不同物候期晚熟桃‘秋彤’微生物群落结构分析
2022-02-16姜莉莉郭腾达宫庆涛武海斌孙瑞红
姜莉莉,郭腾达,宫庆涛,武海斌,孙瑞红
(山东省果树研究所,山东泰安 271000)
0 引言
桃(Prunus persica)属蔷薇科李属植物,其果实肉质鲜美,富含多种维生素和微量元素,深受消费者喜爱,是全球广泛栽培的重要核果类果树之一[1]。近年来国内桃栽培面积和果品产量稳步上升[2]。桃是山东省第二大水果树种,产量和面积均居全国首位,已成为蒙阴等地重要的农业支柱产业[3]。‘秋彤’是从美国引入的晚熟硬溶质桃品种,于2014年通过山东省林木品种审定委员会审定。该品种果实大、绒毛短、色泽艳、风味甜、耐储运,适于在山东及邻近产区栽培[4]。
由于部分地区栽培管理技术落后以及苗木跨区引入等原因,桃树病害发生严重,据报道影响桃生产的病原菌多达90多种[5]。桃树疮痂病是由半知菌亚门黑星孢属嗜果枝孢菌(Cladosporium carpophilum)侵染引起,使病果产生“龟裂”现象[6]。桃褐腐病可由3种链核盘菌属(Monilinia)真菌侵染引起,分别为M.fructicola、M.laxa和M.fructigena,引起果实腐烂,又称“果腐病”[7]。桃树炭疽病病原为炭疽菌Colletotrichum,包括C.gloeosporioides、C.acutatum和C.truncatum,主要侵染果实和枝干,病果出现同心环状皱缩[8]。桃树黑斑病病原为链格孢属真菌(Alternaria alternata),主要危害果实桃尖。桃细菌性穿孔病由黄单胞菌(Xanthomona campestris)侵染引起,主要危害叶片,引起裂纹和穿孔[9]。桃树流胶病发病原因复杂,葡萄座腔菌属(Botryosphaeria)是侵染性流胶病的最主要病原菌[10]。桃白粉病的病原菌包括三指叉丝单囊壳菌(Podosphaera tridactyta)和桃单壳丝菌(Sphaerotheca pannosa),主要危害幼苗叶片[11]。桃软腐病由接合菌亚门匍枝根霉菌(Rhizopus stolonifer)侵染引起,主要发生在果实成熟及储运期。
当前对桃树病原菌的研究多采用传统方法,仅对可培养微生物进行分离鉴定,无法对微生物群落结构进行整体分析。高通量测序是近年来分析微生物多样性的热门技术,具有测序深度高、可鉴定低丰度物种信息等优点,能够更准确地反映微生物的真实组成和分布[12]。本研究拟通过高通量测序技术,分析晚熟桃“秋彤”在萌芽期、花期、幼果期、硬核期、果实膨大期及成熟期不同部位的真菌和细菌群落组成及多样性,为‘秋彤’桃的科学管理及病害节药防控提供理论依据。
1 材料与方法
1.1 样品采集
试验选址于山东省临沂市蒙阴县垛庄镇宝增果品专业合作社(118°7′8″E,35°34′1″N),为山地果园,‘秋彤’为当地主栽品种之一。随机选取3个‘秋彤’种植地块,每地块为1个重复,面积不小于200 m2,每个重复内随机选取5株桃树进行取样。
供采样桃树采用常规栽培管理,不套袋处理。2019年3月9日喷施80%代森锰锌800倍液(陶氏益农农业科技(中国)有限公司)、0.15%梧宁霉素(武汉埃格瑞生物技术工程有限公司)600倍液和3.2%甲维盐·氯氰微乳剂(海利尔药业集团股份有限公司)用于防治褐斑病、腐烂病和食心虫等;4月24日喷施30%苯甲·吡唑酯悬浮剂(上海生农生化制品股份有限公司)3000倍液和40%毒死蜱水乳剂(陕西韦尔奇作物保护有限公司)2000倍液防治褐斑病、蚧壳虫和蚜虫等;5月19日喷施21%噻虫嗪悬浮剂(天津施普乐农药技术发展有限公司)4000倍液和25%灭幼脲悬浮剂(河北中天邦正生物科技股份公司)2000倍液防治蚜虫、食心虫和卷叶蛾等;7月5日喷施20%吡唑醚菌酯可湿性粉剂(海利尔药业集团股份有限公司)2000倍液和20%氟硅唑可湿性粉剂(海利尔药业集团股份有限公司)3000倍液防治炭疽病等;8月1日喷施40%唑醚·咪鲜胺水乳剂(陕西韦尔奇作物保护有限公司)2500倍液防治炭疽病等。
分别于2019年3月7日(萌芽期,germination stage,Ger)采集桃树枝条(branch,Bran)和花蕾(bud),于4月9日(花期,flower,Fl)采集花朵(flower,Fl)、枝条和叶片(leaf,Le),于5月13日(幼果期,young fruit,Yo.Fr)、6月16日(硬核期,hardcore,Hard)、7月26日(果实膨大期,fruit growth,Fr.Gr)和8月23日(成熟期,mature,Mat)采集果实(fruit,Fr)、枝条和叶片。
以蘸有0.2 mol/L磷酸缓冲液的无菌拭子擦取枝条、叶片和果实表面的微生物,将拭子置于无菌离心管中,干冰保存送样。将采集到的15枚桃花蕾和花朵分别置于25 mL无菌磷酸缓冲液中,50 Hz超声处理20 min后取出花蕾和花朵,浸提液以0.22 μm滤膜过滤后,将滤膜置于无菌离心管,干冰保存送样[13]。真菌和细菌群落结构的高通量分析由北京诺禾致源生物信息科技有限公司代理完成。
1.2 测序分析
采用CTAB方法提取样本的基因组DNA,之后利用琼脂糖凝胶电泳检测DNA的纯度和浓度,使用无菌水将样本DNA浓度调整至1 ng/μL。
以上述基因组DNA为模板,使用带Barcode的特异性引物和Phusion®High-Fidelity PCR Master Mix with GC Buffer(New England Biolabs)进行PCR扩增。其中16SV4区引物为515F和806R,ITS1区引物为ITS1-1F-F和ITS1-1F-R。
PCR产物经琼脂糖凝胶电泳检测后,根据浓度进行等量混样,以1×TAE浓度2%的琼脂糖胶电泳进行纯化,以GeneJET胶回收试剂盒(Thermo Scientific)回收产物。
使用Ion Plus Fragment Library Kit 48 rxns建库试剂盒(Thermofisher)构建文库,经过Qubit定量和检测合格后,进行Ion S5TMXL(Thermofisher)上机测序。
1.3 高通量数据分析
对reads进行低质量剪切,截去Barcode和引物序列,去除嵌合体序列,得到有效数据。以Uparse(7.0.1001)软件对各样品的有效数据进行聚类,将一致性高于97%的序列聚类为OTUs,选取频数最高的序列作为代表性序列。用Mothur方法与SSUrRNA数据库对OTUs序列进行物种注释分析,获得分类学信息,并分别在界、门、纲、目、科、属和种7个分类水平统计各样本的群落组成。以数据量最少的样本为标准进行均一化处理,并以此进行后续的α-多样性分析(Chao1、Shannon、Simpson、ACE、Goods-coverage和PD_whole_tree指数)和β-多样性分析(NMDS和LEfSe分析)。
使用 Qiime软件计算 Chao1、Shannon、Simpson、ACE、Goods-coverage和PD_whole_tree指数,使用R软件(2.15.3)绘制稀释曲线和NMDS图,使用LEfSe软件进行LEfSe分析。
2 结果与分析
2.1 各样本测序深度与数据质控
由图1可以看出,随着测序深度的增加,各样本的稀释曲线末端均逐渐趋于平缓,表明测序基本饱和,当前测序深度能够体现各样本的细菌和真菌群落结构组成。

图1 桃不同物候期各部位样本的16S和ITS稀释曲线
2.2 ‘秋彤’桃不同物候期细菌和真菌的α-多样性指数比较
由表1可以看出,对细菌而言,‘秋彤’桃萌芽期花蕾、花期枝条和叶片、幼果期枝条和叶片的Shannon指数显著低于硬核期、果实膨大期和成熟期各部位;萌芽期花蕾和花期叶片的Simpson指数均低于硬核期、果实膨大期和成熟期各部位;萌芽期花蕾、花期叶片、幼果期和成熟期叶片的Chao1指数和ACE指数均低于其他处理。对于真菌而言,花期和幼果期各部位的Shannon指数和Simpson指数均高于硬核期;幼果期各部位Chao1指数和ACE指数均高于其他时期。
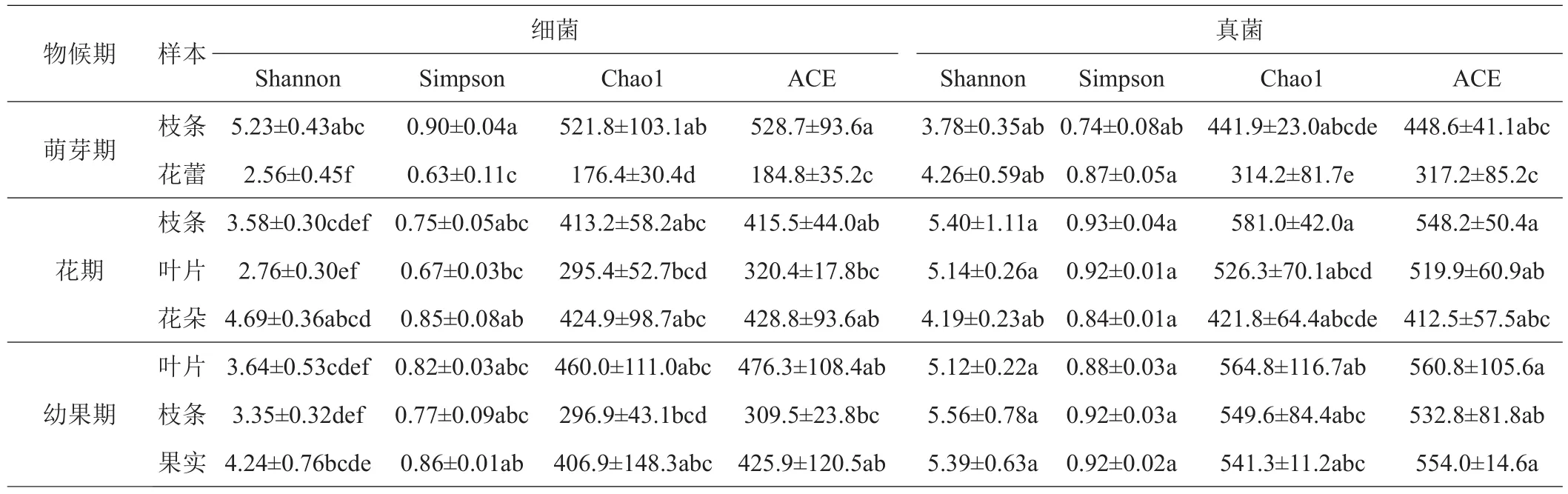
表1 桃不同物候期细菌和真菌的α-多样性指数
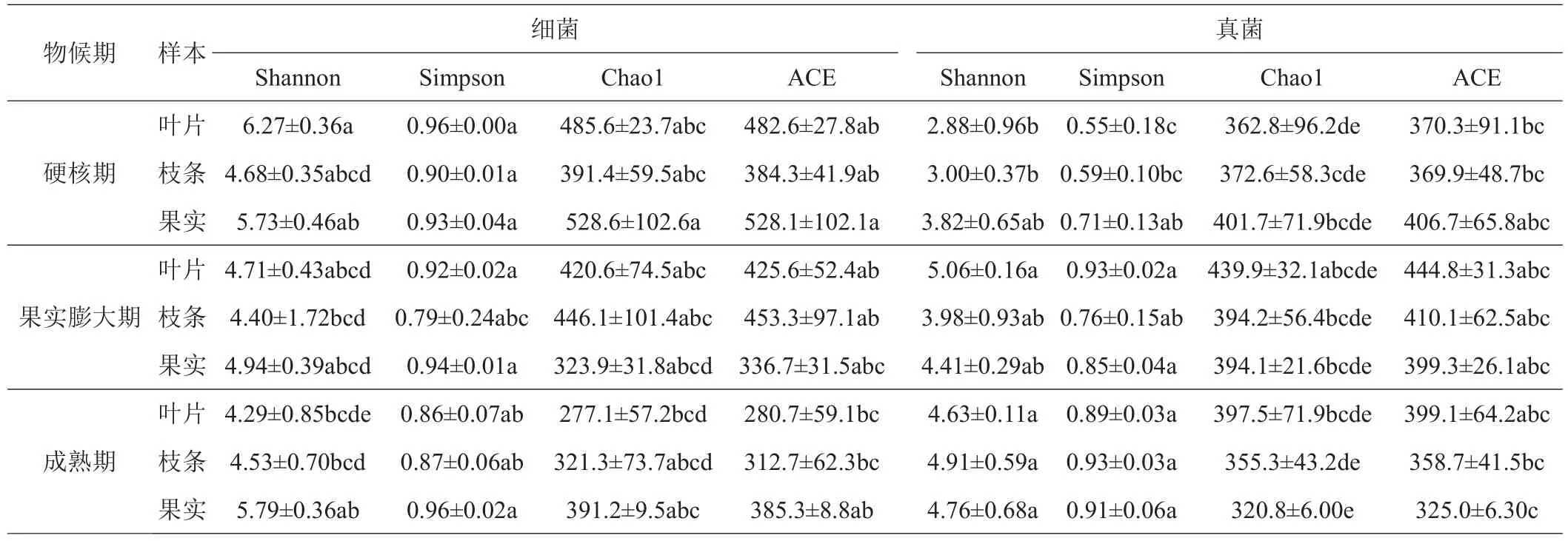
续表1
2.3 ‘秋彤’桃不同物候期、部位的细菌、真菌NMDS分析
由图2~3可以看出,桃同一物候期不同部位的细菌、真菌NMDS距离较近,不同物候期间距离较远,表明‘秋彤’桃树体表面的细菌、真菌受物候期影响较大,受部位影响较小。
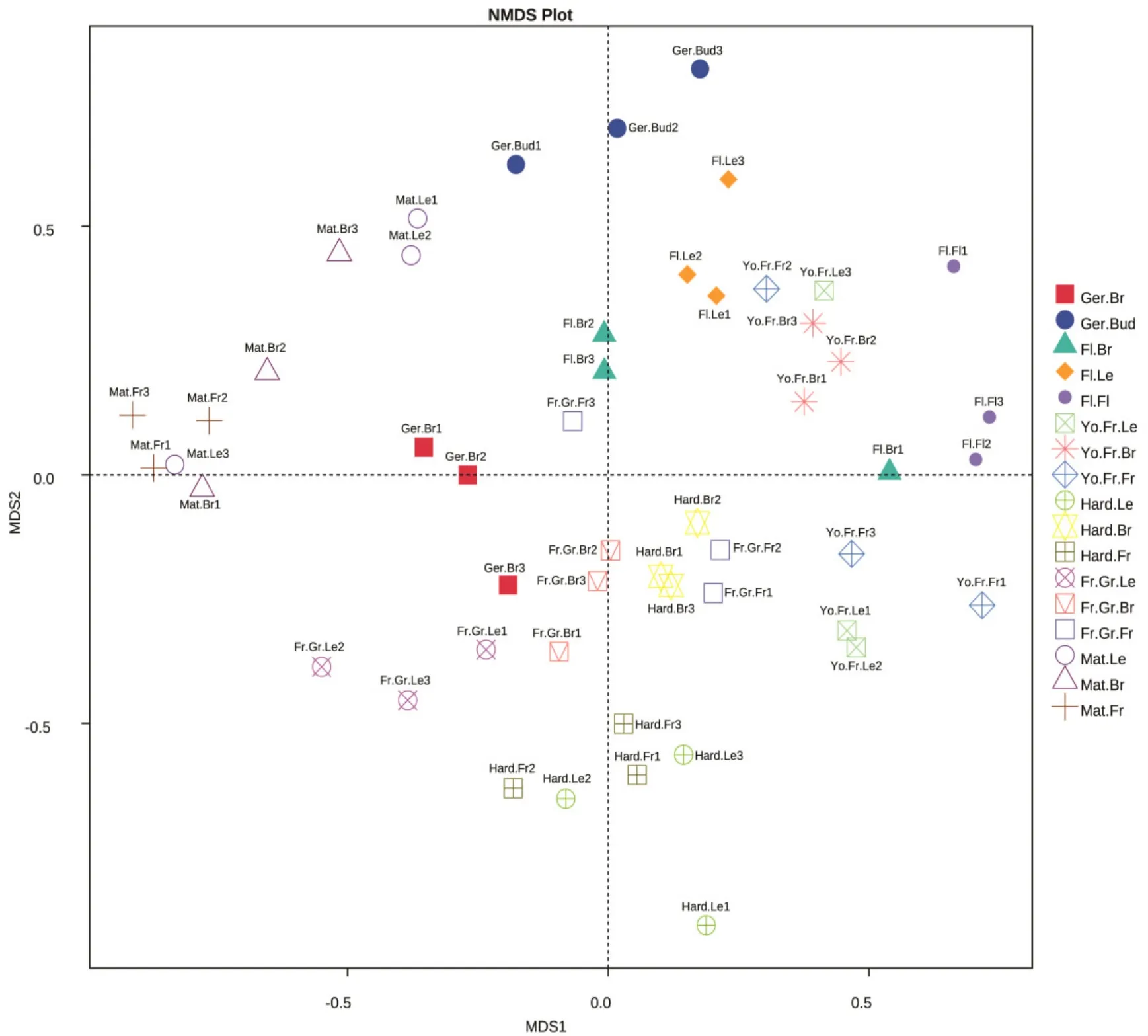
图2 桃不同物候期、部位的细菌NMDS分析
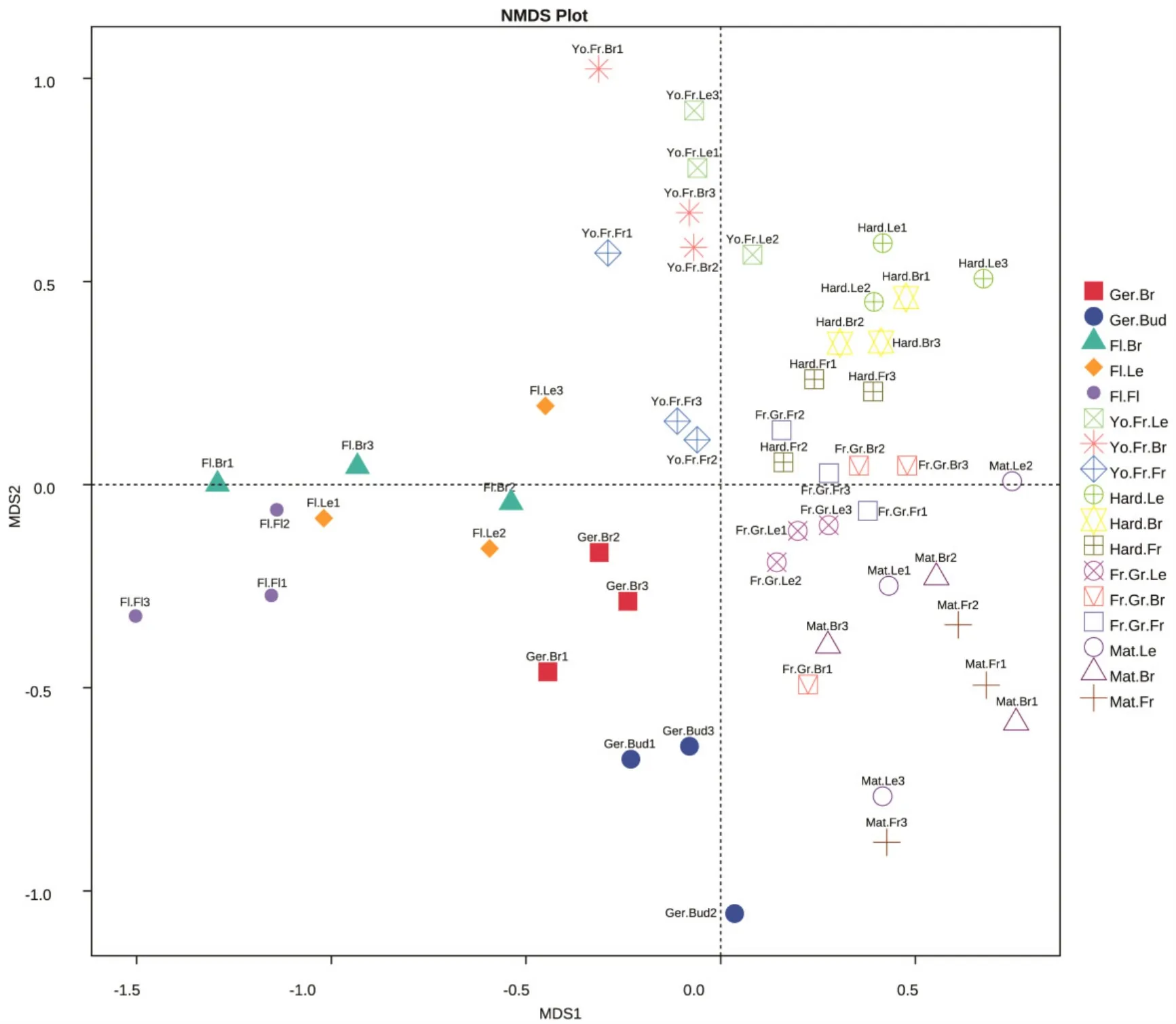
图3 桃不同物候期、部位的真菌NMDS分析
2.4 同一部位不同物候期的细菌相对丰度比较
分析‘秋彤’桃花期、幼果期、果实膨大期和成熟期叶片细菌差异(图4)发现,成熟期的拜叶克林氏菌和甲基杆菌相对丰度显著提高,果实膨大期的鞘脂杆菌、马赛菌、丙型变形菌和伯克氏菌相对丰度显著降低,其他时期未发现相对丰度差异显著的细菌。

图4 桃叶片不同物候期差异显著的细菌
分析‘秋彤’桃花蕾和花朵的细菌差异(图5)发现,花蕾的蓝藻细菌的相对丰度显著提高,花朵的肠杆菌、放线菌、拟杆菌、青枯菌、不动杆菌、莫拉氏菌、伯克氏菌、丙型变形菌、假单胞菌和变形菌的相对丰度显著降低。

图5 桃花朵和花蕾差异显著的细菌
分析‘秋彤’桃枝条不同物候期细菌差异(图6)发现,幼果期蓝藻细菌相对丰度较高,成熟期变形菌、鞘脂单胞菌、甲基杆菌和马赛菌相对丰度较高,硬核期立克次氏体、沃特氏菌和青枯菌相对丰度较高,萌芽期根瘤菌、拜叶林克氏菌、甲基杆菌、放线菌、拟杆菌、微球菌、短小杆菌、气单胞菌、噬纤维菌、薄层菌和黄色单胞菌相对丰度较高,果实膨大期泛菌和蜂窝囊菌相对丰度较高。

图6 桃枝条不同物候期差异显著的细菌
分析‘秋彤’桃果实不同物候期细菌差异(图7)发现,幼果期蓝藻细菌相对丰度较高,成熟期变形菌、根瘤菌、鞘脂单胞菌、微球菌、微杆菌、短小杆菌、拟杆菌、噬纤维菌、柄杆菌、薄层菌、短波单胞菌、生丝单胞菌、动孢囊菌、动球菌、螺状菌和气单胞菌相对丰度较高,硬核期马赛菌、红杆菌和副球菌相对丰度较高,果实膨大期立克次氏体、黄杆菌和泛菌相对丰度较高。

图7 桃果实不同物候期差异显著的细菌
2.5 同一部位不同物候期的真菌相对丰度比较
分析‘秋彤’桃叶片不同物候期真菌差异(图8)发现,幼果期锤舌菌和白粉菌相对丰度较高,成熟期外担菌相对丰度较高,果实膨大期枝孢菌相对丰度较高,花期粪壳菌、担子菌、赤霉菌、拟盘多毛孢、银耳目和酵母菌相对丰度较高。
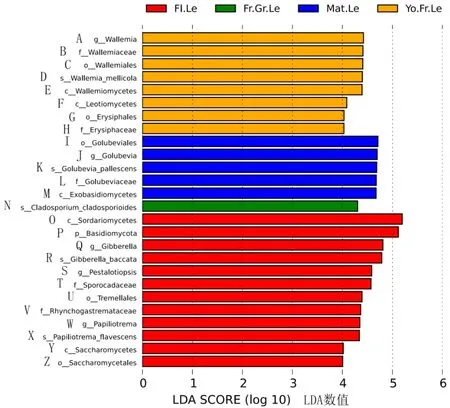
图8 桃叶片不同物候期差异显著的真菌
分析‘秋彤’桃花朵和花蕾的真菌差异(图9)发现,花蕾的座囊菌、短梗霉菌、银耳和担孢酵母的相对丰度显著提高,花朵的炭角菌、子囊菌、担子菌、拟盘多毛孢、角担菌、鸡油菌和伞菌相对丰度显著降低。
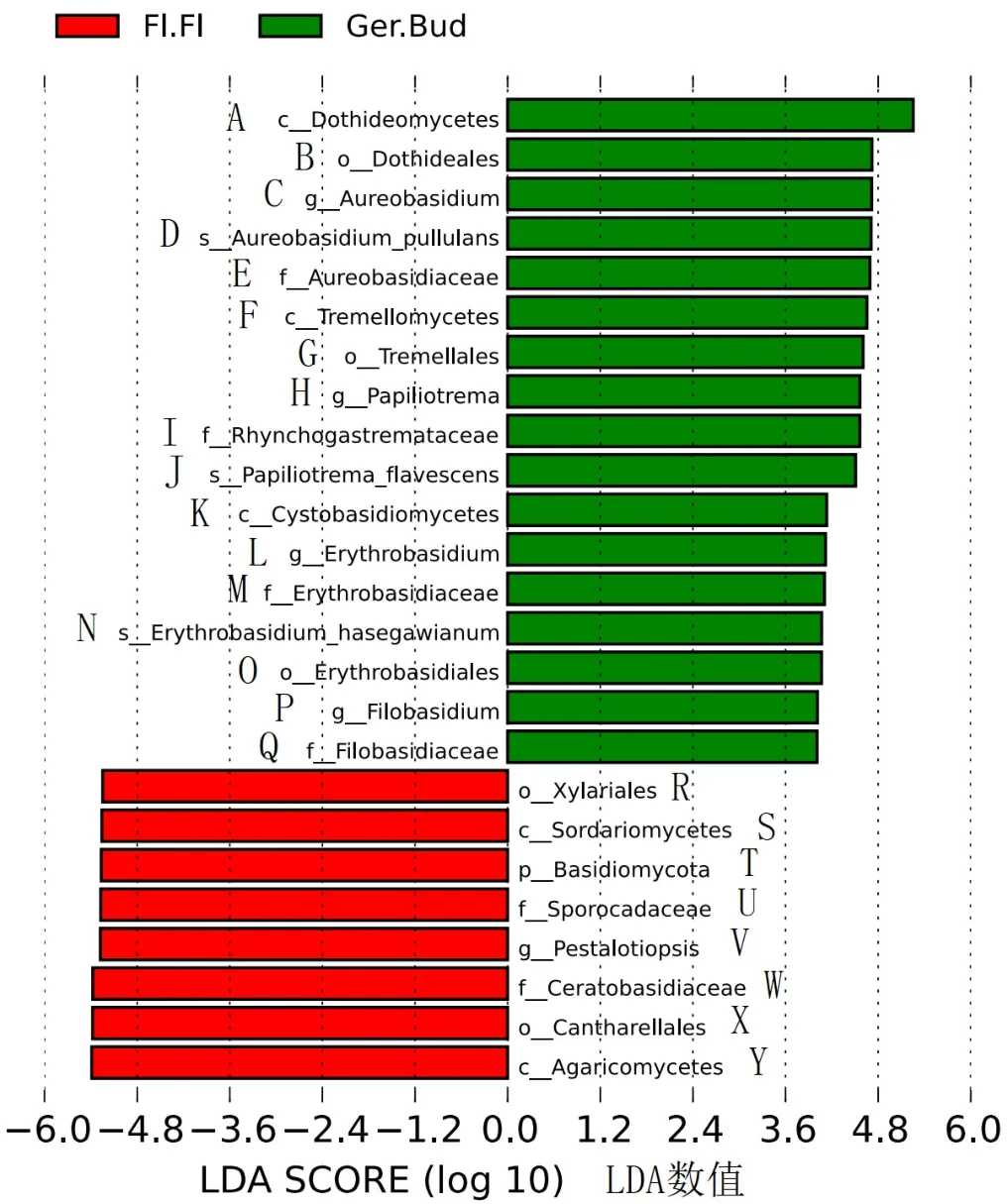
图9 桃花朵和花蕾差异显著的真菌
分析‘秋彤’桃枝条不同物候期真菌差异(图10)发现,成熟期外担菌和赤霉菌相对丰度较高,硬核期座囊菌、链格孢菌、子囊菌、附球菌和亚隔孢壳菌相对丰度较高,萌芽期枝孢菌相对丰度较高,花期担子菌、伞菌、鸡油菌、子囊菌、拟盘多毛孢和酵母相对丰度较高。
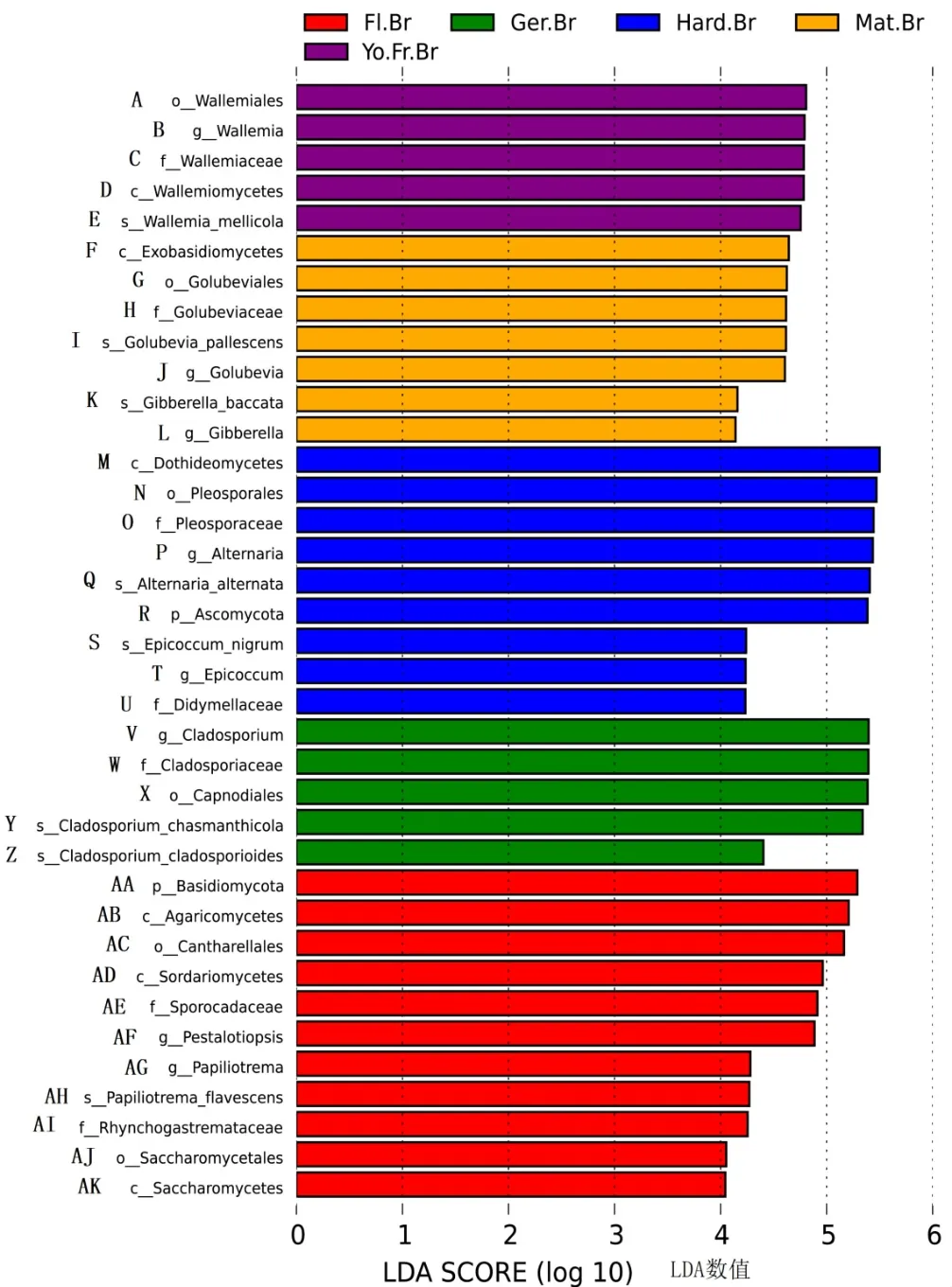
图10 桃枝条不同物候期差异显著的真菌
分析‘秋彤’桃果实不同物候期真菌差异(图11)发现,成熟期担子菌相对丰度较高,硬核期座囊菌、腔菌、子囊菌和链格孢菌的相对丰度较高。
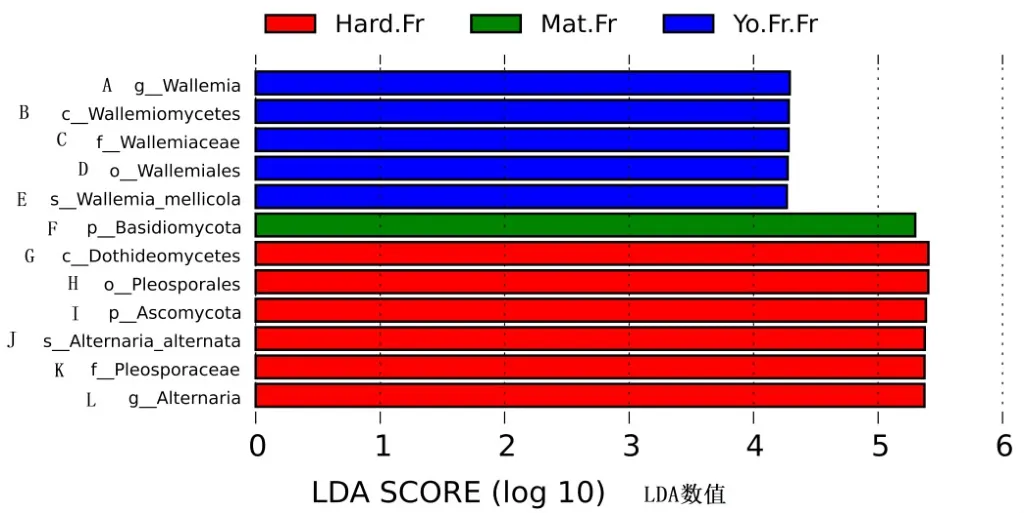
图11 桃果实不同物候期差异显著的真菌
3 结论与讨论
高通量测序技术可灵敏地检测到低丰度、不可培养的物种信息,能够更全面客观地揭示目标微生物群落结构,已成为现阶段研究微生物群落结构的重要手段[14]。本研究采用高通量测序技术分析了‘秋彤’桃不同物候期、部位的细菌和真菌群落结构,得到了3968663条细菌clean reads,属于539个属;以及4020830条真菌clean reads,属于334个属。
α-多样性主要表征局域生境下的物种数目,也被称为生境内的多样性[15]。其中Shannon指数关注样品中的分类总数和每个分类所占的比例,用于表征物种的多样性和均匀性。Simpson指数用于评估常见物种优势度[16]。Chao1和ACE指数可表征群落物种丰度,其中Chao1能够较好地反映群落中低丰度物种的存在,ACE能够反映群落总体情况[17]。牛世全等[18]采用高通量测序技术分析河西走廊地区盐碱土壤的微生物多样性,发现原生盐碱土的Shannon指数最小,表明其微生物多样性低。本研究发现,对于细菌,‘秋彤’桃萌芽期和幼果期各部位的Shannon和Simpson指数均低于硬核期、果实膨大期和成熟期各部位,表明后期的细菌多样性高于前期;花期和成熟期叶片的Chao1和ACE指数均较低,表明其物种丰度较低;对于真菌,花期和幼果期各部位的Shannon和Simpson指数均高于硬核期,表明前期的真菌多样性较高,幼果期各部位Chao1指数和ACE指数均高于其他时期,表明其真菌物种丰度较高。综合比较可以发现,随着‘秋彤’桃物候期的推移,各部位细菌多样性提高,真菌多样性降低,这可能与田间温湿度条件变化、药剂管理措施及微生物互作等有关。
NMDS(Non-Metric Multi-Dimensional Scaling,无度量多维尺度排序)分析是根据物种信息,将样本以点的形式展示在二维平面上,克服了线性模型(PCA、PCoA等)的缺点,能够更好地反映生态学信息[19]。路颖等[20]采用高通量测序技术分析了泰山不同造林树种掉落物分解对细菌群落的影响,NMDS分析表明,细菌群落多样性受掉落物化学性质影响较大。叶雯等[21]采用高通量测序技术分析了不同种植年限香榧根际土壤微生物的群落结构特征,NMDS结果显示,相同种植年限土壤真菌群落距离较近,不同种植年限之间能明显分开,表明其微生物群落结构可随种植年限的不同而发生明显变化。本研究发现,桃同一物候期不同部位的细菌、真菌NMDS距离较近,不同物候期间距离较远,表明‘秋彤’桃树体表面的细菌、真菌受物候期影响较大,受部位影响较小。
农事操作包括农药的使用对果树微生物群落结构影响较大[22-23]。刘晓静等[24]采用高通量测序技术分析油桃和小白杏采后果实内生细菌群落结构,发现泛菌属(Pantoea)为优势菌群,其中菌株XAAS-P1是导致采后油桃和小白杏软腐变质的主要病原细菌。王柯等[25]通过高通量测序证实,苦参碱处理可以降低土壤腐皮镰孢菌、层出镰孢菌、串珠镰孢菌和尖孢镰孢菌的基因拷贝数,有效缓解平邑甜茶连作障碍。本研究详细记录了‘秋彤’桃各物候期的用药情况,比较同一部位、不同物候期的细菌、真菌丰度发现,对细菌而言,花期花朵和硬核期枝条青枯菌丰度较高,萌芽期枝条和成熟期果实根瘤菌、噬纤维菌等丰度较高,果实膨大期枝条和果实泛菌等丰度较高。对真菌而言,幼果期叶片白粉菌丰度较高,果实膨大期叶片和萌芽期枝条枝孢菌(桃疮痂病病原菌)丰度较高,花期叶片和成熟期枝条赤霉菌等丰度较高,硬核期枝条和果实链格孢菌(桃黑斑病病原菌)等丰度较高。由此可见,在现有管理模式下,晚熟桃‘秋彤’硬核期受黑斑病危害风险较高,硬核期和萌芽期受桃疮痂病危害风险较高,生产中可使用苯醚甲环唑、多抗霉素等药剂进行防控。
