施氮和干旱对杉木和青冈幼苗生物量及根系形态的影响
2022-02-19高歌葛晓改周君刚周本智李正才杨南
高歌 ,葛晓改 ,周君刚,周本智 ,李正才 ,杨南
1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2.南京林业大学,江苏 南京 210037;3.国家林业和草原局钱江源森林生态系统定位观测研究站,浙江 杭州 311400;4.杭州市富阳区农业农村局,浙江 杭州 311400;5.云南西双版纳国家级自然保护区管护局,云南 景洪 666100
受全球气候变化的影响,未来局部地区的水分分配状况将发生改变(Sparrius et al.,2012)。在这种背景下,中国亚热带地区降水的季节性波动也愈加明显,干旱风险加大必将对植物的生长带来更严峻的考验(Kudoyarova et al.,2015;孙百生等,2018)。干旱的发生会导致土壤水分有效性降低,使植物体吸水量与耗水量失衡,进而影响植物的正常生理需求(高小敏等,2021;何佩云等,2022)。氮素是土壤养分的重要元素,为植物生长提供必要的养分来源(吕豪豪等,2016)。水分是土壤养分的有效载体,植物生长主要受到水分条件的限制,通过施肥等措施调节植物水分状况是减轻干旱伤害的有效途径之一(Dichio et al.,2007;丁红等,2015)。有研究发现适量的氮素添加能促进植株各器官干物质积累和生物量的分配,达到改善植物水肥关系和强化植株吸收氮素的效果(Zhou et al.,2014)。
根部是获取土壤水分的关键部位,其中根系形态特征影响着植物对营养资源的吸收,施氮通过影响植物根系生长发育进而改变其水分利用特征,(尹丽等,2012;郭京衡等,2014;杨腾等,2014;赵松实等,2021)。
由于水分与氮素之间密切而又复杂的关系,近年来,越来越多的研究开始关注水-氮协同作用(崔婉莹等,2019)。外界环境变化对植物的影响表现出复杂性及不确定性,土壤水分和氮素之间可能会发生叠加、促进或拮抗效应,一方面增加土壤水分能促进养分溶解和迁移进而提高养分有效性;另一方面降水过多也会引起土壤养分的淋溶和反硝化作用,导致养分流失,而这取决于具体的生境及树种类型(Wang et al.,2005;Knapp et al.,2008;Egbert et al.,2009;Schuster et al.,2016)。在应对外界的不利环境时,不同植物亦能通过调整各器官生物量分配策略吸收更多的水分和营养元素以维持自身生存与生长发育(肖冬梅等,2004;尹丽等,2011)。
杉木(Cunninghamia lanceolata)是重要的速生丰产用材树种,也是目前中国人工林中面积最大的造林树种(Chen et al.,2013;Yu et al.,2015)。青冈(Cyclobalanopsis glauca)作为中亚热带常绿阔叶林带分布最广泛的树种之一,是优良的乡土阔叶用材树种(常杰等,1999)。在水分匮乏的条件下,植物会表现出不同的种间差异适应性,杨振亚等(2018)研究发现干旱胁迫下杉木幼苗地上部和根系生长受到显著抑制,吴丽君等(2014)表明不同干旱程度对青冈幼苗生长和生物量等指标产生了明显影响。水分和氮素是两树种生长过程中必不可少的环境因子(Yin et al.,2012)。以往关于亚热带地区树种的研究多集中于水分和氮素单一因素作用或耦合作用下对植物的生长生理特性的研究,干旱下不同树种地上和地下部生长对氮素响应的内容涉及较少,鉴于此,本文基于亚热带针阔叶两树种生物量的研究,结合地下部根系形态,探讨施加氮肥对生长时期遭受干旱胁迫的幼苗生物量累积分配和根系形态特征的影响,为干旱下两树种幼苗期的水肥管理提供理论依据。
1 材料和方法
1.1 试验地概况
试验在浙江省杭州市富阳区中国林业科学研究院亚热带林业研究所温室大棚(119°57′0″E,29°28′48″N)中开展,该区域海拔 130 m,属亚热带典型季风气候,年均气温 17.5 ℃,平均相对湿度70.9%,无霜期307 d,年日照时数为1663.2 h,年均降雨量为1457.7 mm,集中于4—9月。
1.2 试验材料与试验设计
供试植物幼苗包括针叶树种杉木和阔叶树种青冈。选取苗高和基径相对一致的1年生青冈[苗高(29.4±0.95) cm 和基径 (5.82±0.39) mm]和杉木[平均苗高 (27.33±1.07) cm 和基径 (6.44±0.56) mm]幼苗于2020年3月定植于25 cm×30 cm(容量约10 L)规格的盆栽塑料容器中,每盆装当地过10目筛的黄壤土约5 kg,土壤基本理化性质如下:有机碳18.70 g·kg−1,土壤全氮 0.86 g·kg−1,土壤全磷 0.26 g·kg−1,土壤全钾 11.20 g·kg−1,土壤水解氮 85.13 mg·kg−1,土壤有效磷 4.15 mg·kg−1,土壤速效钾 65.73 mg·kg−1,pH值4.91。为避免降雨对土壤水分含量造成影响,试验在隔离雨水,透光且通风透气的大棚内进行,除试验因素(水分、养分)外,其他条件尽可能保持一致,以保证整个试验具有可比性。
试验采用施氮(N0和N1)和水分(CK、MD和 SD)双因素完全随机区组设计,杉木和青冈两树种分别设置6个处理。缓苗期结束后,2020年6月对两树种幼苗进行氮肥添加试验:设未施氮(N0,0 mg·kg−1,以 N 计,下同)和施氮(N1,100 mg·kg−1)2个氮处理水平(黄彩变等,2021),施氮组(N1)以尿素[CO(NH2)2]为氮源,将18 g农用尿素(总氮≥46.4%)溶于 1 L 水中,施肥量为 20 mL·plant−1,每隔半个月施1次肥,共施3次;未施氮组(N0)每盆施加等量去离子水,以避免处理间因额外添加水分造成的影响。氮处理结束后于8月中旬展开水分控制试验:用环刀法测出土壤最大田间持水量为15.7%,设置80%—85% FC作为水分对照组(CK)、50%—55% FC为中度干旱(MD)、30%—35% FC为重度干旱(SD),即确保土壤含水量分别维持在12.6%—13.4%、7.9%—8.6%、4.7%—5.5%(全文选等,2017;赵松实等,2021)。运用 ZL6土壤水分监测系统(Meter,美国)对土壤含水量进行远程监测,同时每天16:00—18:00采用土盆质量法补充当天消耗掉的水量,确保干旱期间各处理组的土壤含水量基本维持在设定范围内(吕豪豪等,2016)。干旱处理60 d后,随机选取杉木和青冈各处理6株(作为6个重复)于当年11月生长季末期收获采样。
1.3 样品采集和测定
取样时用剪刀将植物地上部分和根系分开,抖落根系附着的土壤,将土壤过筛以获取残余根系,把样品标记后标封存在冰盒中带回实验室进行处理。
根系形态测定:用去离子水将根系清洗干净,擦干后用植物根系分析系统(Regent Instruments Inc.,WinRhizo,Canada)对新鲜根进行根系形态扫描,扫描后设置根系直径分级参数并通过根系分析软件(WinRhizo Pro 2016a)对图片进行分析,测定根系长度、根体积、根表面积、根尖数等指标。
生物量测定:根系扫描后,将根、茎、叶分别装入信封中,置于烘箱中105 ℃杀青30 min,然后在 65 ℃下烘干至恒质量,用电子天平(精度为0.001)称量干质量并计算以下指标:

其中:
RRB——根生物量比;
RLB——叶生物量比;
RSB——茎生物量比;
RSL——茎叶比;
RRT——根冠比;
LSR——比根长;
mroot——根生物量;
mleaf——叶生物量;
mstem——茎生物量;
maboveground——地上生物量;
mtotal——总生物量;
l——根长。
1.4 数据处理
数据分析应用SPSS 25.0软件,采用双因素(水分和氮素)方差分析(Two-way ANOVA)检验水分和施氮及交互作用对两树种生物量积累和物质分配影响。单因素方差分析(One-way ANOVA)检验不同水分和氮处理下杉木和青冈树种生物量积累、分配及根系形态的差异,用 Duncan法进行多重比较(α=0.05)。图表制作在Sigmaplot 12.5软件完成。
2 结果与分析
2.1 施氮对干旱下杉木和青冈幼苗各器官生物量的影响
表1双因素方差分析结果显示,氮素添加仅对杉木总生物量无显著影响(P>0.05),水分变化对各器官生物量的影响均达到极显著水平,并且水氮交互作用对杉木茎干质量、地上生物量和总生物量影响极显著(P<0.01);表2表明,青冈根干质量在水分或氮素单一因素作用下未受到显著影响(P>0.05),但两者交互效应对青冈各器官及整体生物量影响极显著(P<0.01)。
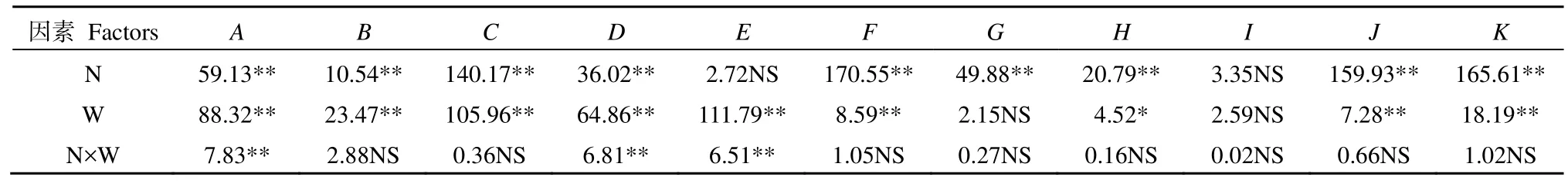
表1 不同水氮条件下杉木幼苗生物量累积及其分配的双因素方差分析(F值)Table 1 Two-way ANOVA results of effects of water, N and their interaction on the biomass accumulation and distribution of C. lanceolata seedlings (F)

表2 不同水氮条件下青冈幼苗生物量累积及其分配的双因素方差分析(F值)Table 2 Two-way ANOVA results of effects of water, N and their interaction on the biomass accumulation and distribution of C. glauca seedlings (F)
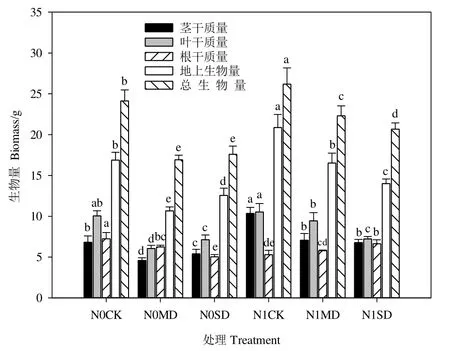
如图1所示,氮素添加使得相同干旱水平下杉木根干质量显著减少(P<0.05),但同一水分条件下施氮组杉木茎干质量和叶干质量均增加,其中茎干质量变化显著(P<0.05)。与未施氮组相比,氮素施加后中度和重度干旱下杉木地上生物量增加,总生物量则呈现减少的趋势。由图2可知,施氮增加了相同干旱梯度下青冈茎干质量、叶干质量及地上和总生物量,且中度干旱下的促进效应均显著(P<0.05),根干质量的变化为N0MD>N1MD,N0SD 图1 施氮对不同水分条件下杉木幼苗生物量的影响Figure 1 Effects of nitrogen on biomass of C.lanceolata seedlings under different water treatments 图2 施氮对不同水分条件下青冈幼苗生物量的影响Figure 2 Effects of nitrogen on biomass of C.glauca seedlings under different water treatments 由表1可知,杉木根生物量比、根冠比和比根长分别对氮素添加、水分变化的响应极显著(P<0.01),茎生物量比仅在氮素作用下受到极显著影响(P<0.01),氮素和水分及交互作用对茎叶比的影响均不显著,交互效应对各器官分配比例及比根长均无显著影响(P>0.05)。表2显示,对青冈而言,除氮素仅对叶生物量比无显著影响之外,氮素、水分及交互作用对其他指标的效应均显著(P<0.05)。 由表3可知,氮素添加使得同一干旱水平下杉木茎生物量比、叶生物量比及茎叶比增加,根生物量比、比根长和根冠比减少,其中,相同干旱处理下根生物量比、茎生物量比、根冠比及比根长差异显著(P<0.05),并且氮素对干旱组比根长的抑制效应在中度干旱下较强(39.8%),对根生物量比及根冠比的抑制效应则在重度干旱组表现更强,分别为30.0%和38.1%。中度和重度干旱条件下,青冈物质分配格局有所不同,施氮后N1MD组根生物量比和根冠比较N0MD组分别显著减少了29.7%和40.7%(P<0.05),茎生物量比、叶生物量比及比根长均增加。当水分含量降低至重度干旱时,根生物量比、根冠比和茎叶比的变化为N1SD比N0SD分别显著增加了10.3%、17.5%和23.7%(P<0.05),而N1SD组叶生物量比和比根长较N0SD组均显著减少(表4)。 表3 施氮对不同水分处理下杉木幼苗生物量分配的影响Table 3 Effects of nitrogen on biomass distribution of C.lanceolata seedlings under different water treatments 2.3.1 施氮对干旱下杉木和青冈各径级根长的影响 图 3a表明,施氮显著抑制了干旱下杉木细根及粗根根长,当氮素作用于径级小于2 mm的细根时,中度干旱下抑制效果强于重度干旱处理组,但对于杉木粗根(>2 mm)根长,则表现为重度干旱下的抑制作用更强。施氮对青冈各径级根长的影响为:正常水分下,施氮后各径级根长皆被抑制,当径级小于1.5 mm和大于2 mm时,施氮抑制了中度干旱下根长的伸长,但重度干旱时施氮表现为促进效应,且氮素对青冈细根(<2 mm)根长的作用效果较强(图4a)。 图3 施氮对不同水分条件下杉木幼苗根系形态的影响Figure 3 Effects of nitrogen on root morphology of C. lanceolata seedings under different water treatments 2.3.2 施氮对干旱下杉木和青冈各径级根表面积的影响 施氮后,同一干旱水平下杉木各径级根表面积显著减小(P<0.05)。中度干旱下,施氮对细根(<2 mm)根表面积的影响比粗根大,重度干旱时则与之相反。总体上,氮素对中度干旱下杉木根表面积的作用更强烈(图 3b)。就青冈而言,施氮抑制了正常水分下各径级根表面积(P<0.05)。与中度干旱下N0组相比,N1MD组粗根(>2 mm)的根表面积减小,水分含量继续下降后,重度干旱下N1SD组粗根根表面积比 N0SD组显著增加了 20%。由图 4b可知,氮素对重度干旱下青冈根表面积影响较大。 2.3.3 施氮对干旱下杉木和青冈各径级根体积的影响 从杉木各径级根体积变化看,同一干旱水平下N1MD及N1SD组分别较N0MD和N0SD组显著降低(P<0.05),且在中度干旱时的下降幅度更大,从图3c可知,氮素作用下,干旱组杉木细根(<2 mm)根体积比粗根更敏感。图 4c表明,施氮后中度干旱下青冈细根的根体积受到了不同程度的抑制,其中在1—2 mm径级差异显著。氮素对重度干旱下青冈根体积的影响比中度干旱大,相较于N0SD组,施氮使得N1SD组青冈根体积总体上呈增加趋势,且粗根根体积比未施氮组增加了53%(P<0.05)。 图4 施氮对不同水分条件下青冈幼苗根系形态的影响Figure 4 Effects of nitrogen on root morphology of C. glauca seedings under different water treatments 2.3.4 施氮对干旱下杉木和青冈各径级根尖数的影响 氮素对中度和重度干旱下杉木各径级根尖数的影响不同,并且在中度干旱下的作用效果更强。施氮促进了N1MD组杉木根尖数的增加,其中粗根(>2 mm)增长率最大,当遭受重度干旱时,在氮素作用下各径级根尖数均被抑制(图3d)。整体上看,施氮使得N1MD组青冈根尖数增加,且氮素对中度干旱下粗根根尖数的影响比重度干旱大,当径级小于1.5 mm时,N1SD组根尖数较N0SD组显著增加(P<0.05);当径级大于1.5 mm时,N1SD组根尖数比N0SD组减少(图4d)。 氮素和水分作为幼苗早期生长的限制性因素,两者对植物的影响具有耦合效应。王海艺等(2006)对洋白蜡(Fraxinus pennsylvanica)的研究显示氮肥对植物生物量的作用受水分因子的影响,不同的水肥配合下生物量的积累有所不同(Huang et al.,2009;周国逸等,2001)。本研究发现,在杉木和青冈幼苗生长过程中,水分和氮素的交互作用显著影响着干物质的累积,且氮素对不同干旱程度下两树种生物量的影响存在差异。施氮后土壤中可利用氮含量增多促使地上生物量积累,植物通常将更多的资源分配给繁殖器官和地上部分,与N0组相比,同一干旱水平下N1组杉木和青冈的地上生物量升高,植株通过调整光合器官的生物量储存来提高同化速率并促进地上部生长,同时地上生物量积累还起到了一定的遮阴效果,有利于减少土壤水分散失(Yang et al.,2011;Mao et al.,2012;吴茜等,2011;李茂等,2020)。当土壤水分含量较低时,根系的吸收面积和吸收能力随之降低,植物对氮素的吸收也相对较低(董雯怡等,2010)。施氮后杉木根系及整体干物质积累均被抑制,祁瑜等(2011)同样发现施氮对地上部分的生物量有促进作用,但对根系生物量积累不利,这可能是当外界资源不足时,植株将氮素首先运输到叶片以加强营养器官对限制性资源的获取利用(Johansson,2000)。基于不同植物对氮素和水分的需求和敏感性差异(Johansson et al.,2012),与杉木根生物量变化不同的是,随着水分减少,重度干旱下N1组青冈根系生物量较N0组显著增加,施氮使干旱下原来对植物生长的“无效”水分变得“有效”,从而使植物能吸收利用更多的水分,这表明氮素的施加促使青冈根生物量累积来提高自身对土壤“有效”水的利用效率(Aujla et al.,2007)。 外界资源受到限制时,植物通常会调整自身的生长方式和资源分配来适应环境变化(Wu et al.,2008)。不同类型植物的生物量分配格局对水分和氮素添加的响应也不尽相同(祁瑜等,2011)。这一观点在本研究中也得到了证实,两树种双因素方差分析结果表明,水氮交互效应对杉木各器官分配比例及比根长均无显著影响,但对青冈的影响则表现为极显著。施氮改变了干旱下青冈和杉木的物质分配格局,生物量分配格局的改变会对植物获取和吸收光能、养分及水分等功能产生影响(Huang et al.,2009)。其中,根冠比和比根长作为衡量植株生长状态及响应土壤水分和养分环境的重要指标,前者反映了生物量在地上和地下的分配情况,后者通过指示根系形态功能来综合表现植物吸收资源和适应生态的能力(Bauhus et al.,2000;吴茜等,2011)。干旱下两种植物的生物量分配格局对氮肥的响应也表现出不同的生态策略,施氮前后杉木地上生物量及根生物量均表现为随干旱程度加重而减少,说明杉木地上部生长及地下根系生物量积累与分配受到不同干旱程度的抑制,氮肥的施加在一定程度上促进正常水分组和干旱组杉木地上生物量积累,但表3表明,在氮素作用下未能促使杉木幼苗通过增加比根长及根冠比以缓解干旱胁迫的伤害,这可能是由于杉木自身对生长环境中的温度、水分及养分条件的要求较苛刻(杨振亚等,2018)。氮添加后重度干旱组青冈根系生物量及根冠比较未施氮组显著增加,这与常宏等(2019)验证施氮减水后大叶青冈(Cyclobalanopsis jensenniana)根冠比增加的结果相似,可见,在重度干旱胁迫下,施氮通过促进青冈幼苗根冠比增大,进而加大植物自身对有限水分及养分的吸收利用(徐炳成等,2003)。 根部是植物获取土壤水分的关键部位,其中根表面积、根长度和根直径等根系形态决定植物在吸收土壤资源和养分方面的竞争能力(Muhammad et al.,2017)。本研究中,水分和氮素影响着杉木和青冈幼苗的根系形态特征,这与孙誉育等(2015)及白亚梅等(2020)分别对红桦(Betual albo-sinensis)幼苗和红砂(Reaumuria soongorica)幼苗根系的研究结果一致。正常水分下氮素对于杉木和青冈根系形态产生负效应,这可能归因于两树种幼苗生长初期对土壤养分需求较小,土壤自身含有的养分已满足幼树早期的生长需要(许文斌等,2021)。施氮对重度干旱(N1SD)组杉木各径级根长、根表面积、根体积表现为负效应,这与N1CK组根系形态的变化一致,原因可能是土壤水分匮乏不是限制杉木幼苗吸收氮素营养的主要因素,杉木对N素的吸收利用还会受到环境中土壤N素浓度、N素形态和温湿度等的影响(闫小莉等,2020)。正常水分下杉木幼苗生长所需的氮素营养已得到满足,氮肥的施加使得N1组氮素过量,对杉木产生了一定的毒害作用,而干旱条件下过量的氮素也没能消除干旱对植物生长的负面影响(王清奎等,2015;陈云玉等,2016)。氮素对中度和重度干旱程度下青冈根系的影响不同,在重度干旱下总体表现为促进作用,这表明外源氮素添加提高了幼苗的抗旱性以应对重度干旱环境对生长的威胁(Drewniak et al.,2017)。细根(≤2 mm)是根系中活跃性和敏感度最高的部分,根系对养分有效性的响应大多是通过细根生物量的调整(Knute,2000;Poorter et al.,2015)。因此,施氮后干旱下杉木和青冈的细根变化相较于粗根更敏感。 今后,为缓解干旱对亚热带树种幼苗期造成的伤害,应注重协调不同植物的水氮关系,并依据树种对环境因子需求和适应的差异性,进行合理的水肥管理,从而促进干旱下植物幼苗对养分的吸收利用能力以及自身抗旱能力的提升。 氮素添加后杉木和青冈的生物量积累与分配和根系形态对干旱的响应具有明显的树种差异。植物自身抗旱机理的复杂性也导致两树种对土壤水肥因子的变化表现出不同的适应策略,适度的氮素营养可以弥补干旱对两树种地上部生物量累积与分配产生的不利影响,其中,施加氮肥后中度干旱组青冈表现较敏感,而氮素对杉木的作用效果则在重度干旱下更强。此外,施氮能够促使干旱下青冈幼苗根系形态的塑造,不断增强植物的耐受性以优化对有限水分的获取,尽管氮添加未对干旱下杉木根系形态产生积极的生态效应,但幼苗期的杉木亦能通过权衡自身地上及地下的物质分配来抵御不利环境的威胁。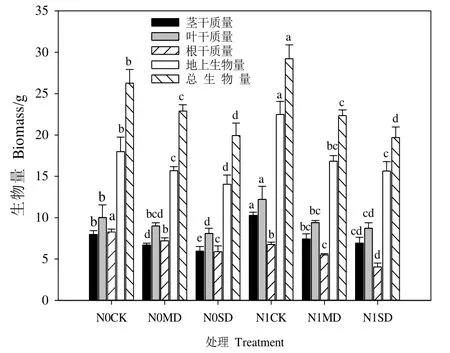
2.2 施氮对干旱下杉木和青冈幼苗生物量分配格局的影响

2.3 施氮对干旱下杉木和青冈幼苗根系形态特征的影响


3 讨论
3.1 施氮后幼苗生物量累积对干旱的响应
3.2 施氮对干旱下幼苗生物量分配格局的影响
3.3 氮素添加对干旱下幼苗根系形态特征的影响
4 结论
