棉花出苗期耐冷综合评价体系的构建及耐冷指标筛选
2022-02-01沈倩张思平刘瑞华刘绍东陈静葛常伟马慧娟赵新华杨国正宋美珍庞朝友
沈倩,张思平,刘瑞华,刘绍东,陈静,葛常伟,马慧娟,赵新华,杨国正,宋美珍,庞朝友
棉花出苗期耐冷综合评价体系的构建及耐冷指标筛选
沈倩1, 2,张思平1,刘瑞华1,刘绍东1,陈静1,葛常伟1,马慧娟1,赵新华1,杨国正2,宋美珍1,庞朝友
1中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳 455000;2华中农业大学植物科学与技术学院,武汉 430070
【目的】综合评价棉花品种(系)出苗期耐冷性,建立可靠评价模型,筛选鉴定指标,为耐冷品种选育鉴定提供简便有效的评价方法。【方法】以200份陆地棉品种(系)为试验材料,设恒定低温、昼夜变温和适宜温度3个处理,测定其出苗率、下胚轴长、根长、百粒重等指标,采用耐冷系数差异分析、频次分析、降幅分析、主成分分析、聚类分析和多元回归分析等方法,对群体进行耐冷型划分,并建立耐冷性评价模型和确定鉴定指标。【结果】各指标在适宜温度下变异幅度较小,变异系数为3.12%—18.89%,各品种(系)出苗率在85.00%以上,具有较高的生活力,可用于后续耐冷性分析。在低温胁迫下群体内各指标变异幅度增大,为7.14%—108.33%,在恒定低温和昼夜变温下,变幅最大的指标依次为根长和萌发指数。主成分分析将14个低温相关指标和百粒重转换为6个相互独立的综合指标,代表了全部数据74.98%的信息量。利用隶属函数法计算综合耐冷评价值(),并对其进行聚类分析,按照耐冷性强弱将200份陆地棉品种(系)划分为5类,第Ⅰ类群属于强耐冷型共2份,第Ⅱ类群属于耐冷型共42份,第Ⅲ类群属于中耐冷型共69份,第Ⅳ类群属于较敏感型共83份,第Ⅴ类群属于敏感型共4份,其中,新陆中16号为耐冷性最强的品种。采用多元回归分析方法,建立棉花出苗期耐冷性预测模型为=-4.10+0.584+0.4014+0.321+0.225(2=0.92),筛选出4个耐冷性鉴定指标,分别为恒定低温下的棉苗总长、出苗率、干物重和昼夜变温下的萌发率。田间早播试验中各品种(系)的出苗率,与室内试验结果基本一致。【结论】采用恒定低温和昼夜变温处理结合多元统计分析方法对棉花出苗期耐冷性评价是可行的,恒定低温下的棉苗总长、出苗率、干物重和昼夜变温下的萌发率,可作为鉴定指标。
棉花;出苗期;耐冷性;综合评价;指标筛选
0 引言
【研究意义】棉花是重要的天然纤维作物,作为战略物资在国民经济中占有重要地位。新疆已成为中国最重要的棉花生产基地,2018—2021年连续4年新疆棉花总产量占全国80%以上。然而,新疆棉区早春低温危害频发,导致棉花萌发出苗率降低,生育期延迟,最终造成产量及品质下降,早期低温已经成为影响新疆棉花生产的主要环境因素之一[1-2]。因此,了解棉花出苗期耐冷特性,建立耐冷性预测模型、筛选评价指标、鉴定耐冷品种,对生产中抵御早春低温冷害具有重要意义。【前人研究进展】出苗期是棉花遭受低温冷害最多的时期[3-4],种子萌发最低温度为10.5—12.0℃,10.0℃以下不能萌发[5-6],下胚轴伸长并形成维管束需要在16.0℃以上[7]。新疆棉花出苗期面临的温度胁迫通常是“倒春寒”形成的0—15.0℃低温[8],低温已经严重制约棉花生产,选育耐冷品种是解决低温冷害问题最直接有效的方法。针对植物出苗期耐冷性鉴定,国内外研究人员从鉴定方法和指标筛选等方面进行了广泛研究。在玉米萌发期耐冷性研究中,在10℃低温胁迫下测定发芽率、发芽指数等性状,采用主成分分析、聚类分析等统计方法对自交系进行耐冷性鉴评,并划分5个耐冷等级[9]。在粒用高粱中,利用10℃低温胁迫对257份种质材料种子萌发期耐冷性进行研究,筛选出萌发率作为高粱萌发期耐冷性鉴定指标[10]。韩德志等[11]利用158份中欧大豆种质,采用人工低温胁迫处理,通过不同时期出苗率、最终出苗率、相对出苗率及出苗指数进行综合分析,获得出苗期耐冷种质6份。目前,多指标的综合评价在作物出苗期耐冷性鉴定中已得到广泛应用。在棉花出苗期耐冷性鉴定和指标筛选研究中,王俊娟[12]利用恒温0℃冷处理7 d后的子叶展平率作为棉花萌发期耐冷性鉴定指标,对13份棉花种质材料进行耐冷评价。张陇艳等[13]在4℃恒定低温下对53份陆地棉品种(系)进行综合评价,发现胚芽鲜/干重、活力指数可作为萌发期耐冷性鉴定的正向指标,而平均发芽时间和平均发芽速度可作为负向指标。目前对棉花出苗期耐冷性综合评价多采用恒定低温,忽略昼夜温差变化,昼夜变温与恒定温度在作物中存在不同的响应机制。在昼夜变温条件下,小麦幼苗的淀粉酶和葡萄糖-6-磷酸脱氢酶活性具有昼夜周期特性,而恒温条件下生长的小麦幼苗则无此特性[14]。作物耐冷性是一个复杂的数量性状,受诸多因素控制,在不同生育时期和不同冷害类型下会表现出显著的形态发育及生理生化差异[15],因此,探讨适当的评价方法和指标尤为重要。【本研究切入点】前人对棉花出苗期耐冷性鉴定研究多在单一温度下进行,其结果对指导生产中耐冷品种筛选具有局限性,并且在建立评价体系和筛选指标中采用材料份数较少,未见大规模的耐冷鉴定研究。因此,采用恒定低温和昼夜变温2种鉴定方法对200份品种(系)进行耐冷性研究,其结果将更为准确,将为出苗期耐冷品种鉴定和遗传改良提供帮助。【拟解决的关键问题】本研究通过对来源于不同生态区的200份材料为研究对象,结合北疆棉区独特的早春低温气候特点,采用恒定低温和昼夜变温模拟田间低温胁迫,结合田间早播验证试验,测定萌发率、出苗率等指标,深入研究棉花出苗期耐冷性差异,精准筛选耐冷种质,为培育和鉴定耐冷棉花新品种,对解决新疆早春低温造成出苗不齐、苗势不一的瓶颈难题具有现实意义。
1 材料与方法
1.1 试验材料
以来自不同生态区的200份棉花品种(系)为供试材料(电子附表1),其中,中国黄河流域棉区59份、长江流域棉区24份、西北内陆棉区81份和北部特早熟棉区15份,以及国外棉区21份,材料均由中国农业科学院棉花研究所早熟课题组收集和提供,全部材料至少经过3代以上自交纯化。
1.2 试验方法
适温下基础性状测定:脱绒后,每个材料挑选大小一致、饱满的棉花种子,采用培养皿砂培法,每个材料播种3皿,每皿20粒种子,沙子质量含水量为15%,置于适温光照培养室(28℃12 h昼/25℃12 h夜)中生长5 d,逐日统计萌发数,用于计算萌发率和萌发指数;另采用纸钵砂培法,每个纸钵播种15粒,3个纸钵为一组,播种后置于适温光照培养室生长7 d,至子叶完全展平后,测定各材料的出苗率、下胚轴长、根长、棉苗总长和干物重。试验重复3次。
设定低温处理温度:统计分析北疆棉区2009—2020年气象数据发现,在4月5日至12日有日均温10—12℃低温危害,4月18日至20日最低温度从8℃骤然降至4℃,结合棉花种子萌发、下胚轴伸长和出苗所需最低温度条件,设定恒定低温(4℃,12 h昼/12 h夜)和昼夜变温(16℃12 h昼/4℃12 h夜)2种低温处理方法。
筛选低温处理时间:随机选20个品种开展低温处理时间筛选试验(品种编号和名称为:8(晋棉23)、18(陕70)、28(新陆早8号)、38(中棉所24)、48(602186)、58(岱字棉14号)、68(鲁棉研36)、78(中棉所12号)、98(新陆早21号)、108(垦N27-3)、118(新陆早37号)、129(新陆中8号)、143(新陆中30号)、148(新陆中41号)、161(辽锦棉6号)、169(鄂棉16号)、178(渤棉1号)、188(赣棉3号)、198(豫棉2号)、199(新陆早53号))。采用纸钵砂培法,播种后置于适温光照培养室生长2 d。随后转至恒定4℃低温冷室中,分别进行低温处理1、3、5、7和9 d,最后移回至适温光照培养室恢复生长,7 d后测定各材料的出苗率;播种后,将纸钵置于昼夜变温环境中,分别培养10、12、13、14和16 d后转移到适温光照培养室中,恢复生长5 d后,测定各材料的出苗率。每个处理设3个重复。
恒定低温处理:采用纸钵砂培法,播种后置于适温光照培养室生长2 d。随后,将全部材料转至恒定4℃低温冷室中培养5 d,最后移回至光照培养室恢复生长,7 d后测定各材料的出苗率、下胚轴长、根长、棉苗总长和幼苗干物重共5个表型指标。同时分别在冷处理前(0 h)和胁迫处理24 h后,去除种壳置于液氮冷冻保存,用于测定可溶性糖含量和游离氨基酸含量。
昼夜变温处理:采用培养皿砂培法,播种后将培养皿置于昼夜变温环境中,培养20 d,每隔1 d统计萌发数,计算萌发率和萌发指数;采用纸钵砂培培养法,播种后,将纸钵置于昼夜变温环境中,培养13 d后转移到适温光照培养室中,恢复生长5 d后,测定各材料的出苗率、下胚轴长、根长、棉苗总长和幼苗干物重共5个表型指标。试验重复3次。
田间试验:以土壤5 cm地温持续3 d平均达到10℃以上作为适宜早播时期,完全随机区组设计,3次重复,共4个小区,各小区面积为56.0 m2(14.0 m×4.0 m),4.0 m行长,每个品种(系)播种1行,每行播种100粒。2021年和2022年早播时间分别为3月28日和3月27日。调查出苗时间和出苗率,待出苗数目稳定后停止调查。
1.3 指标测定方法
1.3.1 形态指标 萌发率(germination rate,GR,%)=种子萌发数/供试种子数×100%(以露白,即胚根伸出0.5 mm为标准判定种子萌发)
萌发指数(germination index,GI)=ΣG/D,G为第D天对应的萌发数(从第一粒种子萌发开始,至连续3 d没有种子继续萌发为止)。
出苗率(emergence rate,ER,%)=出苗数/供试种子数×100%
下胚轴长、根长、棉苗总长和幼苗干物重4个指标,以不同材料的性状测定值为基础数据,对各指标测定值求平均数获得最终值。同时称取各材料的百粒重。
1.3.2 生理指标 采用蒽酮比色法测定植物组织中可溶性糖含量,采用茚三酮显色法测定游离氨基酸含量,测定方法参照《植物生理生化试验原理和技术》。
1.4 数据处理和分析
为了消除不同种质基础性状间差异,采用单个指标的耐冷系数(cold tolerance coefficient,CTC)对耐冷性进行评价。利用Microsoft Excel 2010对数据进行整理和分析,利用SPSS 19.0软件对数据进行频次分析、主成分分析、相关性分析、隶属函数分析、聚类分析和多元回归分析,采用Origin和Graphpad prism作图,相关指标计算及标准如下:
耐冷系数:CTC=,= 1, 2, 3, …,(1)
式中X和CK为第个指标在低温和适宜温度下的测定值。
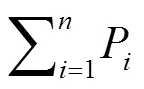
式中,P为第个综合指标贡献率,表示第个指标在所有指标中的重要程度。
各材料单项指标的隶属函数值[(x)]:

式中,x、xmin、xmax分别表示第个指标及第个综合指标的最小值、最大值。
根据因子权重(ω)及隶属函数值[(x)],计算耐冷性度量值(cold tolerance coefficient comprehensive evaluation values,)。
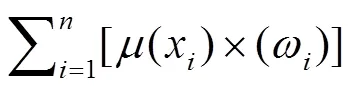
计算主成分权重(W),
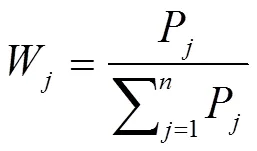
式中,P为各材料第个综合指标的贡献率,为主成分的数目。
计算关联系数(δ)、关联度(γ)和指标权重系数(ω(γ)):
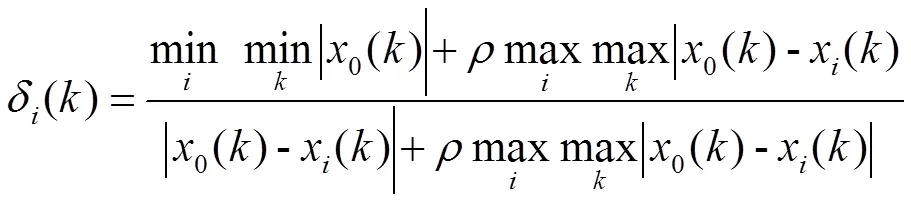
= 1, 2, 3, …,,= 1, 2, 3, …,(6)
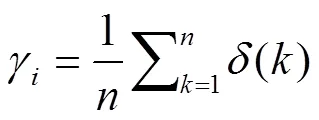
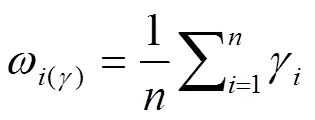
式中,minmin|0()- x()|为二级最小差,maxmax|0()- x()|为二级最大差,为分辨系数(取值0.5)。
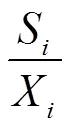
式中,S为第个材料某性状的标准差,X为第个品种某性状的平均值。
2 结果
2.1 不同种质基础性状分析
在适宜温度下,对棉花的10个数量性状进行统计分析,其中出苗率、下胚轴长、根长、棉苗总长和干物重用来衡量材料出苗后生长情况,萌发率和萌发指数反映种子萌发活力,可溶性糖和游离氨基酸含量用来反映低温胁迫下渗透调节物含量变化。对各性状的均值、中位数、变异系数、最值、偏度和峰度进行统计分析(图1),结果表明,只有萌发率在群体内成偏正态分布,其余9个指标为正态分布。供试材料基础萌发率99.05%,在3 d内均可悉数萌发,且出苗率为94.34%,因此,各材料在适宜条件下具有高生活力。各指标在群体内的变异系数介于3.12%—18.89%,萌发指数的变异系数最大,萌发率的变异系数最小,说明各个指标对材料基因型的依赖性不同(表1)。幼苗的下胚轴长、根长、棉苗总长和干物重呈显著正相关关系,百粒重与干物重、可溶性糖和游离氨基酸含量呈正相关关系(图1)。综上可知,在幼苗光形态建成过程中指标间有一定相关性,百粒重对幼苗生长有重要正向调控作用。
2.2 筛选低温处理时间
在恒定低温胁迫下,恢复后,供试材料的平均出苗率随胁迫时间的延长而逐渐降低,变异系数逐渐增大。低温胁迫第5天时供试材料的平均萌发率为40.77%,变异系数为88.04%。当胁迫时间延长至7和9 d时,平均出苗率为降低至2.84%— 2.19%,严重影响棉花出苗。在昼夜变温胁迫下,恢复后的平均出苗率同样随胁迫时间延长呈降低趋势,变异系数呈增加趋势,胁迫13 d时,出苗率的最小值为0.00%,最大值为90.04%,平均值为43.38%。低温处理14和16 d后,出苗率仅约为23.00%(表2)。为保证后续研究中各性状有丰富的变异,确定恒定低温处理时间为5 d,昼夜变温处理时间为13 d。
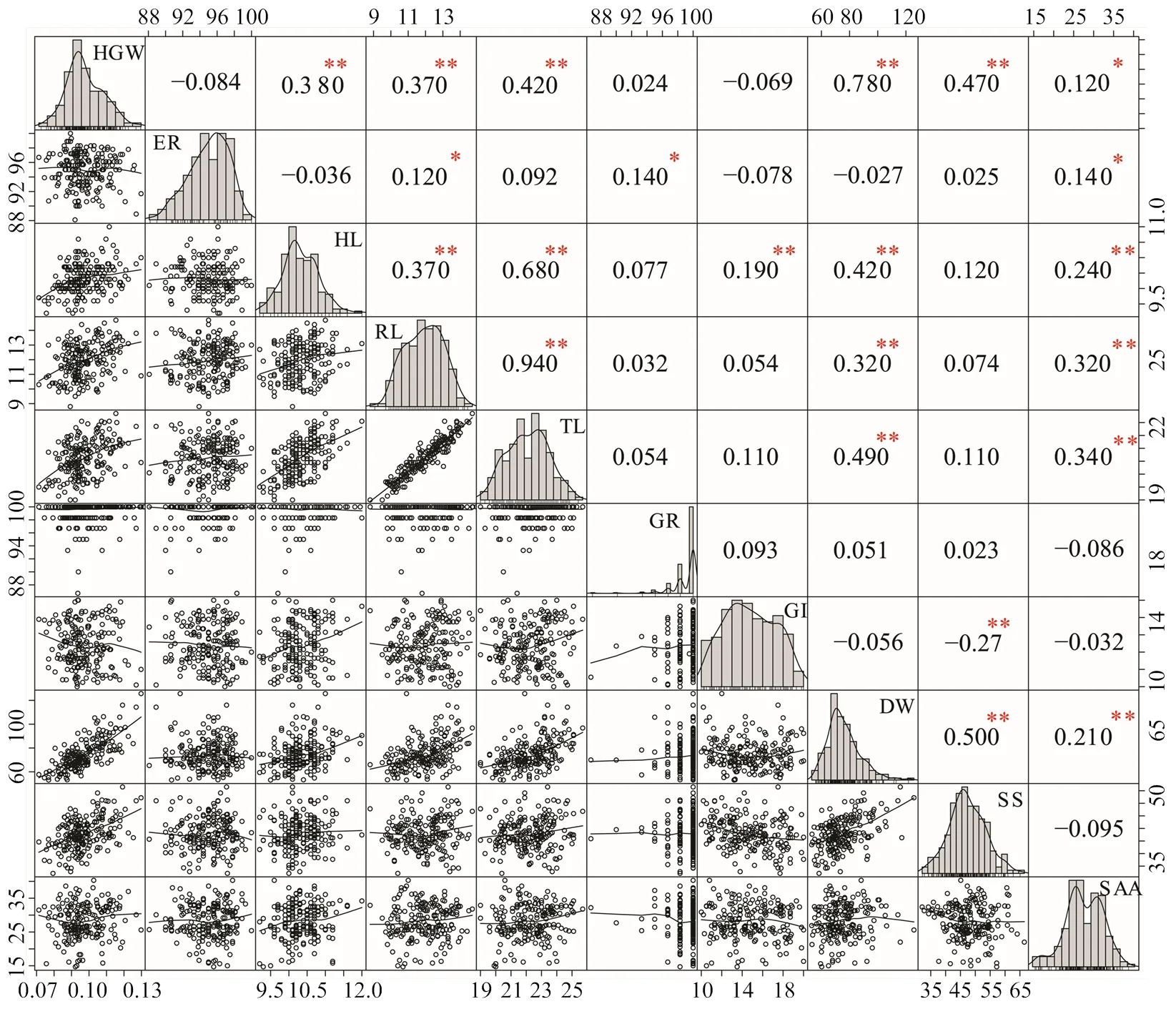
HGW:百粒重;ER:出苗率;HL:下胚轴长度;RL:根长;TL:棉苗总长;GR:萌发率;GI:萌发指数;DW:干物重;SS:可溶性糖含量;SAA:游离氨基酸含量。*和**分别表示在0.05和0.01水平上差异显著。下同

表1 200份棉花品种(系)基础性状的统计学分析

表2 20个棉花品种在低温处理不同时间后出苗率的差异分析
2.3 单项耐冷性指标的差异分析
低温处理后,测定的14个性状均为连续分布,不同低温处理对不同性状的影响程度存在差异。在恒定低温处理5 d,恢复生长7 d后,幼苗根长和低温处理24 h后可溶性糖含量呈偏正态分布,其余下胚轴长、根长、棉苗总长等指标呈正态分布;昼夜变温处理13 d恢复5 d后,根长、棉苗总长和萌发指数呈偏正态分布,其余4个指标呈正态分布。各指标耐冷系数的变异系数介于7.14%—108.33%,其中,昼夜变温下萌发率的变异系数最大,恒定低温下下胚轴长的变异系数最小(表3)。同一区间内各性状耐冷系数分布次数和频率相差较大,CTC值越接近于1,表明耐冷性越强。在CTC大于0.8的指标有恒定低温下的出苗率、下胚轴长、根长、可溶性糖含量等,以及昼夜降温下出苗率、下胚轴长等,其分布频率最高的为可溶性糖含量(99.50%)。CTC值越小且分布频率越高,表明该指标对冷胁迫的敏感度较强,在0<CTC≤0.2区间内,恒定低温下的出苗率,昼夜变温下的出苗率、萌发指数和萌发率指标,分布频率依次为19.00%、1.00%、85.00%和23.00%(表4),表明各指标对冷胁迫反应的敏感程度依次为萌发指数、萌发率、恒定低温下的出苗率和昼夜变温下的出苗率。各指标在低温胁迫后变化趋势不同,在恒定低温处理下的可溶性糖含量在胁迫后含量升高,升幅为23.74%,游离氨基酸含量和表型指标在低温处理后受到显著抑制,降幅在14.13%—86.44%,其中,降幅最大的是昼夜变温下的萌发指数(图2)。不同供试材料间耐冷系数所反映的耐冷性不同,且同一供试材料各指标的耐冷系数值也具有差异较大,表明各指标对低温敏感性不同,因此,只对某一指标的耐冷性分析会忽略指标间复杂关系以及不同的重要性,仍需进一步对多个指标进行综合评价。
2.4 主成分分析
共调查分析出苗期低温相关指标15个,较多性状间相关性会加大研究的复杂度。主成分分析可以对原性状实现综合分析目的。对各指标进行主成分分析,根据特征值>1和主成分载荷矩阵,将具有相同本质的变量归为一类,可将原来各单项性状指标转换为6个新的相互独立的综合指标(用PV1、PV2、PV3、PV4、PV5和PV6表示),且累计贡献率达74.98%,其中,第一主成分的贡献率最大,为22.65%(表5)。在第一主成分中,恒定低温下的棉苗总长、下胚轴长、根长和干物重的特征向量较大,它可能代表棉花出苗期在恒定低温下耐冷性的综合指标。第二主成分中,昼夜变温胁迫下的棉苗总长、根长和下胚轴长等7个指标的系数特征向量较大,表明第二主成分主要表示在昼夜变温胁迫下耐冷性综合指标。第三主成分中,昼夜变温胁迫下的萌发率和萌发指数特征向量相对较大。第四主成分中2种低温胁迫下出苗率的特征向量相对较大。第五主成分中特征向量较大的是昼夜变温下的干物重。第六主成分中百粒重的特征向量较大。同时,在6个主成分中,主要载荷指标与其特征向量值差异较大,说明所测指标在棉花出苗期耐冷表型多样性中均起作用。

表3 不同棉花材料出苗期各耐冷指标的差异分析
CC:恒定低温;DVC:昼夜变温。下同 CC: constant chilling; DVC: diurnal variation of chilling. The same as below
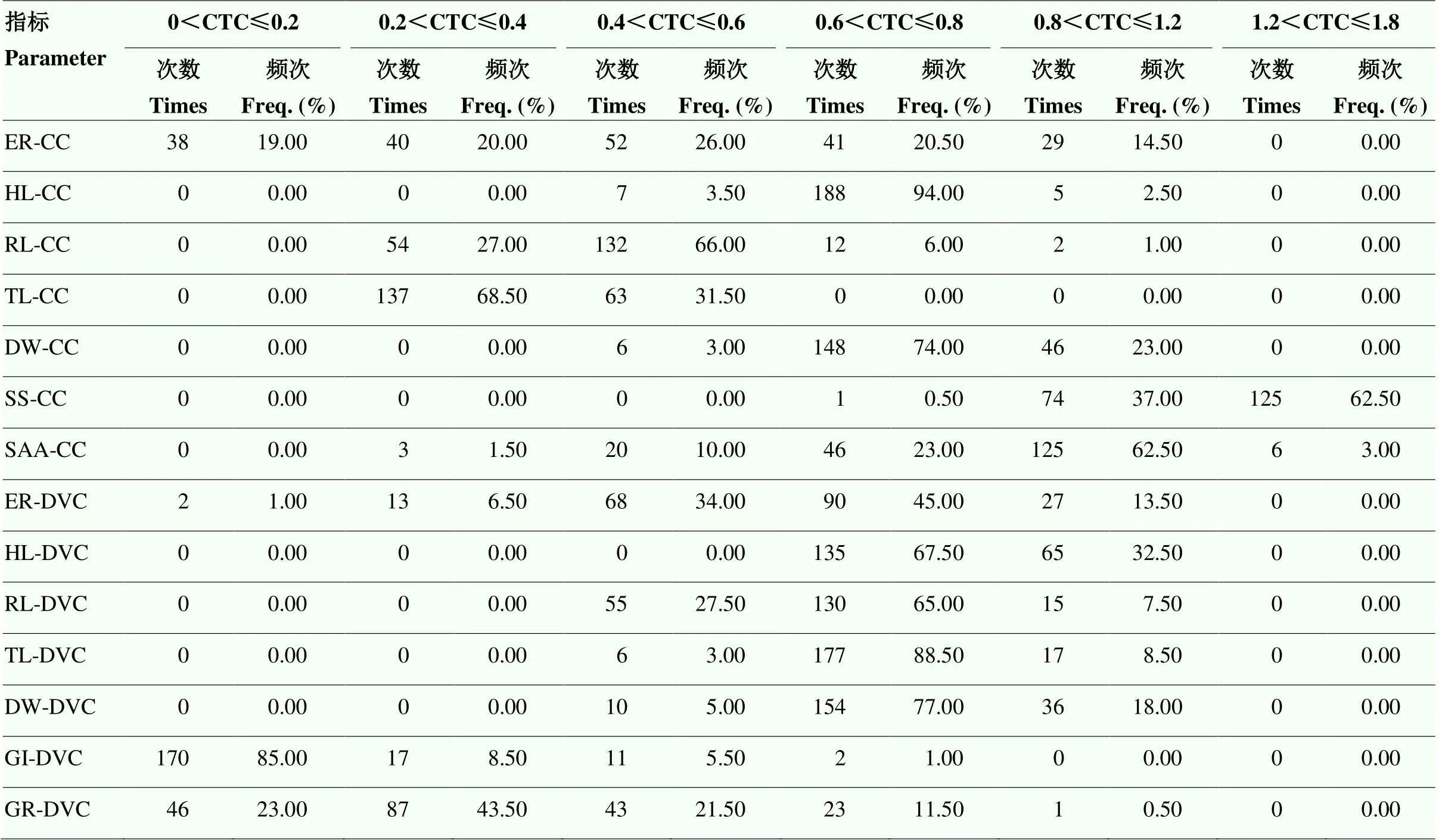
表4 供试棉花材料各性状指标的耐冷系数及其在不同区间的分布
CTC:耐冷系数。下同 CTC: Cold tolerance coefficient. The same as below
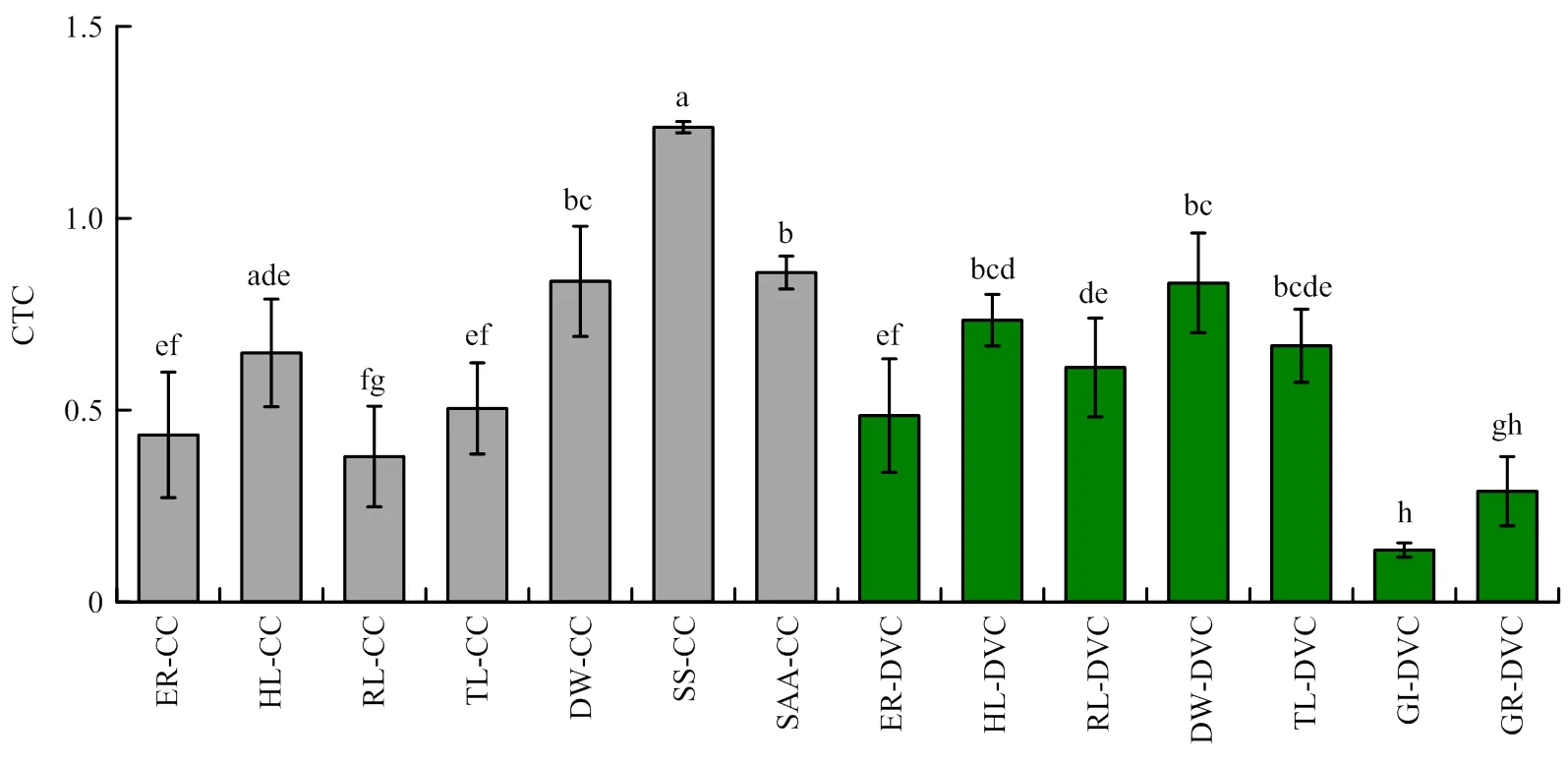
不同小写字母表示差异显著(P<0.05) Different lowercase letters indicate significant differences at P<0.05 level

表5 6个主成分的特征向量、特征值、方差贡献率及累积贡献率
2.5 指标的综合D值评价及耐冷级别划分
依据主成分分析得到6个主成分,计算隶属函数值获得每个材料的值,利用值来评价供试种质耐冷性,值越大耐冷性越强。供试种质资源值介于-3.62—1.78,均值为0.018。根据值对供试材料进行系统聚类分析,对其耐冷性进行等级划分,可将200份棉花种质的耐冷性分为五类(图3-A,电子附表1):第Ⅰ组属于强耐冷型,包含新陆中16号和新陆中12号2份棉花种质,全部来源于西北内陆棉区;第Ⅱ组属于耐冷型,包含新陆中15号、新陆中22号、新陆中21号和新陆早35号等42份,分布于西北内陆棉区的品种(系)数量占此分组的64.29%,来源于国外和长江流域棉区的份数最少,均为7.14%;第Ⅲ组属于中耐冷型,包括新陆早46号、新陆早40号、新陆早16号和新陆早33号等69份,有47.83%的品种(系)来源于西北内陆;第Ⅳ组属于较敏感型,包括新陆早50号、辽棉27号、垦N27-3和辽棉9号等83份,5个棉区的品种(系)数量相差不大,为9.64%—36.14%;第Ⅴ组属于敏感型,包括光叶岱字棉、金垦69-2、银山4号和庄稼汉102号共4份,其中,有分布于西北内陆棉区的2份,国外和长江流域的各1份(图3-B,电子附表1)。由聚类分析结果可知,强耐冷型的种质资源只有2份,占比1.00%,敏感型的种质资源有4份,占比2.00%,剩余大多数为中间类型占到97.00%。从第Ⅰ至第Ⅴ组,耐冷性逐渐降低,各组中分布于西北内陆的品种(系)数量逐渐减少。

A:耐冷性系统聚类图;B:5个分组中品种(系)的地理分布。FR:国外棉区;NEM:北部特早熟;NW:西北内陆;YR:黄河流域;YZR:长江流域
2.6 耐冷指标筛选和耐冷性预测
为建立可用于棉花出苗期耐冷性评价的数学模型,对低温下测定的14个指标和百粒重共15个指标进行多元回归分析。以单项耐冷系数CTC为比较数列,其值为参考数列,得到回归方程。当只有一个变量(TL-CC)时,回归方程判定系数为0.64,检验达到极显著水平,说明模型拟合度较好,回归方程具有一定解释能力(表6)。随着选入方程的自变量依次增加,判定系数逐渐增大并接近1,当自变量为TL-CC、GR-DVC、ER-CC和DW-CC变量时,判定系数为0.92,检验均达到极显著水平,相对应的回归方程能很好评价陆地棉出苗期耐冷性。当再增加一个自变量SS-CC时,判定系数更接近于1,拟合方程更准确,相比表型指标可溶性糖含量测定较复杂,为使鉴定方法简化,选用=-4.10+0.584+0.4014+ 0.321+0.225作为棉花出苗期耐冷性评价模型。同时也表明恒定低温下的棉苗总长、干物重、出苗率和昼夜变温下的萌发率4个指标对低温胁迫更敏感。

表6 棉花品种(系)耐冷性预测模型
1:恒定低温下出苗率;4:恒定低温下的棉苗总长;5:恒定低温下的干物重;6:恒定低温下的可溶性糖含量;14:昼夜变温下的萌发率
1: ER-CC;4: TL-CC;5: DW-CC;6: SS-CC;14: GR-DVC
2.7 耐冷性验证试验
通过田间早播试验对室内鉴定结果进行验证。共挑选30个种质,包括强耐冷型1个(编号为135)、耐冷型4个(编号为138、147、168、144)、中耐冷型13个(编号为122、120、61、95、153、96、141、84、20、83、173、169、9)、较敏感型10个(编号为85、55、140、182、52、60、40、31、41、86)和敏感型2个(编号为165、164),播种后12—13 d开始出苗。品种间出苗率差异较大的时期出现在出苗早期,即播种后15—19 d(4月12日—16日)。强耐冷型和耐冷型品种在2021年和2022年的平均出苗率依次为32.00%和30.45%、46.67%和15.25%,均显著高于中耐冷型、较敏感型和敏感型。其中,强耐冷型品种新陆中16号在2年中出苗率均最高(图4-A)。2个低温条件下获得的综合值,与分别在每种低温胁迫下获得的综合值-CC和-DVC,存在一定相关性。在室内条件下综合值与-DVC值呈显著正相关关系(2=0.97),2021年田间品种(系)的出苗率(ER-2021)与-CC值呈显著正相关关系,2022年田间品种(系)的出苗率(ER-2022)与-DVC值和值呈显著正相关关系(图4-B)。综合值可以用于生产中棉花出苗期耐冷性鉴定,2021年出苗率与-CC值呈极显著正相关关系,可能与2021年4月3—7日连续5 d最低温骤然降至4℃以下有关(图4-C),低温发生形式与室内恒定低温处理方法相似。而在2022年,未出现骤然降温的冷害形式,在播种后15—19 d平均温度在10—13℃(图4-D),低温发生形式与室内昼夜变温处理方式相似。因此,采用恒定低温和昼夜变温相结合的方式,对耐冷品种的筛选更加准确。
3 讨论
3.1 棉花出苗期低温处理方法
选育鉴定耐冷品种是减少冷害的重要途径之一,而耐冷资源的鉴定是培育耐冷品种的基础,因此,如何有效模拟田间低温环境,准确筛选耐冷性种质资源是耐冷品种选育的关键[16]。目前,关于低温对棉花生产的影响以及耐冷种质的筛选已有相关报道,且大多采用恒定低温处理方法。徐建伟等[17]在恒定低温下,以发芽率、发芽指数等为鉴定指标,采用模糊隶属函数值法筛选北疆棉区10年来主栽或大面积推广的棉花品种,获得2份萌芽期强耐冷性材料。高利英等[18]对黄淮棉区不同时期38个代表性棉花品种进行恒定低温18℃、15℃和12℃处理,利用发芽率、发芽指数和活力指数等进行耐冷性鉴定和综合评价,划分耐冷等级。虽然前人对棉花出苗期间耐冷特性进行了有价值的研究,但在处理条件上忽略了田间昼夜温差的影响。因此,确定适合大批量棉花种质耐冷性评价的温度条件是耐冷性鉴定工作有效进行的前提。本研究以200份棉花品种(系)为试验材料,模拟田间播种后可能遭遇的冷害温度,对棉种进行恒定低温和昼夜变温处理,以低温下萌发率、萌发指数、可溶性糖和游离氨基酸含量变化,恢复生长后出苗率、根长、下胚轴长、棉苗总长和干物重,以及百粒重作为评价指标,进行出苗期耐冷性鉴定。恒定低温处理模拟田间播种后骤然降温的危害形式,可造成幼根生长点不可逆损伤,大部分品种(系)在恢复生长后,主根损伤严重,侧根生长旺盛,此低温处理评价了种质芽期的耐冷性。昼夜变温模拟田间播种后日均温为10℃的低温,耐冷型材料在恢复生长后能正常出苗,敏感型材料出现烂芽、烂种,此低温处理评价了种质萌发期的耐冷性。综上所述,本研究使用的鉴定方法与以往恒定温度鉴定相比更接近生产环境,鉴定结果更具有应用意义。

A:田间出苗率;B:室内条件下获得品种(系)的综合值与田间出苗率的相关性分析;D:恒定低温和昼夜变温下各指标的综合值;D-CC:恒定低温下各指标的综合值;D-DVC:昼夜变温下各指标的综合值;ER-2021:供试材料在2021年的出苗率;ER-2022:供试材料在2022年的出苗率;C:2021年田间温度;D:2022年田间温度
3.2 棉花出苗期耐冷性评价指标筛选
植物在逆境条件下,改变自身生长和形态特征如降低植株的高度、鲜重和干重等来适应逆境条件[19-20],通过调整自身生物量分配来维持逆境条件下植株的存活和生长[21-22]。不同品种不同指标对低温胁迫的反应不尽相同,用某一单项指标很难全面准确地反映品种的耐冷实质,采用多个指标对作物的耐冷性进行综合评价更为可靠。以往研究中,棉花出苗期耐冷性鉴定指标,大多为胚芽鲜/干重、发芽率、发芽时间、以及冷处理后出苗率、子叶展平率等表型指标[13, 18],都未涉及百粒重指标。粒重是自然选择和进化的结果,在一定程度上与种子的活力和抗逆性有关[23]。本研究通过统计材料百粒重,发现品种(系)粒重越轻耐冷性越好,这一规律与大豆萌发期耐冷性研究结果一致,百粒重偏小种质耐冷性越显著[11, 24-25],可能是萌发期耐冷相关基因与粒重、籽仁含量等调控相关基因密切连锁导致。本研究在耐冷性综合评价时,增加百粒重作为耐冷鉴定指标,使萌发期耐冷评价更全面准确。
3.3 棉花低温适应与地理分布相关性探讨
棉花原产于热带和亚热带地区,对冷敏感。在长期的人工驯化和自然选择过程中,棉花种植区呈现出生态地理分化,包括一些气温较低的高纬度和高海拔地区[26]。一般认为,西北内陆的品种(系)比黄河、长江流域的棉花更耐冷。通过比较本研究采用的200份材料的地理分布,发现来源于中高纬度区域的种质,如西北内陆棉区的棉花品种(系)多数耐低温,而黄河和长江流域棉区的棉花种质则多数不耐低温。张陇艳等[13]研究中也发现分布于西北内陆的品种新陆中4号等5份材料表现为强耐冷性,同时在拟南芥[27]、水稻[28]、白三叶[29]、黄瓜[30]耐冷性研究中也得到了类似结果,高纬度地区的材料耐冷性更强。因此,推测在棉花生产过程中高纬度地区棉花耐冷性更强,可能是由耐冷基因人工驯化造成的。
4 结论
获得出苗期强耐冷型品种新陆中16号和新陆中12号。恒定低温(4℃)处理5 d后恢复生长7 d;昼夜变温(16℃/4℃)处理13 d后恢复生长5 d的方法,适合作为棉花种质出苗期耐冷性鉴定的条件。恒定低温恢复生长后幼苗的棉苗总长、出苗率、干物重和昼夜变温下的萌发率,可作为棉花出苗期耐冷性快速鉴定的指标。
[1] KARGIOTIDOU A, DELI D, GALANOPOULOU D, TSAFTARIS A, FARMAKI T. Low temperature and light regulate delta 12 fatty acid desaturases (FAD2) at a transcriptional level in cotton (). Journal of Experimental Botany, 2008, 59: 2043-2056.
[2] 范文波, 江煜, 吴普特, 马枫梅, 刘兵. 新疆石河子垦区50年气候变化对棉花种植的影响. 干旱地区农业研究, 2011, 29: 244-248.
FAN W B, JIANG Y, WU P T, MA F M, LIU B. Impacts of climate change on planting proportion of cotton in Shihezi area in the recent 50 years. Agricultural Research in the Arid Areas, 2011, 29: 244-248. (in Chinese)
[3] RUELLAND E, VAULTIER M N, ZACHOWSKI A, VAUGHAN H. Cold signalling and cold acclimation in plants. Advances in botanical research, 2009, 49: 35-150.
[4] JOUYBAN Z, HASANZADE R, SHARAFI S. Chilling stress in plants. International Journal of Agriculture and Crop Sciences, 2013, 5(24): 2961-2968.
[5] LAUTERBACH B, KRIEG D R, JIVIDEN G. Fatty acid composition of lipid fractions in germination cotton as affected by temperature. Proceeding, 1999, 1: 564-565.
[6] 王钰静, 谢磊, 李志博, 魏亦农, 林海荣. 低温胁迫对北疆棉花种子萌发的影响及其耐冷性差异评价. 种子, 2014, 5(5): 74-77.
WANG Y J, XIE L, LI Z B, WEI Y N, LIN H R. Effects of low temperature stress to germination of cotton seeds and evaluation of their cold resistance in Northern Xinjiang. Seed, 2014, 5(5): 74-77. (in Chinese)
[7] 毛树春. 中国棉花栽培学. 上海: 上海科学技术出版社, 2013.
MAO S C, Chinese cotton cultivation. Shanghai: Shanghai Science and Technology Press, 2013. (in Chinese)
[8] 雷斌, 李进, 段留生, 张鹏忠, 李杰, 谭伟明. 种衣剂对低温处理下棉花胚根及幼苗外部形态和超微结构的影响. 中国农业气象, 2017, 38(4): 248-256.
LEI B, LI J, DUAN L S, ZHANG P Z, LI J, TAN W M. Effect of seed coating agents on external morphology and ultrastructure of cotton radicles and seedlings under low temperature treatments. Chinese Journal of Agrometeorology, 2017, 38(4): 248-256. (in Chinese)
[9] 刘杰, 张春宵, 李淑芳, 郑大浩, 梁烜赫, 王宇, 刘文平, 刘学岩, 曹铁华, 李晓辉. 95份玉米自交系萌发期耐冷性鉴评与遗传基础分析. 分子植物育种, 2021, 19(7): 2391-2401.
LIU J, ZHANG C X, LI S F, ZHENG D H, LIANG X H, WANG Y, LIU W P, LIU X Y, CAO T H, LI X H. Evaluation on chilling tolerance of ninety-five inbred lines at maize germination stage and analysis on genetic basis. Molecular Plant Breeding, 2021, 19(7): 2391-2401. (in Chinese)
[10] 徐建霞, 丁延庆, 曹宁, 程斌, 高旭, 邹桂花, 张立异. 粒用高粱种质资源耐冷性分析. 种子, 2021, 40(11): 40-45+60.
XU J X, DING Y Q, CAO N, CHENG B, GAO X, ZOU G H, ZHANG L Y. Analysis of cold tolerance of grain sorghum germplasm resources. Seed, 2021, 40(11): 40-45+60. (in Chinese)
[11] 韩德志, 孙如建, 徐江源, 闫晓飞, 贾鸿昌, 柴燊, 刘章雄, 鲁玉清, 孙宾成, 鹿文成, 邱丽娟.中欧大豆种质资源耐冷性综合评价. 植物遗传资源学报, 2022, 23(5): 1383-1392.
HAN D Z, SUN R J, XU J Y, YAN X F, JIA H C, CHAI S, LIU Z X, LU Y Q, SUN B C, LU W C, QIU L J. Comprehensive Evaluation of Soybean Germplasm Resources Collected from China and Europe under Cold Conditions. Journal of Plant Genetic Resources, 2022, 23(5): 1383-1392. (in Chinese)
[12] 王俊娟. 棉花抗冷性鉴定及相关基因的表达研究[D]. 北京: 中国农业科学院, 2016.
WANG J J. Identification of the chilling resistance of cotton and the expression of cold resistance related genes[D]. Beijing: Chinese Academy of Agricultural Sciences, 2016. (in Chinese)
[13] 张陇艳, 程功敏, 魏恒玲, 王寒涛, 芦建华, 马峙英, 喻树迅. 陆地棉种子萌发期对低温胁迫的响应及耐冷性鉴定. 中国农业科学, 2021, 54(1): 19-33.
ZHANG L Y, CHENG G M, WEI H L, WANG H T, LU J H, MA Z Y, YU S X. Chilling tolerance identification and response to cold stress ofvarieties (lines) during germination stage. Scientia Agricultura Sinica, 2021, 54(1): 19-33. (in Chinese)
[14] 孙存华, 毛健民, 白宝璋, 白岩. 昼夜变温促进小麦幼苗生长的酶学研究. 吉林农业大学学报, 2000, 22(1): 30-33.
SUN C H, MAO J M, BAI B Z, BAI Y. Studies of enzymology on diurnal change of temperature accelerating the rate of wheat seedling growth. Journal of Jilin Agricultural University, 2000, 22(1): 30-33. (in Chinese)
[15] 张鹤, 蒋春姬, 殷冬梅, 董佳乐, 任婧瑶, 赵新华, 钟超, 王晓光, 于海秋. 花生耐冷综合评价体系构建及耐冷种质筛选. 作物学报, 2021, 47(9): 1753-1767.
ZHANG H, JIANG C J, YIN D M, DONG J L, REN J Y, ZHAO X H, ZHONG C, WANG X G, YU H Q. Establishment of comprehensive evaluation system for cold tolerance and screening of cold-tolerance germplasm in peanut. Acta Agronomica Sinica, 2021, 47(9): 1753-1767. (in Chinese)
[16] SHIM J, GANNABAN R B, REYES B G D L, ANGELES-SHIM R B. Identification of novel sources of genetic variation for the improvement of cold germination ability in upland cotton (). Euphytica, 2019, 215(11): 1-16.
[17] 徐建伟, 张晨, 曾晓燕, 张小均, 李志博, 魏亦农. 近十年新疆北疆主栽棉花种子低温萌发能力差异评价. 新疆农业科学, 2017, 54(9): 1569-1578.
XU J W, ZHANG C, ZENG X Y, ZHANG X J, LI Z B, WEI Y N. Differential evaluation of low temperature germination ability of main cotton seeds in northern xinjiang in recent 10 years. Xinjiang Agricultural Sciences, 2017, 54(9): 1569-1578. (in Chinese)
[18] 高利英, 邓永胜, 韩宗福, 孔凡金, 申贵芳, 李汝忠, 尹燕枰. 黄淮棉区棉花品种种子萌发期低温耐受性评价. 棉花学报, 2018, 30(6): 455-463.
GAO L Y, DENG Y S, HAN Z F, KONG F J, SHEN G F, LI R Z, YIN Y P. Evaluation of the low-temperature tolerance of cotton varieties in the Huang-Huai Region during seed germination. Cotton Science, 2018, 30(6): 455-463. (in Chinese)
[19] FALSTER D S, WESTOBY M. Plant height and evolutionary games. Trends in Ecology & Evolution, 2003, 18(7): 337-343.
[20] 夏军, 时晓娟, 郝先哲, 李楠楠, 田雨, 李军宏, 罗宏海. 低温对不同基因型棉种萌发过程中酶活性及激素含量的影响. 植物生理学报, 2019, 55(9): 1291-1305.
XIA J, SHI X J, HAO X Z, LI N N, TIAN Y, LI J H, LUO H H. Effects of low temperature on enzyme activity and hormone content in germination of different genotypes of cotton seeds. Plant Physiology Journal, 2019, 55(9): 1291-1305. (in Chinese)
[21] ZHANG H, ZHAO Y, ZHU J K. Thriving under Stress: How plants balance growth and the stress response. Developmental Cell, 2020, 55(5): 529-543.
[22] Zhu J K. Abiotic stress signaling and responses in plants. Cell, 2016, 167(2): 313-324.
[23] ZHANG W, XU W, ZHANG H, LIU X, CUI X, LI S, SONG L, ZHU Y, CHEN X, CHEN H. Comparative selective signature analysis and high-resolution GWAS reveal a new candidate gene controlling seed weight in soybean. Theoretical and Applied Genetics, 2021, 134(5): 1329-1341.
[24] 于奇, 曹亮, 金喜军, 邹京南, 王孟雪, 张明聪, 任春元, 张玉先. 低温胁迫下褪黑素对大豆种子萌发的影响. 大豆科学, 2019, 38(1): 56-62.
YU Q, CAO L, JIN X J, ZOU J N, WANG M X, ZHANG M C, REN C Y, ZHANG Y X. Effects of melatonin on seed germination of soybean under low temperature stress. Soybean Science, 2019, 38(1): 56-62. (in Chinese)
[25] 盖志佳, 张敬涛, 刘婧琦, 蔡丽君, 杜佳兴, 陈磊. 耐低温大豆品种(系)的筛选与研究. 中国种业, 2018(6): 57-60.
GAI Z J, ZHANG J T, LIU J Q, CAI L J, DU J X, CHEN L. Screening and research on low temperature resistant soybean varieties (lines). China Seed Industry, 2018(6): 57-60. (in Chinese)
[26] 于雅雯, 余国新, 魏敬周. 供给侧改革背景下新疆棉花生产布局空间变化及影响因素分析. 干旱区资源与环境, 2019, 33(5): 74-80.
Yu Y W, Yu G X, Wei J Z. Analysis of spatial distribution and influencing factors of cotton production lay-out in Xinjiang under the backdrop of supply side reform. Journal of Arid Land Resources and Environment, 2019, 33(5): 74-80. (in Chinese)
[27] MARTÍNEZ-BERDEJA A, STITZER M C, TAYLOR M A, OKADA M, EZCURRA E, RUNCIE D E, SCHMITT J. Functional variants of DOG1 control seed chilling responses and variation in seasonal life-history strategies in. Proceedings of the National Academy of Sciences of the USA, 2020, 117: 2526-2534.
[28] LV Y, GUO Z L, LI X K, YE H Y, XIONG L Z. New insights into the genetic basis of natural chilling and cold shock tolerance in rice by genome-wide association analysis. Plant, Cell & Environment, 2016, 39: 556-570.
[29] INOSTROZA L, BHAKTA M, ACUÑA H, VÁSQUEZ C, IBÁÑEZ J, TAPIA G, MEI W, KIRST M, RESENDE M, MUNOZ P. Understanding the complexity of cold tolerance in white clover using temperature gradient locations and a GWAS approach. Plant Genome, 2018, 11: 3835.
[30] 王伟平, 宋子超, 薄凯亮, 董邵云, 魏爽, 苗晗, 李锦斌, 张圣平, 顾兴芳. 黄瓜核心种质幼苗耐低温性评价及GWAS分析.植物遗传资源学报, 2019, 20(6): 1606-1612.
WANG W P, SONG Z C, BO K L, DONG S Y, WEI S, MIAO H, LI J B, ZHANG S P, GU X F. Evaluation and genome-wide association study (GWAS) of low-temperature tolerance at seedling stage in cucumber core germplasm. Journal of Plant Genetic Resources, 2019, 20(6): 1606-1612. (in Chinese)
Construction of A Comprehensive Evaluation System and Screening of Cold Tolerance Indicators for Cold Tolerance of Cotton at Seedling Emergence Stage
SHEN Qian1,2, ZHANG SiPing1, LIU RuiHua1, LIU ShaoDong1, CHEN Jing1, GE ChangWei1, MA HuiJuan1, ZHAO XinHua1, YANG GuoZheng2, SONG MeiZhen1, PANG ChaoYou1
1Institute of Cotton, Chinese Academy of Agricultural Sciences/State Key Laboratory of Cotton Biology, Anyang 455000, Henan;2College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070
【Objective】In this study, the purpose was to comprehensively evaluate the cold tolerance of cotton varieties (lines) at seedling emergence stage, establish a reliable evaluation model, screen and identify indicators, and provide a simple and effective evaluation method for the selection and identification of cold-tolerant varieties in cotton.【Method】200 upland cotton varieties (lines) were used to test hypocotyl length, root length and 100-grain weight, etc. under three treatments of constant chilling (CC), diurnal variation of chilling (DVC) and normal conditions. A combination of integrated cold tolerance coefficient difference analysis, frequency analysis, drop analysis, principal component analysis, cluster analysis, and multiple regression analysis were used to classify their cold tolerance types, establish cold tolerance prediction models, and screen evaluation parameters. 【Result】The variation of each parameters at normal conditions were minor fluctuations ranging from 3.12% to 18.89%. The seedling emergence rate was above 85.00%, which had high viability and could be used for subsequent cold tolerance analysis. The variability of each parameter within the accessions increased under chilling stress, ranging from 7.14%-108.33%, and the most variable parameter were root length under CC condition and germination index under DVC condition. Principal component analysis converted the 14 parameters under chilling stress and 100-grain weight measured into six mutually independent composite indicators, representing 74.98% of the total data information. The comprehensive cold tolerance evaluation value () was calculated by the affiliation function method and then clustering analysis was performed. 200 cotton varieties (lines) were divided into five categories according to their cold tolerance, with 2 of the group Ⅰ being strongly cold tolerant, 42 of the group Ⅱ being cold tolerant, 69 of the group Ⅲ being medium cold tolerant, 83 of the group Ⅳ being more sensitive, and 4 of the group Ⅴ being sensitive, of which Xinluzhong 16 was the most cold-tolerant material. A multiple regression analysis was applied to establish a prediction model for cold tolerance of cotton at seedling emergence as=-4.10+0.584+0.4014+0.321+0.225(2=0.92), and four parameters for cold resistance evaluation were confirmed, namely total length of seedling, emergence rate, and dry weight under CC stress, germination rate under DVC stress. The cold-tolerant varieties (lines) had higher seedling emergence rates of early sowing experiment in the field, which were basically consistent with the results of the indoor results. 【Conclusion】It is feasible to use CC and DVC stress combined with multivariate statistical analysis to evaluate the cold tolerance of cotton at seeding stage, and total length of seedling, emergence rate, and dry weight under CC stress, germination rate under DVC stress can be used as evaluation parameters.
L.; seedling emergence stage; cold tolerance; comprehensive evaluation; parameters screening

10.3864/j.issn.0578-1752.2022.22.002
2022-07-13;
2022-08-15
2021年度第七师胡杨河市财政科技计划(2021C02)、中国农业科学院科技创新工程(CAAS-ASTIP-2020-ICR)
沈倩,E-mail:shenqian429@126.com。通信作者宋美珍,E-mail:songmzccri@163.com。通信作者庞朝友,E-mail:chypang@163.com
(责任编辑 李莉)
