陆地棉细胞色素P450基因GhP450-94C1黄萎病抗性功能验证
2022-02-15胡子曜李秀青代培红雷建峰柳建飞邓嘉辉刘晓东
胡子曜,李秀青,代培红,雷建峰,柳建飞,赵 燚,邓嘉辉,刘 超,刘晓东,李 月
(1.新疆农业大学 生命科学学院,新疆 乌鲁木齐 830052;2.新疆农业大学 农学院,新疆 乌鲁木齐 830052)
细胞色素P450(Cytochrome P450,CYP450)是最大的蛋白质超家族之一,因其与还原态CO结合后在450 nm处吸光值最高而得名,其广泛存在于植物、微生物、昆虫及哺乳动物细胞内[1]。目前对该超家族蛋白按CYP(细胞色素的正式缩写)、家族(数字)、亚家族(字母)和同工型(数字)顺序命名,例如CYP94C1属于CYP家族94亚家族C中的一类蛋白质[2]。CYP450基因家族在植物基因组中数目庞大,约占编码蛋白质基因总数的1%[3];CYP450蛋白在进化上高度保守,暗示其功能具有一定的相似性,通常与线粒体、高尔基体及内质网等细胞器膜结合在一起,广泛参与萜类、激素、苯丙烷、脂肪酸、生物碱、信号分子及类黄酮的合成与降解等多种植物生命活动中的次生代谢反应,在植物生长发育和提高植物对逆境胁迫的适应能力中发挥着重要作用[4]。Ping等[5]过表达苹果CYP450基因MdCYPM1后,植株的生长素运输及开花时间均受到显著影响,表现出叶绿素的减少及下胚轴伸长等表型,表明该基因在调控植物的生长发育上发挥着重要作用;Xiao等[6]研究发现,CYP86A8和CYP86A22个基因均通过参与拟南芥角质层的合成进而增强植株的耐旱性。Chopra等[7]研究发现,高粱CYP99A1和CYP709C1基因的表达受低温诱导显著,推测它们可能在抵御冷胁迫中发挥一定的作用;Wang等[8]研究表明,小麦CYP450基因TaCYP81D5通过参与植株体内的活性氧清除进而介导植株的抗盐反应;而Mao等[9]发现,拟南芥AtCYP709B3基因的表达受高盐胁迫诱导,推测其可能与小麦TaCYP81D5基因功能类似,同样介导植株的耐盐反应;Zhang等[10]过表达苜蓿CYP2E1基因后显著增强了植株的抗汞毒性;Yan等[11]过表达GmCYP82A3基因后显著增强了转基因大豆对疫霉菌的抗性,且JA、ET信号通路相关基因表达量显著上调,推测该基因通过参与JA和ET信号通路的正向调控,介导大豆的抗病反应。
棉花是关系我国民生的重要经济作物之一[12]。黄萎病是棉花生产最具破坏性的生物胁迫之一,严重制约着我国的棉花产量及纤维品质,阻碍着国民经济的发展。大丽轮枝菌(VerticilliumdahliaeKleb)是引发我国棉花黄萎病的主要病原菌,作为一种土传维管束真菌病害,其通过根系感染,继而引起植株的枯萎、变色、落叶,最终导致植株的死亡[13]。大丽轮枝菌寄主广泛,可以侵染包括棉花在内的200多种双子叶植物;微菌核是大丽轮枝菌的休眠状态,对不良环境具有很强的抗逆能力,因此,该病原菌在没有合适寄主的情况下也可以在土壤中长期存活[14]。目前,还缺乏可以有效防止大丽轮枝菌侵染的杀菌剂,因此,挖掘黄萎病相关抗性基因并培育抗病新品种,对于棉花黄萎病的防治及农业安全生产至关重要。
目前,对于细胞色素P450基因在棉花抗黄萎病反应中的研究主要集中在海岛棉。Zhou等[15]抑制海岛棉GbCYP94C1基因表达后降低了植株对黄萎病菌的敏感性,表明该基因是海岛棉抵御黄萎病菌入侵的负调节因子;Wang等[16]过表达海岛棉GbCYP86A1-1基因后通过激活相关抗病免疫信号途径,增强了植株对黄萎病菌的耐受性;但目前关于P450家族基因参与陆地棉抗黄萎病反应的相关研究还未见报道。
本研究通过前期转录组数据筛选得到一个响应黄萎病菌侵染的细胞色素P450基因GhP450-94C1,利用病毒诱导的基因沉默(VIGS)技术初步探究其在棉花抵御黄萎病菌的应激反应中的生物学功能,旨在为挖掘、鉴定陆地棉黄萎病相关基因提供一定的理论基础。
1 材料和方法
1.1 试验材料
供试材料为陆地棉遗传标准系TM-1,其种子由新疆农业大学生命科学学院农业生物技术重点实验室繁存;大丽轮枝菌菌株V991及TRV病毒载体均由新疆农业科学院黄全生研究员惠赠。FastPfu Fly DNA聚合酶、DH5α和GV3101感受态细胞、B-Zero平末端克隆载体、T4DNA 连接酶和DNA分子量Marker均购于北京全式金公司;各种限制性内切酶购于赛默飞(Thermo)公司,琼脂糖凝胶回收试剂盒和RNase A购于南京诺维赞公司,RNA提取试剂盒购于杭州博日公司;反转录和荧光定量试剂盒均购于加拿大爱必梦(abm)公司;抗生素化学试剂均购于北京索莱宝公司;试验所用引物的合成及测序均由上海生工公司完成。
1.2 试验方法
1.2.1 棉花种植 挑选颗粒饱满的TM-1种子,参照Li等[17]的方法进行培育及种植。
1.2.2 黄萎病菌的培养及诱导处理 将-80 ℃保存的大丽轮枝菌V991取出后,参照Li等[17]的方法进行活化培养及接菌处理,对照组(CK)为Czapek′s培养基处理,分别于处理前(0 h)和处理后6,12,24,48,72 h 对其根部进行取样;每个时间点均取4个生物学重复。
1.2.3 总RNA提取与cDNA合成 使用RNA提取试剂盒对棉花根部样品进行RNA的提取,经1.5%的琼脂糖凝胶电泳检测其完整性后,使用反转录试剂盒进行cDNA合成。
1.2.4 棉花GhP450-94C1基因的克隆及序列分析 通过转录组数据分析获得GhP450-94C1基因序列,并在NCBI(https://www.ncbi.nlm.nih.gov/)数据库中检索其登录号:XM_016813513。GhP450-94C1基因生物信息学分析工具参见表1。根据该基因的CDS序列设计相应的克隆引物(表2),以反转录获得的cDNA为模板,参照胡子曜等[18]的方法进行基因克隆。

表1 生物信息学分析工具Tab.1 Bioinformatics analysis tools and corresponding functions
1.2.5GhP450-94C1基因在黄萎病菌V991侵染下的表达模式 以反转录的cDNA为模板,选取GhUBQ7为内参基因[19],利用qRT-PCR技术探究GhP450-94C1基因在黄萎病菌侵染下的表达模式。反应结束后参照各基因的Ct值,使用2-ΔΔCt方法计算目的基因的相对表达量[20]。相关引物见表2。
1.2.6GhP450-94C1基因的VIGS载体构建 根据GhP450-94C1基因的ORF(1503 bp)序列,利用SGN-VIGS(https://vigs.solgenomics.net/)在线网站设计抑制GhP450-94C1基因表达的沉默片段(442 bp),设计相应引物并在其上、下游5′端分别加入EcoR Ⅰ和KpnⅠ酶切位点(表2),克隆VIGS靶序列,具体方法同1.2.4。用测序结果正确的重组质粒及TRV2载体经EcoR Ⅰ和KpnⅠ酶切后用T4DNA Ligase连接4 h后转化DH5α感受态细胞并进行酶切鉴定,正确的重组质粒取10 μL转化GV3101感受态细胞,28 ℃培养2 d,用于后续试验。
1.2.7 农杆菌介导的VIGS侵染棉花及沉默效率检测 将TRV∶GhP450-94C1、TRV∶GhCLA1、TRV∶RNA1和TRV∶RNA2分别转化农杆菌后,选取生长一致、2片子叶完全展开且真叶未露出的棉花幼苗,参照Li等[17]的方法进行菌体的活化、重悬及VIGS侵染。大约21 d后,当阳性对照TRV∶∶GhCLA1的叶片出现明显的白化表型时,取各组分的根部及真叶组织进行RNA提取及cDNA合成,通过qRT-PCR检测沉默效率。相关引物见表2。
1.2.8 棉苗的接种处理 选取生长状况一致的GhP450-94C1沉默植株和阴性对照植株,参照Li等[17]的方法进行黄萎病菌(V991)的接种处理。
1.2.9GhP450-94C1基因的黄萎病抗性鉴定 接种黄萎病菌20 d后拍照记录GhP450-94C1沉默植株和阴性对照植株的表型差异并采用叶片分级法统计病情指数[17],同时对GhP450-94C1沉默植株和阴性对照植株进行剖秆检测及茎段恢复培养试验[21]。

表2 引物及用途Tab.2 Primer and application
2 结果与分析
2.1 GhP450-94C1基因的克隆与序列分析
以陆地棉TM-1的叶片cDNA为模板进行PCR扩增,大小与预期相符(图1),表明已成功克隆GhP450-94C1基因的ORF序列。该基因的ORF为1 503 bp,编码500个氨基酸,分子式为C2597H4025N691O725S22,分子质量为57.23 ku,定位于内质网膜,理论等电点为6.87,平均疏水性为-0.030,表明该蛋白为酸性、亲水性蛋白,脂肪系数为93.20,不稳定指数为46.44,属于不稳定蛋白,含有跨膜结构,属于跨膜蛋白;蛋白结构域预测结果显示,该蛋白含有一个P450结构域(图2);信号肽预测结果显示,GhP450-94C1蛋白有86.45%的概率存在信号肽(图3);二级结构预测结果显示,该蛋白含有24个α-螺旋和8个β-折叠,三级结构预测结果完全符合二级结构特征(图4)。

M.Trans 2000 PlusⅡ DNA Marker。图6同。M.Trans 2000 PlusⅡ DNA Marker.The same as Fig.6.

图2 GhP450-94C1蛋白结构域预测Fig.2 The prediction protein domain of GhP450-94C1

图3 GhP450-94C1蛋白信号肽预测Fig.3 The prediction signal peptide of GhP450-94C1 protein
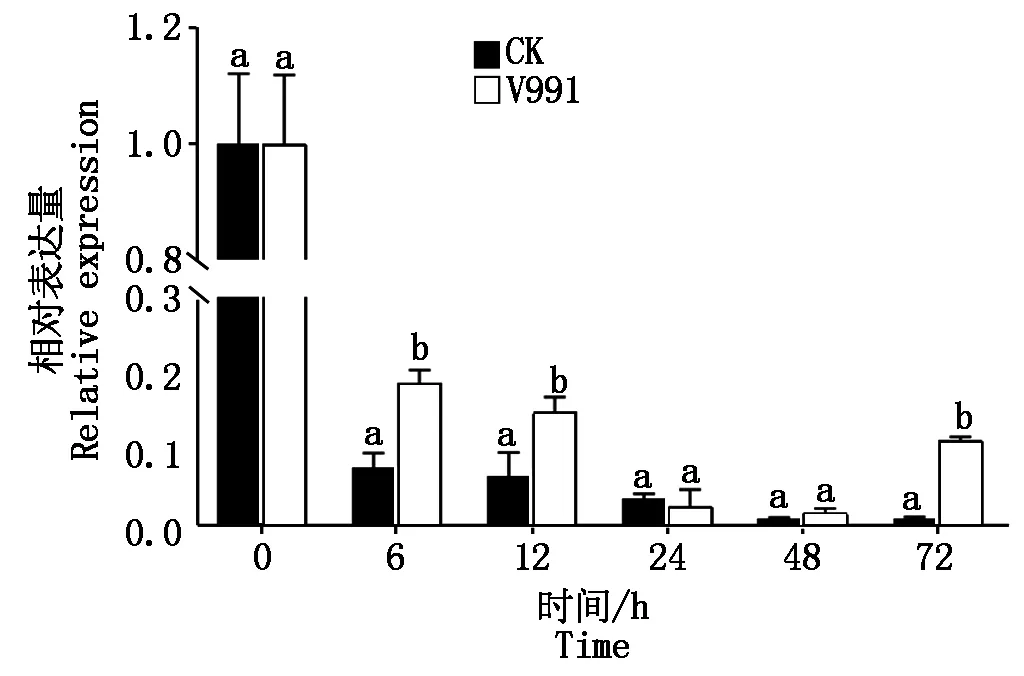
2.2 黄萎病菌V991处理下GhP450-94C1基因的表达模式分析
黄萎病菌V991处理后(图5),与对照相比GhP450-94C1基因表达量在6,12,72 h显著上调(P<0.05),24,48 h无显著差异。表明GhP450-94C1基因响应黄萎病菌胁迫处理,可能在棉花抵御黄萎病菌侵染反应中发挥一定的作用。
2.3 GhP450-94C1基因的VIGS载体构建
载体构建示意图如图6-A所示,利用PCR技术扩增抑制目标基因表达的靶序列(CM-GhP450-94C1),大小符合预期。将其进行克隆并测序,测序正确的质粒酶切后与TRV2载体连接并进行双酶切鉴定,结果呈2条带(TRV∶GhP450-94C1),一条8 000 bp以上的载体条带和一条约500 bp的目标片段(图6-B),表明已成功构建GhP450-94C1基因的VIGS载体。
2.4 GhP450-94C1基因的沉默效率检测
将TRV∶GhP450-94C1、TRV∶GhCLA1和TRV∶RNA2分别与TRV∶RNA1混合后进行棉花侵染。15 d后进行表型观察,与阴性对照TRV∶00相比,阳性对照TRV∶GhCLA1的叶片出现明显的白化现象(图7-A);qRT-PCR检测结果显示,GhCLA1和GhP450-94C1基因的表达均受到显著抑制(图7-B、C),表明TRV-VIGS体系可以在棉花体内正常工作并成功获得GhP450-94C1沉默植株。

A.GhP450-94C1蛋白的二级结构预测;B.GhP450-94C1蛋白的三级结构预测。A.The prediction secondary structure of GhP450-94C1 protein;B.The prediction tertiary structure of GhP450-94C1 protein.
相同小写字母表示差异不显著(P>0.05),不同小写字母之间表示差异显著(P<0.05)。图7—8同。
2.5 GhP450-94C1基因沉默植株的黄萎病抗性鉴定
选取生长状况一致的GhP450-94C1基因沉默植株和阴性对照植株进行黄萎病菌侵染,20 d后拍照记录其表型差异并统计病情指数及剖秆检测和茎段恢复培养试验。结果显示,与对照植株相比,沉默植株的萎蔫程度及叶片黄化更加严重,其健康状况明显更差(图8-A);病情指数统计结果显示,沉默植株病情指数显著高于对照植株(P<0.05)(图8-B);剖秆检测结果显示,与对照植株相比,沉默植株茎秆的褐变程度更为严重(图8-C);茎段恢复培养结果显示,沉默植株的茎段在培养基上长出的真菌菌丝数量明显多于对照植株,(图8-D)。以上结果初步表明,GhP450-94C1基因是一个棉花抵御黄萎病菌入侵的正调节因子。

A.GhP450-94C1的VIGS载体构建示意图;B.GhP450-94C1目标片段的PCR扩增;C.TRV∶GhP450-94C1载体的酶切鉴定。A.Construction of VIGS vector of GhP450-94C1;B.PCR amplification of target fragment of GhP450-94C1;C.Identification of TRV∶GhP450-94C1 vectors by restriction enzyme digestion.

A.沉默GhCLA1的表型;B、C.GhCLA1和GhP450-94C1的沉默效率检测。A.The phenotype of silenced GhCLA1;B,C.Silencing efficiency of GhCLA1 and GhP450-94C1.
3 结论与讨论
黄萎病被称为棉花的“癌症”,严重制约着棉花的纤维品质及扩大生产,已成为影响我国棉花质量及产量的首要生物因素。作为目前我国主要的栽培品种,陆地棉不同于海岛棉,其对黄萎病具有较强的抗性,对黄萎病菌较为敏感[22]。由于种间杂交育种难度大且效率低,抗病品种选育进展缓慢;因此,筛选、鉴定棉花抗黄萎病关键基因并利用现代基因工程和分子育种技术培育抗病新品种具有重要意义。
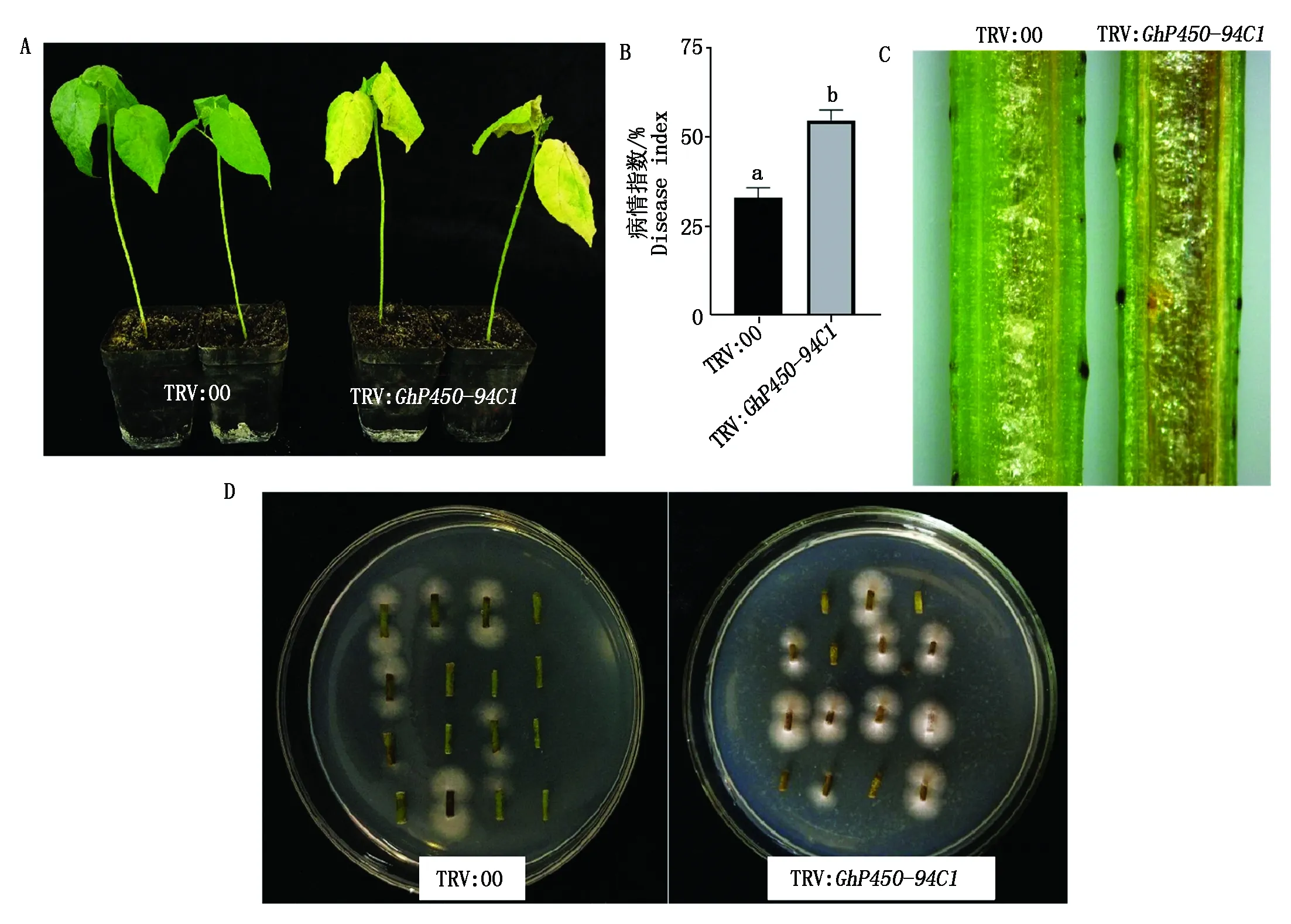
A.GhP450-94C1基因沉默后植株的表型分析;B.病情指数统计;C.剖秆鉴定;D.病原菌恢复培养。A.Phenotype analysis of plants after GhP450-94C1 gene silencing;B.Disease index statistics;C.Stem cutting phenotype;D.Restoration culture of pathogenic bacteria.
棉花对黄萎病的防御过程是十分复杂的,需要一系列抗性基因和相关蛋白的参与。随着棉花基因组测序的完成,越来越多的黄萎病抗性相关基因被克隆并鉴定出来:Zhang等[23]利用VIGS技术抑制棉花琥珀酸脱氢酶(SDH)基因GhSDH1-1表达后显著提高了植株对大丽轮枝菌的敏感性,且水杨酸(SA)通路相关基因表达显著下调,表明GhSDH1-1基因通过介导SA途径进而在棉花抗黄萎病反应中作为一个正调节因子;Li等[17]利用VIGS技术沉默棉花GhBsr-k1基因后显著增强了植株对大丽轮枝菌和尖孢镰刀菌的抗性,苯丙烷代谢途径相关基因的表达显著上调,木质素积累量显著增加,表明该基因是棉花抗病反应的一个负调节因子,其在植物防御中的潜在机制可能与调控木质素通路基因的转录有关;Xiong等[24]利用RNAi技术干涉WRKY转录因子GhWRKY70D13基因表达后显著提高了植株的抗病性,植株中乙烯(ET)和茉莉酸(JA)合成及信号通路基因表达均显著增加,表明该基因通过负调控ET和JA信号通路,参与了棉花抗病反应;Zhu等[25]研究发现,棉花PLPs基因GhPLP2通过调节脂肪酸的积累和JA信号通路,正向调节棉花的抗病反应;Xiong等[26]利用VIGS技术抑制GhGDH2基因表达后激活了JA合成及信号通路基因的表达但抑制了SA合成及信号通路基因的表达,增强了植株的抗病性,判断该基因是棉花抗病反应的一个正调节因子,其最终表现出的抗性是2种抗病途径效果总和抵消后的结果;Li等[27]过表达F-box蛋白基因GhACIF1后增强了转基因植株对大丽轮枝菌的抗性,表明该基因正向调控棉花对黄萎病菌的抗性;Pei等[28]研究发现,棉花GLP蛋白基因GhABP19通过激活JA途径和SOD活性,在棉花抵御黄萎病菌和枯萎病菌入侵中发挥着重要作用;Ma等[29]研究发现,棉花BEL1-Like转录因子GhBLH7-D06通过抑制木质素的生物合成和JA信号传导途径进而负调节棉花对黄萎病菌的抗性。此外,近年来还有GhAAT[30]、GbVIP1[31]、GbMPK3[32]、GhRPS6[33]、GhTBL34[34]、GbABR1[35]、GhSKIP35[36]等棉花黄萎病相关基因被挖掘、鉴定出来。
作为陆生植物中最大的酶蛋白家族之一,目前已有许多研究表明,细胞色素P450基因在植物抵御病原菌胁迫反应中发挥着重要作用。李文奇等[37]过表达水稻P450基因OsCYP71Z2后显著提高了转基因水稻对稻瘟病菌的抗性,植保素含量及生物合成通路关键基因的表达量均显著增加,推测该基因通过介导植保素的生物合成参与水稻对稻瘟病抗性反应的调控;Zhou等[38]从拟南芥中克隆得到一个CYP71B15基因,该基因同样通过调控植保素的生物合成,在作物抵御病原菌侵染中发挥着重要作用;Maeda等[39]研究发现,水稻CYP78A基因BSR2基因的敲除显著增强了植株对立枯病菌的敏感性;Li等[40]研究发现,小麦对枯萎病菌和赤霉病菌的抗性与细胞色素P450基因的激活密切相关;Liu等[41]研究发现,拟南芥细胞色素P450基因CYP82C2通过正向调节JA诱导的相关防御基因的表达,在植株抗灰霉菌反应中扮演一个正调控因子的角色。目前,对于陆地棉P450家族基因的功能研究多集中在对植株的生长发育的调控:张翼等[42]从陆地棉中克隆出一个P450基因GhCYP85A2-1,该基因表达后植株的株高显著降低,判断其在棉花株高发育过程中起着重要的调控作用;辛珊等[43]过表达GhCYP714A1基因后显著增强了转基因烟草叶片中过氧化氢(H2O2)量的积累,推测其可能参与了活性氧的产生;Gu等[44]研究发现,GhCYP94C1基因参与了WRKY转录因子GhWRKY27调控的陆地棉衰老途径;此外Sun等[45]发现棉花CYP82亚家族成员CYP82D基因通过介导细胞的系统性死亡,参与陆地棉抵御枯萎病菌的反应,表明陆地棉P450基因同样参与陆地棉抗病反应的调控;但目前关于该基因家族在陆地棉抗黄萎病反应中的功能研究尚无报道。
本研究通过前期转录组数据筛选并克隆出一个细胞色素P450基因GhP450-94C1,该基因的ORF长度为1 503 bp,编码500个氨基酸,分子式为C2597H4025N691O725S22,分子质量为57.23 ku,定位于内质网膜,所编码蛋白为酸性、亲水的不稳定跨膜蛋白,有86.45%的概率存在信号肽;含有一个P450结构域;此外该蛋白在结构上含有24个α-螺旋和8个β-折叠。该基因响应黄萎病菌V991侵染,可能在棉花抵御黄萎病菌入侵中发挥一定的功能;利用VIGS技术抑制该基因表达后降低了植株对黄萎病菌的抗性,其病情指数显著高于阴性对照植株;剖秆检测表明,与对照植株相比,沉默植株维管束的褐变程度更加严重;茎段恢复培养试验表明,沉默植株茎段的真菌菌丝数量明显多于对照植株。以上结果初步表明,GhP450-94C1基因是一个棉花抗黄萎病反应的正调节因子,然而该基因的抗病分子机制及参与的抗病信号通路仍需深入探究。
