大豆杂种优势利用研究进展*
2022-01-19孙妍妍赵丽梅张春宝
孙妍妍,赵丽梅,张 伟,张春宝
(吉林省农业科学院大豆研究所,长春 130033)

大豆[Glycine max(L.)Merr.]起源于中国,已有五千多年的种植历史,是食用植物性优质蛋白和植物油的重要来源。近年来,我国大豆需求量增长迅猛,但自产供应量却呈负增长趋势。据海关总署公布的大豆进口数据显示,2020年我国进口大豆量再创新高,突破10 032.7万t,同比2019年增加1 181.4万t,增长率为13.3%。究其原因主要是大豆单产较低,种植效益差,农民种植大豆的积极性不高,国内大豆产量难以满足需求。因此,快速大幅度地提高大豆产量,稳定大豆生产,是提高我国大豆自给率的最有效途径之一,也是积极应对复杂国际贸易环境,打好种业翻身仗推进大豆振兴的重要条件。
杂种优势利用是提高作物单产的有效途径之一,已在水稻、玉米、油菜等作物中得到推广应用,取得了良好的经济效益和社会效益。大豆杂种优势利用研究起步较晚,20世纪七、八十年代,美国连续授权了5项大豆杂种优势利用相关专利[1-5],但此后再无相关研究进展。与此同时,我国科研人员对大豆杂种优势利用也展开了技术攻关,并取得了突破性进展。1993年,孙寰等[6]育成了世界上第一个实际可用的大豆细胞质雄性不育系[7],1995年实现“三系”配套,2002年育成世界上首个大豆杂交种“杂交豆1号”,较对照增产21.9%[8]。
经过30余年的系统研发,几代杂交大豆研究人员的不懈努力,国内育成并通过审定杂交大豆品种近40个,大豆杂种优势利用新技术、新方法获授权发明专利、制定技术标准30余项。上述成果奠定了我国在大豆杂种优势利用领域的国际领先地位。文中将从杂交大豆亲本创制、杂交种选育、制种技术和基础研究四个方面论述近年来大豆杂种优势利用研究进展,以期为今后开展强优势杂交种选育、规模化制种技术研发、杂种优势分子机理解析等方面研究提供参考。
1 亲本创制
1.1 细胞质雄性不育系的创制
大豆杂交种的选育,目前主要采用“三系”法,其中综合性状优良、育性稳定性高的不育系是创制强优势杂交种的关键。目前,国内外已有RN 型[9-10]、ZD 型[11]、XXT 型[11]、M 型[12]、N8855[13]、N21566[14]、N23661[15]等多个大豆不育胞质的报道。RN型胞质主要用于北方春大豆区杂交种选育及组合配制,目前育成的RN型不育系及同核异质保持系数量达500余对,其中高不育率、高配合力、高异交率的“三高”不育系40余个。这主要得益于吉林省农业科学院在国内率先建成的以不同规格的隔离网室为基础,以人工驯化蜜蜂为传粉媒介的杂交大豆高效育种平台。通过该平台,不育系转育、提纯扩繁完全摆脱了人工杂交,还实现了规模化恢、保鉴定及异交率鉴定,大幅提升了亲本选育规模。黄淮夏大豆区ZD型胞质应用较多,也已育成了50余对不育系和配套保持系,但尚未有“三高”不育系育成。目前,国内研究单位虽育成了大批细胞质雄性不育系,但由于缺乏规范鉴定标准,不育系鉴定尚存在表型一致性、花粉败育率、育性稳定性等指标不规范统一的问题。随着国内首个大豆细胞质雄性不育系鉴定标准在2020年底发布实施[16],这将为今后科学、规范地开展不育系选育和鉴定提供操作规程,也为品种审定部门开展大豆杂交种规范化审定及不育系植物新品种权保护提供技术支撑。
1.2 恢复系的鉴定与评价
恢复系作为杂交种的父本,其所含恢复基因的恢复能力对杂种F1能否正常结实起决定作用。目前恢复系主要采用测交鉴定法,其步骤主要是通过对其与不育系测交所获的F1代单株进行花粉育性鉴定,若败育率在0~60%[17],则认为其含有恢复基因,可作为恢复系用于杂交组合配制。虽然花粉败育率达到60%以下就可以鉴定为恢复系,但在实际应用中发现测交F1的花粉败育率越低,其恢复能力越强,配制杂交组合F1育性越稳定,这可能是由于恢复系所含恢复基因恢复能力较强或含有多个恢复基因共同作用所致。2014年,张井勇等[18]研发了一套快速鉴定恢复系及其恢复能力的方法。该方法通过待测材料与不同被恢复能力不育系测交,下一生长周期对不同被恢复能力不育系所得F1单株进行花粉育性鉴定,依据花粉败育率快速鉴定待测材料是否为恢复系及其恢复能力,大大加快了恢复系选育进程。
1.3 优异亲本材料的创新
“三系”法虽被广泛的应用于大豆杂交种选育,但由于受特异性基因源的限制,已有大豆材料的利用率仅在20%左右,极大限制了大豆杂种优势潜力的发挥。近年来,研究者针对现有杂交大豆“三系”亲本个别性状的特点,通过“恢×恢”“保×保”“恢×新种质”“保×新种质”等方式,陆续创制出了一批优异的亲本材料,极大地拓宽了杂交大豆亲本的选择和利用范围,为培育强优势大豆杂交种提供了材料补充。
2 杂交种选育
自2002年,世界上第一个大豆杂交种杂交豆1号通过审定以来,在众多育种单位和育种工作者近20年的努力下,先后培育了杂交大豆品种39个,其中春播区审定29个,夏播区审定7个,春、夏播区同时审定3个,见附表。这其中十五至十二五期间共审定13个,十三五期间审定19个,十四五开局之年就已审定了7个。这体现了近年来国内各育种单位在育种规模及杂种优势利用技术上的不断提升。
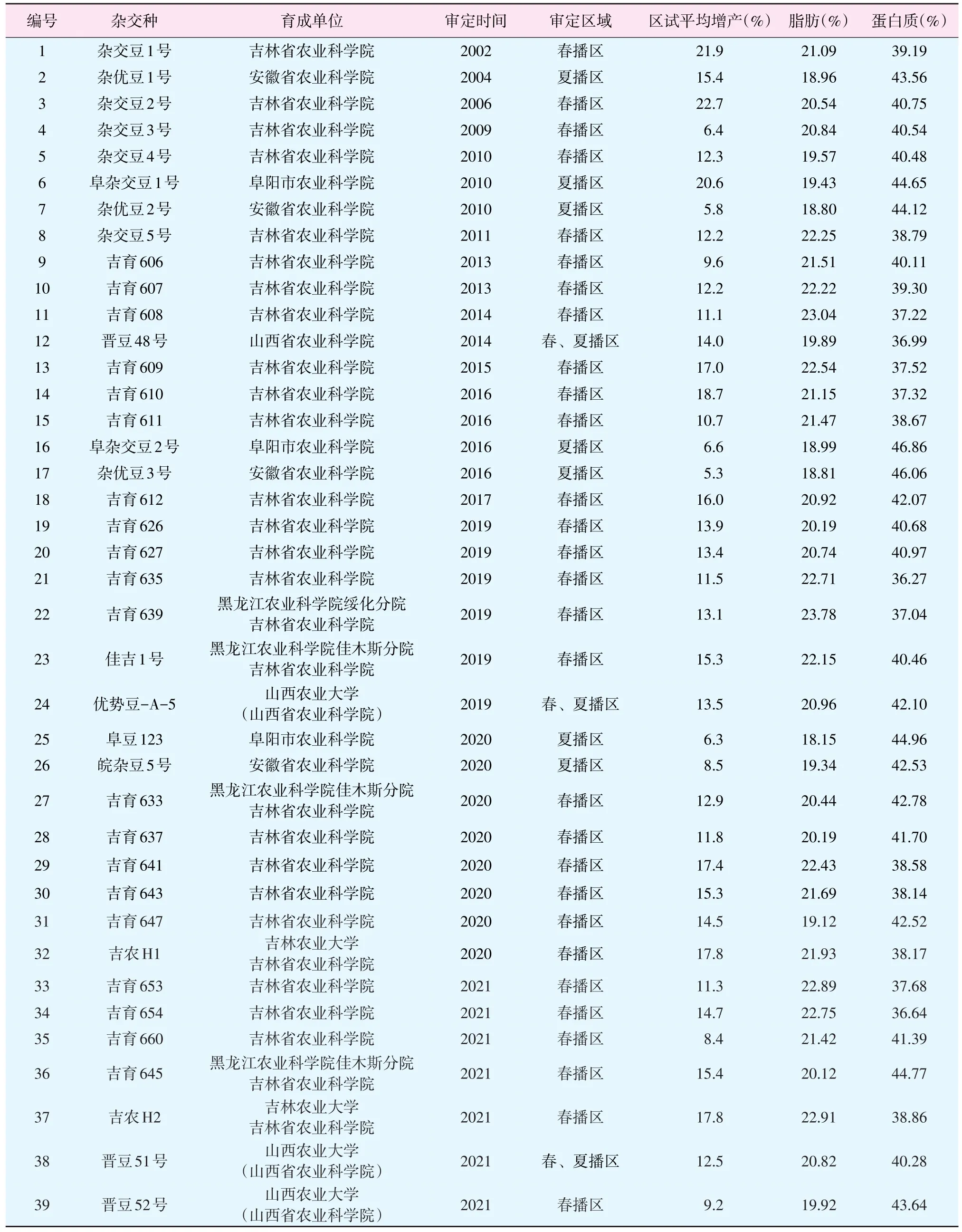
附表 2002—2021年国内审定杂交大豆品种情况
2.1 春大豆杂交种的选育
在东北春播区,吉林省农业科学院通过育种平台的不断完善,目前年均配制600个杂交组合,并进行小批量杂交种生产;结合多年、多点的育性、产量、生育期、农艺性状、抗病虫及适应性鉴定,育成杂交大豆品种28个,其中6个是与黑龙江省农业科学院绥化分院、佳木斯分院和吉林农业大学三家单位联合育成。尤其是在十二五以来,24个春大豆杂交种通过了黑龙江和吉林省品种审定,其中吉育610较对照绥农28增产18.7%,在东北中部地区具有良好的推广前景;吉育635、吉育639、吉育641、吉育643、吉育653、吉育654、佳吉1号、吉农H1、吉农H2 9个杂交种脂肪含量超过21.5%高油大豆标准;适合东北中南部种植的吉育612是国内首个高产双高春大豆杂交种,区试较对照吉育72平均增产16.0%[19]。2021年审定的吉育645为首个高产高蛋白春大豆杂交种,其蛋白质含量高达44.77%,蛋脂合计更达64.89%,区试较对照绥农26平均增产15.4%。上述品种的育成,实现了东北春大豆杂交种产量和品质的协同提升。2018年专家对吉林辽河源的杂交豆5号千亩示范田进行测产,平均产量达3 442.5 kg/hm2,较对照品种吉育47增产15.6%;对吉林范家屯的吉育611的百亩示范田进行测产,平均产量达3 664.5 kg/hm2,较对照品种吉育47增产18.7%。
2.2 夏大豆杂交种的选育
在黄淮夏播区,7个杂交种通过了安徽省品种审定,其中安徽省农业科学院育成4个,阜阳市农业科学院育成3个。2004年审定的杂优豆1号和2010年审定阜杂交豆1号,区试平均增产分别达15.4%和20.6%,但之后育成的5个品种区试增幅仅在5.3%~8.5%,增产幅度与常规品种没有显著差异。究其原因可能是黄淮地区育成亲本数量偏少,亲缘关系过近,没有充分发挥杂种优势潜力所致。山西省农业科学院(山西农业大学)近年来杂交大豆品种选育进展较快,2014年审定了山西省第一个大豆杂交种晋豆48号,并在黄淮地区率先建立了比较完善的杂交大豆育种平台,近年来恢复系鉴定和不育系转育规模显著提升,优势组合配制和鉴定数量大幅增加,相继育成3个杂交大豆品种,其中优势豆-A-5和晋豆51号增产幅度分别达12.1%和12.5%。2020年,在山西运城晋豆52号示范田,专家实收500.25 m2测产,折合产量达4 237.5 kg/hm2。可见依据地缘优势建立适度规模的育种平台,提升亲本创制及杂交组合配制能力,将有效提升黄淮地区杂交种的增产潜力。
3 制种技术研究
3.1 传粉媒介的研究与利用
自实现“三系”配套起,学者们就不断探索杂交大豆异交传粉的最佳方式[20-24]。大豆因其叶腋开花结荚的特点,且花粉粘重由龙骨瓣包裹,无法像玉米那样通过风媒或像水稻那样通过赶粉完成杂交授粉。大豆花具备虫媒花特征,具有蜜腺可分泌少量花蜜[25],这决定了虫媒传粉为其杂交制种的有效方式。20世纪90年代初,由吉林省农业科学院从法国和加拿大引进的苜蓿切叶蜂最早被用于杂交大豆传粉,其传粉效果显著,网室内高异交率不育系利用苜蓿切叶蜂传粉,结实率可达93.5%~100%。然而,国内人工繁殖切叶蜂技术存在瓶颈,1999年起,吉林省农业科学院和吉林省养蜂科学研究所合作,开展了利用蜜蜂为杂交大豆传粉的研究。此后,通过对蜜蜂的定向选育、引诱剂研发[26]、蜂群规模结构[27]、饲喂方法、管理方式等方面的不断改进,实现了人工驯化蜜蜂高效为大豆授粉。同时,研究人员也一直探索在大田开放环境下,依靠野生昆虫进行杂交大豆制种传粉的研究。通过在吉林西部开展野外调查及架设电子监控设备观察访花昆虫种类,明确了切叶蜂、西蜂、隧蜂等为杂交大豆主要传粉昆虫[28]。近年来,为了适应规模化制种需求,王鹏年等[29]通过在制种基地周边穿插种植苜蓿、草木樨等蜜源植物,显著提高了野生传粉昆虫的种类和群体数量,建立了传粉昆虫保护、繁殖和利用技术。
3.2 制种环境的选择
制种区域的选择对不育系能否正常完成异交授粉起关键作用。通过多年试验,研究人员明确了干旱,少雨,7月份降雨量≤100 mm且可灌溉,植被和环境有利于野生传粉昆虫生存、繁殖是杂交大豆制种的最佳区域。通过大量试验,确定了东北西部、内蒙古,西北的山西、陕西、宁夏、甘肃、新疆等地区均适合规模化制种。
3.3 制种技术体系的建立
除了传粉昆虫和制种环境外,亲本异交率的高低、花期是否一致、种植比例及密度均对制种产量产生重要影响。经过多年研究,吉林省农业科学院建立了一套完善的“昆虫—环境—植物”三位一体,综合调控的制种技术体系[22,30-31],赵丽梅等[31]研究表明,该制种体系的应用可使高异交率不育系的结荚率达到90%以上。2019—2020年,在吉林西部的松原、黑水等地连续开展了百亩连片杂交大豆制种试验,通过父母本1∶2行比种植,错期播种确保花期相遇,优化栽培模式及病虫草害防治等技术措施,专家大面积实收测产,在父本行额外收获70~80 kg基础上,杂交种吉育608制种产量达1 653.94 kg/hm2,吉育611制种产量达1 708.05 kg/hm2,两个品种繁殖系数均达到1∶30以上,接近了常规大豆的水平。
近年来,随着国内优势杂交种的不断育成,为适应大豆产业化要求,各育种单位均以本地区的大豆种植模式为依据,加强制种技术研究。山西因其独特地理条件非常适合野生传粉昆虫繁殖,也因此成为较为适宜的制种地区。2018年,在山西省农业科学院东磨支基地杂交组合(SXCMS15A×TH40)制种田,天然昆虫传粉,父母本1∶2种植,专家实收715.95 m2,制种产量达到了1 173.6 kg/hm2。由于该单位育成不育系异交率有待进一步提高,制种潜力仍有较大的提升空间。安徽省农业科学院在黄淮南部地区也开展了小规模的杂交大豆制种工作,但近年来该地区“症青”症严重,制种田花期喷施农药会影响昆虫传粉,不喷施农药点蜂缘蝽等刺吸类害虫的大量繁殖造成“症青”危害,严重影响制种产量。因此,下一步拓展制种区域,依托地理优势向西北适宜制种地区拓展,将是黄淮地区杂交大豆制种规避“症青”风险的有效措施。
4 分子基础研究
我国在大豆杂种优势利用上虽处于国际领先水平,但机理研究方面与玉米、水稻等作物仍有不小差距。近年来,随着分子生物学技术发展及高通量测序技术的广泛应用,基础研究方面也取得了显著进展,主要集中在细胞质雄性不育机理、育性恢复基因及核不育基因定位克隆等方面。
4.1 细胞质雄性不育分子机理研究
在不育机理研究方面,Jiang等[32]和刘海军等[33]分别发现线粒体基因atp9和atp6在不育系和保持系间存在RNA编辑。He等[34]、林春晶等[35]和Lin等[36]分别对不育系和保持系的线粒体和叶绿体基因组进行测序和组装,通过二者间的序列差异,预测了多个可能影响细胞质雄性不育的基因。Ding等[37-39]和Zhang等[40]分别对不育系和保持系进行了miRNA测序分析,发现miR156b、miR2119b、miR169可能参与调控大豆细胞质雄性不育。Li等[41]、Han等[42]和Lin等[43]分别开展了不育系和保持系DNA甲基化差异分析方面的研究。此外,研究人员还开展了不育系和保持系的转录组[44]、环状RNAs[45]、泛靶向代谢组[46]等各类组学分析,获得了一大批差异基因并预测了多个可能与不育有关的调控网络。虽然上述研究在不育机理方面取得一些进展,但至今仍未有大豆细胞质雄性不育基因克隆的报道。目前在水稻、油菜、玉米中均有细胞质雄性不育基因被克隆鉴定,研究人员今后可借鉴成功经验,早日鉴定大豆细胞质雄性不育基因,以期为今后新型不育系的创制提供理论基础。
4.2 育性恢复基因的定位与克隆
育性恢复基因的研究开展较早,多家单位的科研人员对大豆恢复基因(Rf)进行了定位研究。大多数定位结果发现Rf基因在大豆16号染色体上,如赵丽梅等[47]、Wang等[48]和郭凤兰等[49]利用RN型不育系和恢复系构建的F2分离群体为材料进行Rf基因定位,最终将Rf基因定位于16号染色体224.1 kb区间。焦东燕[50]和Dong等[51]分别利用ZD型不育系与恢复系构建的F2分离群体将Rf基因定位在16号染色体标记BARCSOYSSR_16_1064和BARCSOYSSR_16_1082之间。Wang等[52]用M型不育系和恢复系构建的F2群体将Rf基因定位于16号染色体162.4 kb区间内,随后又对Rf候选基因进行了qRT-PCR分析,推断Glyma.16G161900可能为Rf基因。最近,Wang等[53]利用NJCMS1A和NJCMS1C杂交获得的F2群体,通过分子标记定位及候选基因敲除验证,最终NJCMSIC所含Rf基因为16号染色体上一个编码576个氨基酸的PPR蛋白。在大豆其他染色体上,研究人员也通过分子标记定位,获得了新的Rf基因位点[54-57]。今后若能鉴定更多的恢复基因,实现多恢复基因的聚合育种,创制恢复力强、恢复谱广的新型恢复系,这将为强优势杂交种的创制提供重要的材料支撑。
4.3 核不育基因的定位与克隆
随着基于核不育基因的第三代杂交育种系统在玉米、水稻中的研发和应用,大豆核不育基因的研究也日益受到关注。在大豆已知的27种核不育突变体中,有15个基因控制位点实现分子标记定位,包括ms0、ms1-ms9、ms12、msp、msMOS、mst-M和msN[58-79]。MS4是大豆中第一个被克隆的核不育基因[80],其编码一个PHD蛋白,移码突变导致不育。2021年,编码驱动蛋白的MS1基因被国内多个团队克隆。其中Jiang等[81]和Nadeem等[82]相继报道了MS1的一种大片段缺失突变类型,Fang等[83]则报道了MS1的另外两种单碱基缺失和氨基酸突变类型。随后,Yu等[84]也完成了MS6基因克隆,其为一个R2R3类MYB转录因子。上述核不育基因的陆续克隆,为大豆第三代智能杂交育种系统的构建和应用提供重要的基因资源。
此外,研究人员还在其他基础研究方面取得了一些进展,如Zhang等[25]研究发现不育系异交率与其花蜜分泌量相关。Yan等[85]发现农艺性状与异交率的关联分析可有效指导高异交率亲本筛选。李蓉等[86]发现FLS基因可能与异交率有关。Zhang等[87-88]则从转录调控和表观遗传层面探索了大豆杂种优势机理。
5 结语
杂种优势在植物中普遍存在,通过杂种优势的利用已显著提高了水稻、玉米、油菜等作物的产量、品质和抗逆性。多年研究表明,大豆同样存在较强的杂种优势。育成的大豆杂交种中杂交豆2号增产幅度最高,两年区域试验比对照平均增产达22.7%。2006年在吉林省农安县专家测产达到了5 332 kg/hm2,创造了吉林省当时的高产记录。虽然杂交豆2号产量优势显著,亲本配合力较好,但由于其母本JLCMS47A异交率低,导致制种成本过高,很难大规模推广应用。因此,创制高异交率、高配合力的不育系是今后进行杂交种选育的先决条件之一。
近年来吉育609、吉育611、吉育645等一批中早熟春大豆杂交种通过审定,为吉林中东部、黑龙江南部地区提供了品种支撑,但随着东北春大豆种植面积日益北移,适宜东北北部种植的杂交种应用前景将更为广阔。由于目前东北春大豆区育种单位尚缺乏MG000-MG0生育期组的亲本材料,今后侧重点应更多的偏向于极早熟亲本材料的创制和杂交组合的配制,适宜该区域大豆杂交种的育成尚需时日。
大豆杂种优势利用研究初期,由于亲本遗传资源少、遗传差异小,缺少高效的杂种优势预测方法作指导等问题,杂交大豆品种选育和亲本创制存在盲目性。近年来,随着研究人员对大豆杂种优势群理论认识的深入,利用分子标记和高通量测序划分了大豆杂种优势群并初步建立了全基因组预测模型,这为杂交组合的合理配制提供了理论支撑,大幅提高了优势组合的选育效率,我国大豆杂种优势利用也会进入崭新阶段。

