PLINs:参与脂解的脂滴相关蛋白
2022-01-18王宇祥邹亦冲王鑫雨庞永佳
王宇祥,邹亦冲,王鑫雨,刘 捷,庞永佳
(1.东北农业大学动物科学技术学院,哈尔滨 150030;2.农业农村部鸡遗传育种重点实验室,哈尔滨 150030;3.黑龙江省普通高等学校动物遗传育种与繁殖重点实验室,哈尔滨 150030)
白色脂肪组织(White adipose tissue,WAT)是脂肪组织主要类型,其主要功能是控制机体能量平衡,在能量过剩时储存三酰甘油(Triacylglycerol,TAG),在能量匮乏时调动TAG,同时也扮演内分泌和副内分泌器官角色[1]。脂肪组织中含有大量脂肪细胞,调控脂质储存和脂解。脂肪细胞形成分为两个阶段,第一阶段间充质干细胞分化为脂肪祖细胞并进一步形成前脂肪细胞,第二阶段前脂肪细胞经克隆增殖和终末分化,转变为成熟脂肪细胞[2]。成熟脂肪细胞内含有一个大脂滴(Lipid droplets,LDs),脂滴中心是一个中性脂核,脂核内含有大量TAG和胆固醇酯(Cholesteryl ester,CE),外面由磷脂单层包裹,并镶嵌多种蛋白质[3-5]。蛋白质组学研究已确定脂滴中200多种蛋白成分,其中一些成分仅定位于脂滴,其余成分同时定位于脂滴及细胞质或其他细胞器中。脂滴包被蛋白家族(Perilipins,PLINs)是脂滴表面含量最多的蛋白,通过与脂滴表面特异性位点结合,调节脂类储存和水解。截至目前,在哺乳动物细胞中尚未发现缺乏PLINs的脂滴,机体各细胞中储存中性脂质,均表达两个或更多PLINs蛋白家族成员[6],可见,此家族蛋白在机体脂类代谢中具有重要作用。本文从PLINs家族成员发现、结构、表达特征,以及在脂解过程中作用等方面展开综述,旨在为PLINs深入研究提供参考。
1 PLINs发现
1991年,Greenberg在大鼠脂肪细胞脂滴周围发现一种未知蛋白质,命名为Perilipin,同时发现Perilipin具有调节TAG沉积和水解作用[7];随后,Jiang等在脂肪细胞分化过程中发现脂肪分化相关蛋白(Adipose differentiation-related protein,ADRP;或称Adipophilin)[8-9],Díaz等在酵母双杂交筛选中鉴定出47 ku的尾连蛋白(Tail-interacting protein of 47 ku,Tip47)[10];2003年,研究者们又发现一种可诱导脂肪细胞分化的蛋白质:S3-12,其中一部分结构类似Perilipin蛋白[11];2006年,最后一个家族成员—心肌脂滴蛋白(Myocardial LD protein,MLDP)在小鼠体内被发现[12-13]。这5个蛋白含有相似氨基酸序列,且在不同物种之间(如果蝇和小鼠)存在保守的外显子-内含子边界,具有较近亲缘关系,可归类为同一蛋白家族。因此,研究者按照发现顺序将其依次命名为PLIN1-5[14]。PLINs家族成员基本信息见表1[15]。
2 PLINs结构与表达
PLINs家族结构具有高度保守性。在哺乳动物中,PLIN1、PLIN2、PLIN3、PLIN5拥有相同疏水PAT结构域(由前3个蛋白首字母命名)和11聚体重复序列;而PLIN4缺少PAT结构域,但其拥有更多11聚体重复序列,且多肽长度几乎是其他PLIN蛋白3倍(见图1)[15]。
研究表明,PLINs通过11聚体重复序列与脂质结合,形成类似alpha螺旋结构(Amphipathic helices,AHs)[16-17],有利于PLINs与脂滴或微粒结合。在哺乳动物中,PLIN1-5由5个单拷贝基因编码各自PLIN蛋白[18],PLINs氨基酸序列的对比结果表明,PLIN2和PLIN3总体相似度最高[19],在哺乳动物细胞和组织中广泛表达。PLIN5氨基酸序列与PLIN2、PLIN3氨基酸序列在整体上具有相似性,但PLIN5主要在褐色脂肪组织、心肌和骨骼肌中表达,还有一小部分在肝脏组织中表达,并在脂肪分解过程中将脂肪输送到线粒体进行氧化[20]。PLIN1与PLIN2、PLIN3和PLIN5序列相似之处主要在氨基末端,而羧基末端与其他家族成员无明显相似性;PLIN1仅在白色和棕色脂肪组织的脂肪细胞中大量表达,在肾上腺皮质、睾丸和卵巢类固醇生成细胞中表达水平较低[21]。PLIN4氨基酸序列与其他成员差异最大,其11聚体重复序列扩展氨基末端与其他家族成员相似性有限,这些重复序列也可折叠成AHs结构,有利于将新生蛋白特异性地靶向到脂滴[17];PLIN4在白色脂肪组织中表达水平最高,在心肌和骨骼肌中表达水平较低。在亚细胞定位方面,PLIN1和PLIN2主要定位于脂滴,而PLIN3、PLIN4、PLIN5则定位于细胞质或内质网富集区[22-23]。
3 PLINs参与中性脂质的脂解
TAG和CE是脂滴内能量储存的主要成分。脂滴在机体能量匮乏或需脂肪酸(Fatty acid,FA)和胆固醇时调动TAG和CE,利用脂肪酶将其水解为甘油、胆固醇、FA等物质并释放能量。脂肪酶包括脂肪甘油三酯脂肪酶(Adipose triglyceride Lipase,ATGL)、激素敏感型脂肪酶(Hormone-sensitive lipase,HSL)、脂蛋白脂酶(Lipoprotein lipase,LPL)、单酰甘油脂肪酶(Monoacylglycerol lipase,MGL)等[24]。而作为脂滴表面含量最多的蛋白,PLINs通过与脂肪酶互作调控脂滴内中性脂质存储和脂解,从而通过代谢达到为机体储存或提供能量的目的。
3.1 PLIN1
PLIN1对脂滴形成具有双向调节作用。一方面,在基础(饲喂)状态下,PLIN1包被在脂滴表面,充当中性脂质保护屏障,发挥抑制脂解作用。ATGL(又称Patatin-like phospholipase domain containing-2,PNPLA2)是机体脂解反应第一步过程中的关键脂肪酶,其功能发挥需与激活因子α/β水解酶结构域蛋白5(α/βHydrolase domain containingprotein 5,ABHD5)结合。在基础状态下,PLIN1受ABHD5长链酰基辅酶A(LC-CoA)的变构调节后,与之相结合,从而抑制ABHD5与PNPLA2结合[25];同时,PNPLA(Phospholipase domain containing)脂肪酶家族的另外一个成员—PNPLA3(Patatinlike phospholipase domain containing-3)可与PNPLA2竞争结合,促使PNPLA2与ABHD5分离,这些作用限制PNPLA2活性,进而抑制脂解。另一方面,在空腹或锻炼情况下,PLIN1可被蛋白激酶A(Protein kinase A,PKA)高度磷酸化而有利于脂肪酶接近脂滴表面,促进脂解作用。在此状态下,肾上腺素(Adrenaline)和去甲肾上腺素(Norepinephrine,NE)与脂肪细胞膜上的β-肾上腺素受体(βadrenergic receptors,β-AR)结合,启动G蛋白介导的信号通路,激活腺苷酸环化酶(Adenylate cyclase,AC),增加细胞内的环磷酸腺苷(Cyclic adenosine monophosphate,cAMP)水平。cAMP升高触发四聚体PKA调控亚基释放,进而激活催化亚基[26]。随后,PKA磷酸化脂解复合物大部分组分。在受刺激的脂肪细胞中,磷酸化程度最高的脂滴相关蛋白即PLIN1[27]。PLIN1磷酸化释放ABHD5,使其与PNPLA2相互作用,水解TAG形成二酰甘油(Diacylglycerol,DAG);PKA还可磷酸化HSL促进其自身转运到脂滴并与PLIN1相互作用,水解DAG形 成 单酰甘油(Monoacylglycerol,MAG);MAG再被MGL进一步水解为FA和甘油,完成脂解(见图2)[28]。PLIN1对脂滴融合也具有一定促进作用。研究表明,PLIN1可与Fsp27(Fat-specific protein 27)的CIDE-N结构域相互作用,通过改变Fsp27构象促进大脂滴形成[29]。此外,PLIN1还参与机体能量代谢,对肥胖、炎症反应、脂肪营养不良等疾病发生和发展具有重要影响[30]。
3.2 PLIN2
PLIN2最初被命名为脂肪分化相关蛋白,说明PLIN2在脂肪细胞分化早期具有诱导分化作用。PLIN2存在于前脂肪细胞中,在分化为成熟脂肪细胞过程中被PLIN1取代,且PLIN2仅与脂滴特异性结合,而在脂滴脂解或缺失情况下PLIN2迅速降解。Gao等研究发现,过表达PLIN2可诱导COS-7细胞和成纤维细胞中脂滴积聚[31],而敲除Plin2基因的小鼠对饮食诱导的肥胖、脂肪肝和酒精诱导的脂肪变性具有抵抗力,表明PLIN2参与脂质积累调控,这种作用在肝脏中尤为明显[32]。Nocetti等研究发现,长期饲喂高脂饲料导致小鼠患胰岛素抵抗和非酒精性脂肪性肝病,同时小鼠肝细胞脂滴中PLIN2表达明显增加[33]。研究表明PLIN2与肝脏中脂滴形成密切相关。此外,PLIN2也具有调节脂解的作用,但不受PKA激活影响[34]。
3.3 PLIN3
哺乳动物PLIN3和PLIN2具有相似氨基酸序列,且广泛分布于各种组织中。Wolins等研究表明,PLIN3是第一个具有在细胞质和脂滴表面之间转移能力的PLINs家族蛋白[35]。现阶段对于PLIN3功能知之甚少,尚未有Plin3基因敲除小鼠模型的报道。但Carr等研究表明,当使用反义寡核苷酸抑制PLIN3在小鼠体内表达时,肝脏和血清中TAG水平降低[36];当抑制PLIN3在HeLa细胞中表达时,TAG储存减少,而FA生成未受影响[37],表明PLIN3在储存TAG方面具有一定作用。但也有研究表明,抑制PLIN3在小鼠AML12肝癌细胞中表达并未减少TAG储存,仅是脂滴变小并被PLIN2包裹;而将PLIN2和PLIN3表达均抑制后,TAG储存减少,细胞内的脂滴数量更少,但体积更大[38]。表明PLINs可能还具有表面活性物质的性质,当其不包被在脂滴表面时,促进脂滴通过融合减小表面积。
3.4 PLIN4
PLIN4是PLINs中最特殊的蛋白,拥有较其他PLINs成员更长的AHs区,该结构在体外已被证明可直接与TAG结合[39]。在细胞中,PLIN4可修复由于卵磷脂(Phosphatidylcholine,PC)生物合成中限速酶CCTα耗尽,细胞中PC含量下降而导致的脂滴形态缺陷。结果表明,PLIN4是直接与脂滴中性脂质核心结合而不是磷脂,从而作为脂滴表面活性物质覆盖于脂滴表面发挥作用[39]。Nimura等研究发现PLIN4还参与脂肪细胞分化。在人骨髓间充质干细胞(HMSCs)分化后,PLIN4出现在脂肪细胞中;且在人类脂肪细胞分化早期阶段,PLIN2表达短暂增加,但在分化后期则被PLIN1和PLIN4取代[40]。
3.5 PLIN5
PLIN5已被证明可参与调解氧化组织中的基础脂解[41]。Kuramoto等建立Plin5基因敲除小鼠,并报道PLIN5在为肌肉细胞线粒体提供能量方面的核心作用。在这项研究中,研究者在高倍电子显微镜下发现,敲除Plin5基因小鼠的心肌中未观察到脂滴,而禁食后小鼠的比目鱼肌肌管内可见少量脂滴;敲除Plin5基因小鼠的身体状况正常,但对耐力运动不耐受[42]。这些数据表明,肌肉中脂滴作为FA储存库和分配者发挥供能作用。
PLIN5还具有促进脂滴和线粒体耦联的作用。研究者通过脂多糖(Lipopolysaccharide,LPS)激活小鼠干细胞,并提取脂滴观察脂滴对大肠杆菌细胞的影响,结果发现PLIN5在LPS刺激后表达水平下降。这是因为线粒体是机体先天免疫细胞器,脂滴接触线粒体可为线粒体提供FA,促进氧化磷酸化;但在线粒体免疫激活情况下,先天免疫细胞更需要糖酵解而非氧化磷酸化;因此细胞下调PLIN5表达量促进脂滴和线粒体解耦联,减少氧化磷酸化。值得关注的是,在此过程中细胞PLIN2表达水平上升,暗示PLIN2与免疫功能相关[43-44]。
4 PLINs参与自噬脂解
在脂肪细胞中,PLINs通过脂肪酶和脂滴调节蛋白相互作用调控脂解,脂肪酶参与动员脂肪TAG的储存和脂解[45]。ATGL和HSL等脂肪酶缺失、敲除或药物抑制会降低大部分脂肪酶活性,并严重影响FA在脂质水解中的释放,影响脂质代谢循环[46]。然而,在非脂肪组织中,其他中性脂肪酶也可对储存在脂滴中的TAG分解代谢,这表明可能存在不同分子机制动员脂肪细胞内TAG储存和脂解[47],如自噬脂解即通过溶酶体降解中性脂质达到调节脂质代谢作用。自噬最初被认为是饥饿期间蛋白质周转和氨基酸循环的途径,最新研究发现自噬还有助于脂质代谢和脂滴分解代谢[48]。到目前为止,已确定两种自噬过程类型参与脂滴生成和脂解,分别是巨型细胞自噬(Macrolipophagy)和分子伴侣介导的自噬(Chaperone-mediated autophagy,CMA)。这两种类型的自噬均需识别脂滴并为脂滴表面提供一个动态界面,该动态界面含有与脂类代谢和运输有关的蛋白质,以促进自噬蛋白在自噬过程中与溶酶体上相关组件对接[49-50]。鉴于PLINs在脂滴上的丰富性、对脂滴相对特异性定位及其在协调“经典”脂解过程中的重要作用,可合理推测PLINs在自噬脂解调节中也发挥重要作用。
4.1 PLINs与巨型细胞自噬
巨型细胞自噬首先是细胞质中被称为吞噬体或隔离膜的膜池可将自噬目标隔离成一个双膜自噬小体,自噬小体通过其外膜与溶酶体限制膜融合,将隔离的自噬目标转移到溶酶体内,完成降解[51]。脂滴被识别为自噬目标并被自噬小体隔离的机制尚不清楚,但小GTPase家族的Rab7(Ras-associated binding)与脂滴表面的结合被认为是启动脂肪细胞和肝细胞“吞脂”作用的早期步骤[52]。在培养的3T3-L1脂肪细胞中,Rab7与PLIN1包被的脂滴在非刺激(基础)条件下结合较少,但在β-肾上腺素能受体刺激下Rab7与PLIN1包被的脂滴结合增加,随后激活PKA和PLIN1磷酸化。Rab7与脂滴的这种募集不是通过与PLIN1相互作用介导。相反,PLIN1敲除增加Rab7和溶酶体结合。这些数据表明,未磷酸化的PLIN1阻断Rab7与脂滴对接,因此在抑制脂质体吞噬方面起保护作用,而磷酸化诱导的PLIN1构象改变使Rab7可重新聚集到脂滴中,使脂滴与溶酶体结合[53],促进脂解。PLIN2和PLIN5也可保护储存的中性脂质免受胞液脂肪酶影响。Tsai等研究发现,过表达PLIN2可保护肝脏脂滴免受自噬脂解,而小鼠肝脏中缺乏PLIN2,TAG储存量比野生型小鼠少[54]。
4.2 PLINs与分子伴侣介导的自噬
与巨型细胞自噬不同,分子伴侣介导的自噬(CMA)是细胞器(包括脂滴)普遍循环过程,其作用是通过CMA受体溶酶体相关膜蛋白2A(Lysosome-associated membrane protein type 2A,LAMP-2A)降解运输到溶酶体膜的特定蛋白来实现。CMA底物蛋白中含有一个五肽基序KFERQ,该基序选择性地被细胞质中70 ku热休克同源蛋白(Heat shock cognate protein of 70 ku,hsc70)识别并结合,再与L-2A相互作用,将蛋白输入溶酶体中降解[55]。最近研究表明,在营养供应有限条件下,分子伴侣介导的自噬是从脂滴中去除PLIN2和PLIN3的主要机制;小鼠PLIN2和PLIN3分别含有与CMA相关的SLKVQ和LDRLQ五肽序列,当PLIN2五肽序列突变为SLKAA时,hsc70与脂滴上PLIN2的结合几乎被消除[55-56];突变的PLIN2随后滞留在脂滴上,通过减少ATGL等脂肪酶与脂滴关联来减弱“经典”脂解作用,同时也减少巨型细胞自噬引起的多种蛋白质受体在脂滴上募集,从而减少自噬脂解。相反,通过CMA从脂滴中去除PLIN2和PLIN3可促进“经典”脂解和自噬脂解,此过程如图3所示[57]。
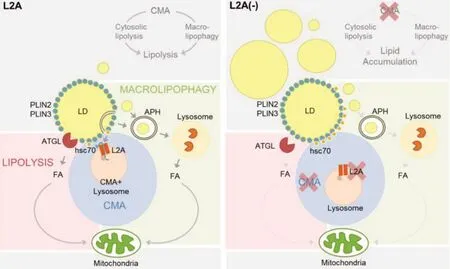
图3 CMA通过降解PLIN2和PLIN3调控脂解[57]Fig.3 CMA regulates lipolysis by degrading PLIN2 and PLIN3
5 展望
近30年研究已揭开PLINs调控脂类物质生成和分解机制的面纱,但大多数调控脂解相关机制研究均集中在PLIN1,对于家族中其他成员研究较少。深入了解PLINs其他成员功能,有利于更好理解非脂肪脂滴在全身能量稳态和代谢性疾病中作用,有助于解决肥胖、脂肪过度沉积、胰岛素抵抗和免疫性疾病等难题。PLINs在细胞脂质稳态中发挥关键作用,其家族成员在储存脂质方面的功能较复杂,涉及多种分子机制和多个细胞内位点。此外,PLINs在不同物种上研究进展不一致,如在家禽脂质代谢中作用机制研究刚刚起步[58-59],深入探究PLINs在家禽脂质代谢过程中功能,有助于控制家禽体内脂肪过度蓄积,推动家禽业向经济、绿色方向发展。
