脲酶之间的交互作用,旨在揭示高寒草地土壤养分和酶活性的内在关系,探究土壤碳氮磷元素在氮沉降背景下如何响应,为高寒草地的保护提供重要的理论依据。
1 材料与方法
1.1 研究区概况
西藏色季拉山位于念青唐古拉山与喜马拉雅山接合处的林芝县境内,属念青唐古拉山余脉,山体位于 93°12′—95°35′E,29°10′—30°15′N。山体走向主要为西北-东南,形成较大范围的东西坡面。本试验选定色季拉山西坡(29°38′8″N,94°37′26″E,平均坡度 21.4°)海拔 4400 m 高寒草地为试验样地,样地视野开阔,盖度较大。样地属湿润山地暖温带和半湿润山地温带气候,年平均气温为−0.73 ℃,年平均降水为1134.1 mm,降水主要集中在4—10月,无霜期180 d,平均相对湿度60%—80%,土壤以山地棕壤和酸性棕壤为主,草地主要植被为五叶双花委陵菜(Potentilla bifloravar.lahulensisTh.Wolf)、白心球花报春(Primula atrodentataW.W.Sm.)和蓝钟花(Cyananthus hookeriC.B.Clarke),占样地面积80%。施肥前土壤理化性质见表1。

表1 试验样地土壤原始状况Table 1 Original soil conditions of the test plot
1.2 试验设计
本试验样地所在的草地采用封闭管理,减少外界环境的干扰。本试验共有4个处理,分别为CK(对照组,自然环境条件)、N(硝酸铵NH4NO3,15 kg·hm−2·a−1)、P(过磷酸钙 Ca(H2PO4)2,75 kg·hm−2·a−1)、NP(NH4NO315 kg·hm−2·a−1N、Ca(H2PO4)275 kg·hm−2·a−1P)。样地内设置 3 个重复区组,每个区组内布设4个大小为3 m×3 m的样方,样方与样方之间设置5 m的缓冲带,区组与区组之间设置10 m的缓冲带。本试验研究区于2019年5月进行实地考察布设,于8月初植被生长旺盛期进行施肥处理,施肥时选择天气晴朗的早上,每次施加的水量大约相当于当地2.0 mm的自然降水量,CK添加等量的水。施肥采用人工背负式喷雾器喷洒的方式,将NH4NO3和Ca(H2PO4)2均匀喷洒在土壤表面。
1.3 土壤取样与测定
土壤样品在2020年8月施肥前(施肥1年后)进行收集,以便于测定施肥1年后土壤养分和酶活性变化情况。在每个样方内用直径5 cm的土钻采用五点取样法采集0—20 cm表层土壤,将同一样方内5个样品均匀混合为1个土壤样品(共12份鲜样)。利用2 mm的网筛去除土壤样品中较大的根和石头,冷藏保存带回实验室,将样品自然风干后测定土壤的理化性质,测量方法见表2(范珍珍等,2018;文旻等,2021;吴建波等,2021)。
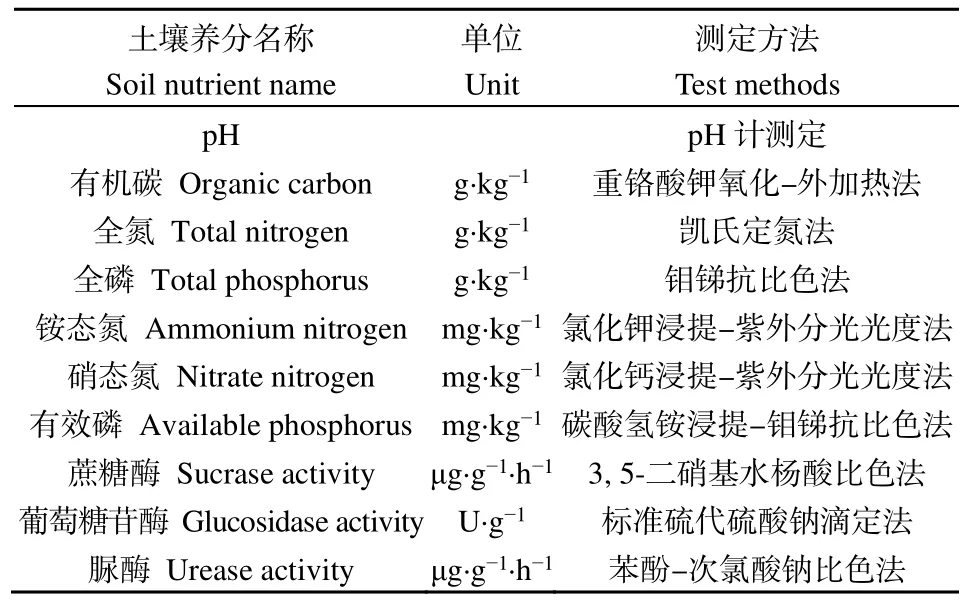
表2 土壤理化性质和酶活性测定方法Table 2 Methods for determination of soil physical and chemical properties and enzyme activity
1.4 数据统计与处理
应用Excel 2019、SPSS 25结合Sigmaplot 14.0和Origin 2022对获取的数据进行整理和统计分析。采用多因素方差分析检测N添加、P添加、NP混合添加对土壤养分和酶活性的影响。采用单因素方差分析(One-way ANOVA)和最小显著差异法分别进行各试验处理间差异显著性检验及多重比较。应用Origin绘图。
2 结果与分析
2.1 土壤pH
本试验中P和NP添加处理均显著降低了高寒草地土壤的pH值(P<0.05)(图1),各添加均使土壤酸化程度加深,其中P添加处理最低(4.8)。
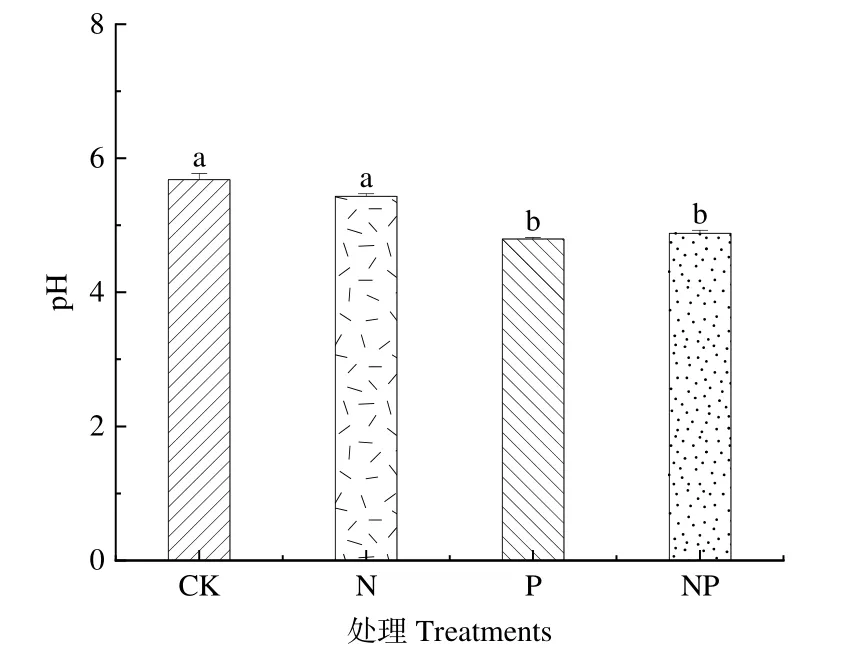
图1 氮磷添加对土壤pH的影响Figure 1 Effects of nitrogen and phosphorus addition on soil pH
2.2 土壤养分
图2a显示,NP添加与对照相比显著促进了土壤有机碳含量(P<0.05),具体表现为在NP处理下有机碳含量最高,提高了63.50%,N、P单独处理次之,分别提高了50.86%、38.33%,相比于单一添加,只有在氮磷混合添加下对土壤有机碳含量影响最大。图2b—d显示,N、P添加与对照相比显著增加了土壤全氮含量(P<0.05),具体表现为在N添加下全氮最高,提高了59.14%;P、NP添加次之,分别提高了40.86%、11.11%。氮磷共施减弱了氮肥的影响,使N处理下全氮含量显著高于NP添加。施肥处理下,土壤铵态氮和硝态氮含量显著增加(P<0.05)。在N添加下土壤铵态氮含量与硝态氮含量均达到最高,相比于 CK分别增加了47.58%、333.85%。不同处理下铵态氮含量的整体表现为 N>NP>P>CK,硝态氮含量整体表现为N>P>NP>CK。图2e、f显示,不同处理下土壤全磷含量分别为 0.71、0.74、0.69、0.84 g·kg−1。NP 添加下显著促进全磷含量(P<0.05),提升幅度为19.41%;在N添加下土壤有效磷含量最高,增加了59.68%;P、NP添加下分别增加了 25.25%和38.85%;有效磷含量整体表现为N>NP>P>CK。
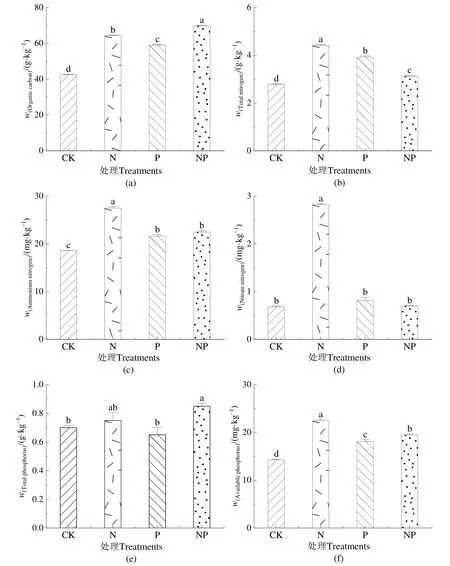
图2 氮磷添加对土壤养分含量的影响Figure 2 Effects of nitrogen and phosphorus addition on soil nutrient content
2.3 土壤酶活性
图3a、b显示,不同施肥处理均显著促进蔗糖酶和葡萄糖苷酶活性(P<0.05),与CK相比,P添加下土壤碳相关酶增幅较小,分别增加了24.96%、20.06%。蔗糖酶活性在 NP添加下最高,增加了92.63%,N添加下增加了81.92%。葡萄糖苷酶活性在N添加下最高,增加了71.94%,NP添加下增加了53.08%。图3c显示,在施肥处理下土壤脲酶活性均显著高于CK处理(P<0.05),且P添加下增幅最大,增加了140.73%,N和NP添加下分别增加了58.13%、115.31%。
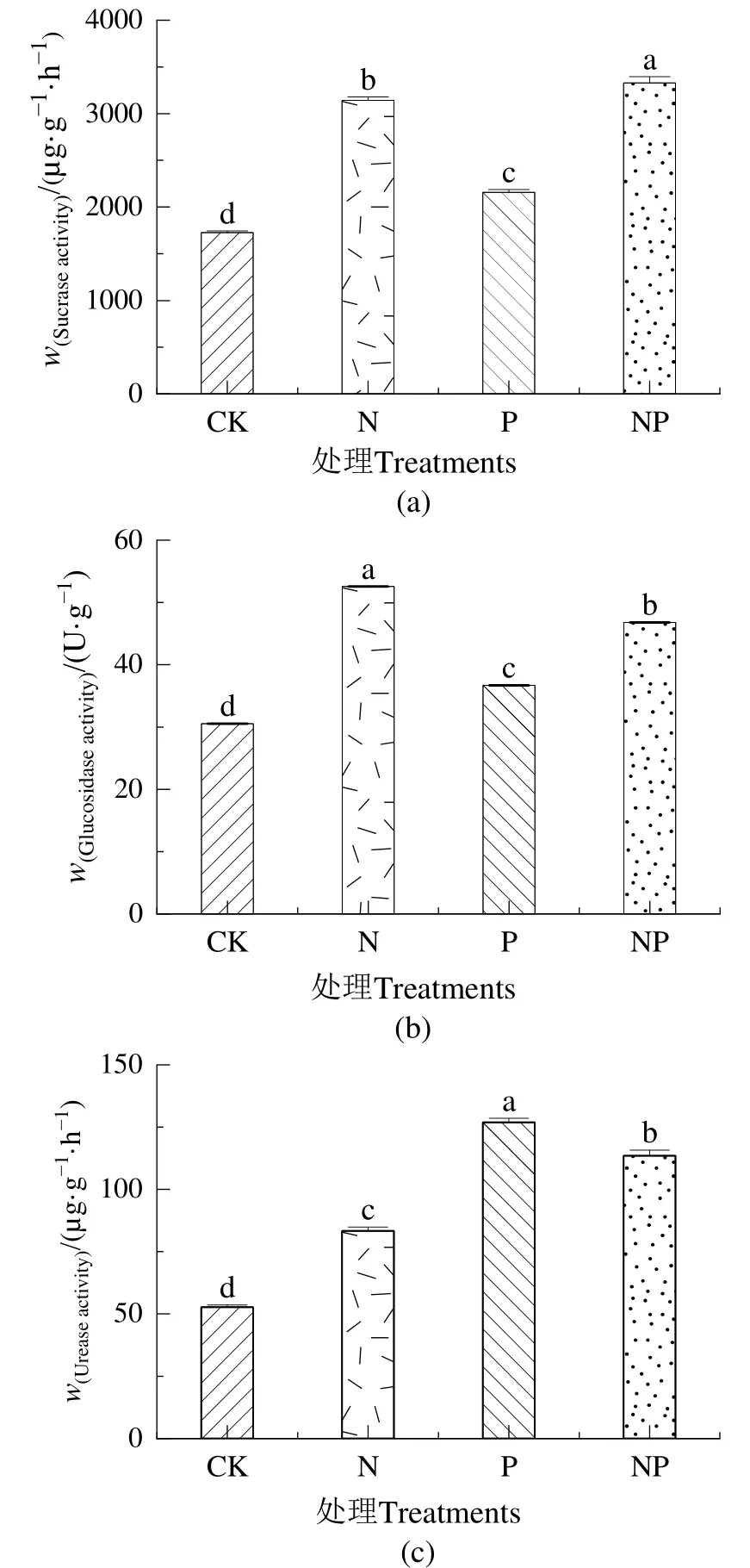
图3 氮磷添加对土壤蔗糖酶、葡萄糖苷酶和脲酶活性的影响Figure 3 Effects of nitrogen and phosphorus addition on activity of soil sucrase, glucosidase and urease
2.4 土壤养分与酶活性的相关性分析
相关性分析表明(表3),pH与全氮、有机碳与全磷和有效磷、全氮与铵态氮、硝态氮与葡萄糖苷酶呈现显著正相关(P<0.05)。pH与铵态氮、硝态氮、有效磷、葡萄糖苷酶,有机碳与蔗糖酶和葡萄糖苷酶,全氮与硝态氮,铵态氮与硝态氮、有效磷和葡萄糖苷酶,硝态氮与有效磷,葡萄糖苷酶与有效磷、蔗糖酶,均呈现极显著正相关(P<0.01)。脲酶与pH、有机碳、铵态氮、硝态氮、有效磷、葡萄糖苷酶呈现极显著负相关(P<0.01)。
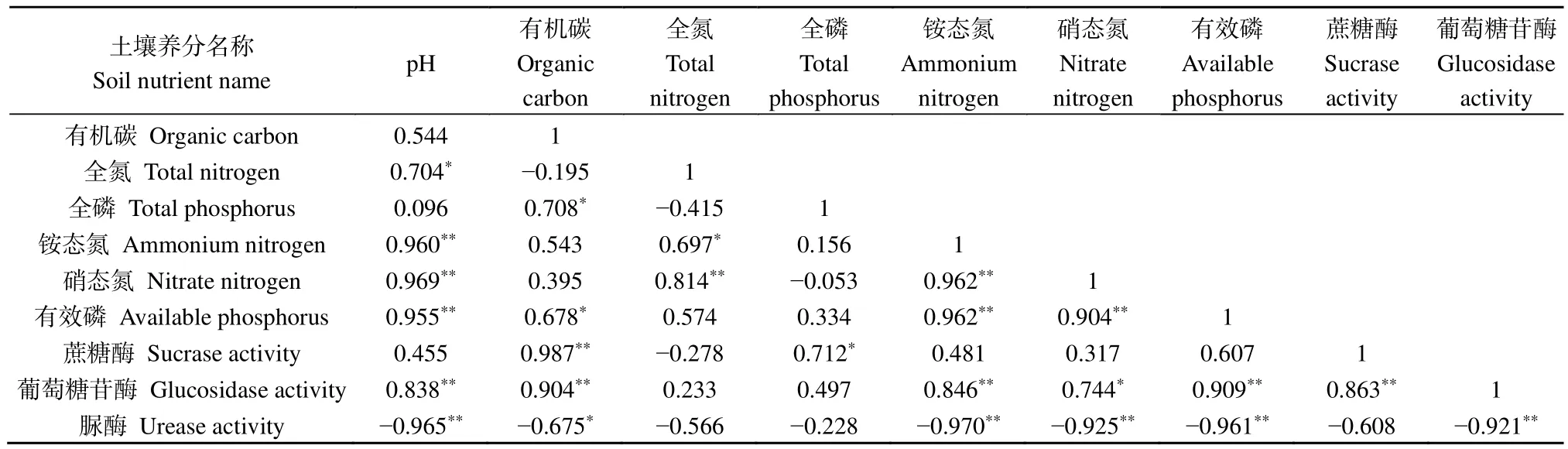
表3 不同处理后土壤性质Pearson相关性分析Table 3 Pearson correlation analysis of soil properties after different treatments
3 讨论
3.1 氮磷添加对土壤pH和养分的影响
土壤酸碱度是影响土壤肥力的因素之一,与土壤中的大部分化学反应以及各种生物过程均有极大的关联,其值大小主要由土壤水溶液中氢氧根离子与氢离子的浓度决定(Li et al.,2021)。本研究中施肥处理均降低了高寒草地土壤的pH值(图1),与大多数实验结果一致(苏洁琼等,2014;刘红梅等,2018),可能是由于铵态氮通过细菌硝化作用向土壤中释放更多的氢离子,从而降低了土壤的pH值。
有机碳是植物生长发育的必需元素,是植物有机质的重要组分之一。研究表明土壤中施加有机物可以给微生物供给足够的底物,加速有机碳的矿化,释放更多的活性碳组分促进土壤碳循环(赵晓琛等,2016)。本研究中,施肥处理均显著增加土壤有机碳含量(图2a),且NP合施处理有机碳含量最高,这与翟珈莹(2020)在青藏高原0—20 cm土层研究结果相一致,可能由于氮、磷含量处于一种动态平衡过程,氮添加下土壤氮含量增加,打破了氮磷平衡,使磷限制更加明显,说明NP合施可以缓解由于单独施氮造成的磷素限制。史晓鹏(2018)研究了 N(70 kg·hm−2·a−1)、P(17、34 kg·hm−2·a−1)和 NP 混合添加处理对苜蓿的影响,发现施肥均增加了有机碳含量。然而,Luo et al.(2019)和Li et al.(2020)研究均表明,氮磷富集加速青藏高原高寒草地土壤有机碳分解,降低了土壤总有机碳库。
土壤全氮包括有机氮和无机氮,是土壤中各种形态氮元素含量总和(王常慧等,2004)。硝态氮与铵态氮在植物生长和发育过程中以离子的形式直接被植物所吸收利用,其在土壤中含量将直接影响植物群落生产力的变化趋势(Liu et al.,2022)。叶彦辉等(2017)在高寒草地进行的氮添加试验与本研究结果一致,一定量的氮添加可以提高土壤中全氮含量。相关性分析表明,pH与全氮呈现显著正相关关系,与铵态氮和硝态氮呈现极显著正相关关系,养分添加后pH的改变是氮元素含量变化的原因之一。本研究中不同施肥处理下全氮、铵态氮和硝态氮变化趋势相似(图2b—d),均在N添加下增幅最大,氮输入对土壤环境改变较大。此外,在N添加下硝态氮变化较铵态氮变化大,Fu et al.(2017)对高寒草地综合分析得出一致结论,其原因有两种可能,一是高寒植物对铵态氮的利用率要高于硝态氮,二是不同植物类型对铵态氮和硝态氮养分吸收不同,当铵态氮含量达到植物需求后多余的量通过硝化作用转化为硝态氮。本研究发现,氮磷混施下土壤全氮和硝态氮含量低于氮和磷添加处理,这可能是氮磷混施下植物生长加快,消耗了土壤中无机氮(苏渝钦等,2016)。
本研究发现,土壤全磷含量仅在NP添加下显著增加(图2e),高寒草地氮元素缺乏且氮磷比失衡,单独的N、P添加对土壤全磷无显著影响,与刘姝萱等(2022)对荒漠草原和Widdig et al.(2020)在温带草原研究结果一致。有效磷是土壤中可被植物吸收的磷组分,本研究中有效磷变化显著,各处理均呈现增加趋势(图2f),原因可能是养分输入提高了微生物和磷相关酶的活性,磷相关分解酶活性的提高增强了微生物对土壤有机质的分解,加快了土壤中磷元素的转换过程,从而提高有效磷含量(魏金明等,2011)。Hu et al.(2022)通过Mate分析发现环境因素中 pH、海拔、年平均温度和年平均降水量显著影响有效磷含量,因此,环境因素是不同地区磷元素研究结果不同的重要原因。对鄱阳湖苔草研究表明,N添加对湿地土壤全磷及有效磷含量影响均不明显,研究区的土壤周期性淹水使土壤有机质分解相对较缓,土壤磷含量相对较高(文旻等,2021)。本研究中N、P添加下全磷含量变化指示了高寒草地养分限制因素,有效磷是土壤中可被植物吸收的磷组分,有效磷含量变化表明高海拔下进行施肥处理有利于地上植被生长。
3.2 氮磷添加对土壤酶活性的影响
土壤蔗糖酶和葡萄糖苷酶均是主要的碳获得酶,是土壤碳循环过程的主要指标。蔗糖酶能通过水解形成果糖和葡萄糖,葡萄糖苷酶可以通过水解作用生成葡萄糖(Zi et al.,2022)。本研究各处理下蔗糖酶和葡萄糖苷酶变化趋势相同,均在N和NP添加下促进效果最大,P添加对土壤碳获取酶影响较小。相关性分析结果表明(表3),蔗糖酶和葡萄糖苷酶均与有机碳呈现极显著正相关关系,这可能是由于养分增加,植物根系生长,导致土壤碳有效性不足,增加了微生物对碳的需求,改变了微生物对养分获取策略,增加了蔗糖酶活性和葡萄糖苷酶活性,从而提高酶活性,然而,由于施肥量不同导致促进效果不同(Liu et al.,2022;赵晓琛等,2016)。碳获取酶活性研究有不同结论,孙亚男等(2016)在高寒草地研究结果与本研究不同,4年NP添加下各处理对0—10 cm和10—20 cm土层蔗糖酶活性均没有显著影响,这与施肥时引入碳元素有关。
本研究发现,施肥处理显著促进脲酶活性,这与宋学贵等(2009)和涂利华等(2009)在不同森林生态系统中的研究结论相类似。N和NP添加对脲酶活性影响较大,N添加下脲酶活性增加,而在NP共同添加下其活性则进一步显著增加。从相关性分析结果可知,脲酶活性与pH、铵态氮、硝态氮、葡萄糖苷酶呈极显著负相关。脲酶主要来自植物根系的分泌物,而氮添加促进了植株根系的生长,导致根系分泌物增加,增强脲酶活性。本研究中施肥处理中P添加下脲酶活性最高,表明P添加使土壤铵态氮含量和葡萄糖苷酶活性降低导致有机氮的矿化速率升高,从而使土壤微生物群落对土壤脲酶的分泌增加,其活性也随之增高,与Zi et al.(2022)在 P添加下脲酶活性增高结果一致。冯慧芳等(2022)对 5种不同密度大叶相思(Acacia auriculiformisA.Cunn.ex Benth.)研究结果有不同结论,N和NP添加对脲酶活性均有显著抑制现象,P添加对高密度林分脲酶活性有显著促进作用,这可能与N添加量植被类型、N添加时间长短不同有关。
土壤酶主要来自于土壤微生物的释放,还包括植物根系分泌物以及土壤动植物残体腐解过程的释放(Yang et al.,2017)。本研究中,各施肥处理对蔗糖酶、葡萄糖苷酶和脲酶活性均表现出显著促进作用(图3a—c),这与Xiao et al.(2021)的施氮试验结果一致,施氮显著提高转化酶活性,NP添加通过改变表层土壤养分条件,改变植物对养分的吸收利用,影响高寒草地土壤有机层细菌的生长活力和营养有效性,使植物、微生物群落结构及数量发生变化,进而不同程度提高有关碳氮获取酶的含量。
4 结论
本研究中施肥处理相比于对照处理显著改变了高寒草地土壤养分和酶活性条件,降低了土壤pH。氮和氮相关(NP添加)添加对碳含量有显著的促进作用,主要表现为N添加后显著促进土壤有机碳含量,并增加蔗糖酶和葡萄糖苷酶活性,水解更多大分子含碳有机物供植物吸收利用。N添加和P添加显著促进土壤氮含量,N添加显著促进土壤氮元素含量,增幅较大,P添加促进土壤脲酶活性,水解含氮物质,给植物提供养分。然而,NP添加下氮元素并没有促进迹象,具体表现为氮素含量较单独的 N和P添加下显著降低。本研究未对磷相关酶和地上地下生物量进行研究,今后应进一步研究。
