小麦三雌蕊性状的遗传分析与连锁标记开发
2022-01-14侯光生闫贵云乔麟轶常利芳张晓军畅志坚
侯光生,闫贵云,李 欣,乔麟轶,常利芳,张晓军,畅志坚
(山西农业大学 农学院/作物遗传与分子改良山西省重点实验室,山西农业大学 农学院,山西太谷 030801)
小麦(TritiumaestivumL.,2n = 6x = 42,AABBDD) 是中国三大粮食作物之一,全国40%以上的人口以小麦作为主食。小麦的安全生产与产量提高,对促进中国粮食安全和保障市场需求具有重要影响[1]。穗粒数是影响小麦产量的三大要素之一,一直以来都是遗传育种学家研究的焦点[2]。正常情况下,普通小麦的每一朵小花之内都包含1个雌蕊和3个雄蕊,最终发育成1粒种子[3]。陈济世等[4]最早报道了1种生有3个雌蕊的小麦材料,并将之命名为“三粒小麦”,其主要特点是每朵小花都包含3个雌蕊和3个雄蕊,比正常小麦多出2个雌蕊,且每朵小花都可以发育成3粒种子。
40余年来,国内外已对小麦三雌蕊性状进行了大量研究。王宏霞等[5]通过SSR标记分析三雌蕊性状的可能来源。Peng[6-7]等证实了三雌蕊性状是受1对显性核质基因控制,为三雌蕊性状的遗传分析和染色体定位奠定了基础。Yang等[8]对三雌蕊突变体中的差异表达基因进行了鉴定,Wei等[9]分析了可能与三雌蕊性状相关联的WAG-2D基因的功能,为三雌蕊基因克隆提供了有用的信息。Watanabe等[10]将三雌蕊突变体与具有蓝粒糊粉层基因的品种杂交,对三雌蕊性状的发育起源进行了研究。Peng等[7]将三雌蕊基因定位在小麦4B染色体上。Wang等[11]利用中国春2D缺失系将控制小麦三雌蕊性状的基因定位到了染色体2DL的末端区域。但目前对于小麦三雌蕊性状基因的克隆和三雌蕊性状形成原因及其遗传机理尚无明确结论,在生产上的应用也无较大进展。
CH257为山西农业大学农学院利用‘三粒小麦’与普通小麦品系‘石优20’回交3次后选育的一个三雌蕊材料,本研究通过对CH257与普通小麦中国春构建的后代群体进行遗传分析,并利用90K SNP芯片扫描结合分离群体分组分析法(bulked segrgant analysis,BSA)进行基因定位和连锁标记开发,以期为合理有效地利用三雌蕊小麦种质提供理论依据。
1 材料和方法
1.1 供试材料
本实验使用组合“CH257×中国春”的F1、F2、BC1及F3进行三雌蕊性状的遗传分析,CH257来源于组合‘三粒小麦/石优20*3’的F5选系。中国春(Chinese Spring)小麦种子由电子科技大学生命科学学院杨足君教授惠赠。其他实验材料均来自山西农业大学农学院。
1.2 遗传群体构建
将CH257与中国春进行正反杂交构建F1群体,F1世代单株套袋自交,收获种子后播种构建F2分离群体,F2群体单株编号挂牌后,于4~5叶期每株取5 cm长叶片置于2 mL离心管中,采用CTAB法[12]提取基因组DNA。收获后每株取40粒种子播种于温室,构建F2:3群体,用于推测F2代基因型。
1.3 遗传群体表型调查
田间性状统计数据收集可以在小麦的开花期或成熟期(4月底或5月初)两个时期进行。在开花期进行数据的统计,如果有3个雌蕊在一朵小花里面则被认为是三雌蕊性状,如果只有1个雌蕊和3个雄蕊在一朵小花里面,则被认为是正常雌蕊,即正常小麦性状。
1.4 BSA法混池及SNP芯片扫描
根据F2:3单株表型结果,在“CH257×中国春”组合中选取三雌蕊和单雌蕊各40个F2单株,按照BSA法[13]分别取每个F2单株等量DNA组建三雌蕊和单雌蕊DNA混合池,利用illumina Wheat 90K SNP芯片分别对CH257、‘中国春’、三雌蕊混合池和单雌蕊混合池进行芯片扫描,委托中玉金(北京)生物技术股份有限公司进行SNP分型。扫描结果使用Microsoft Excel 2016进行数据分析[14]。
1.5 连锁标记开发
根据SNP分析结果,确定相关基因所在染色体区段。利用在NCBI数据库(https://www.ncbi.nlm.nih. gov/genome/?term=wheat)中检测中国春基因组(IWGSC v1.0)数据,利用SSR Hunter 1.3检索该序列片段中的SSR,提取所有SSR序列两侧各300 bp序列,利用Primer Premier 6.0设计引物(表1),Tm值为55 ℃,引物长度为18~22 bp。设计的引物在PCR仪(C1000型,美国伯乐公司)进行扩增,扩增产物加入2 μL 6×Loading Buffer,混匀后采用8%的非变性聚丙烯酰胺凝胶电泳分离(220 V恒压),银染法染色,显影后,在胶片观察灯上观察电泳结果,照相并对F2群体进行条带统计。

表1 16个在小麦CH257和‘中国春’上有扩增产物的SSR引物
2 结果与分析
2.1 三雌蕊基因的遗传分析
利用CH257与‘中国春’进行正交构建F1群体,其F1再与双亲进行回交,并对F1、BC1F1和F2群体的性状进行调查分析,结果如表2。其中,F1群体共35个单株,且都为三雌蕊性状;BC1(CH257/中国春//中国春)群体共126株,65株为三雌蕊,61株为单雌蕊,卡方检验符合1∶1;BC1(CH257/中国春//CH257)群体共85株,全部为三雌蕊;F2代群体共312株,224株为三雌蕊,88株为单雌蕊,卡方检验结果表明三雌蕊:单雌蕊符合3∶1比例。以上遗传结果分析表明在CH257中存在一个显性单基因位点控制小麦三雌蕊发育的性状。
2.2 三雌蕊基因SNP芯片分析
运用90K SNP基因芯片分别对‘CH257/中国春’组合两个极端混池以及双亲进行全基因组检测,在两个混池及双亲间均表现差异的SNP进行比对,获得两个混池和两个亲本间共同表现差异的SNP共有557个,根据90K SNP基因芯片遗传整合图谱信息,将表现差异的位点整合到小麦的21条染色体上。由图1,A 可知,3D、4D、6D、7D四条染色体上SNP位点差异不明显;2D染色体上差异位点分布多达247个,约占总SNP数量的44%;3A、5A、6B三条染色体上均分布着12个差异位点;2A染色体上分布32个位点,约占总SNP数量的6%;2B染色体上分布着25个位点,约占总SNP数量的4%;其余的SNP差异位点分布在其他染色体上。另外,由图1,B初步推测控制三雌蕊性状的基因位于2D染色体的长臂上。
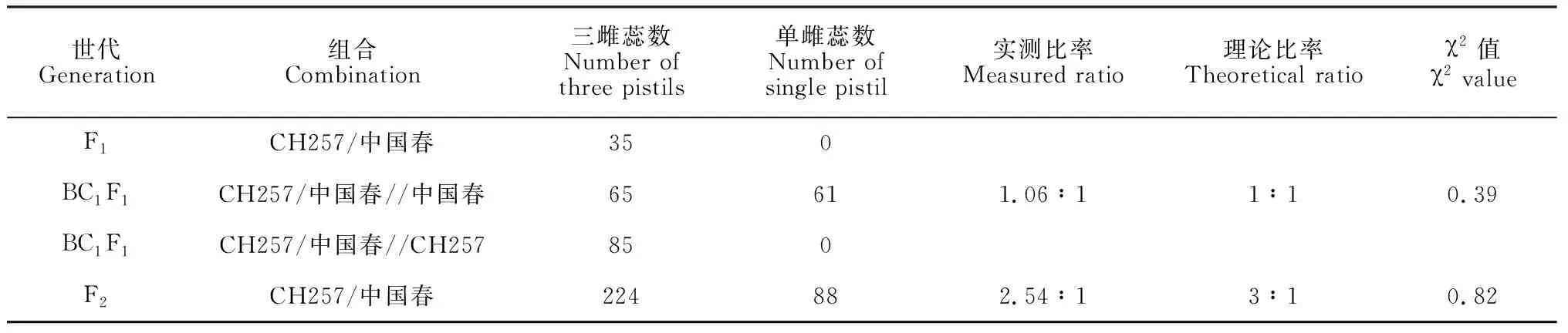
表2 遗传群体性状分析
2.3 连锁标记的筛选
根据SNP芯片检测结果,在2D染色体上设计了48对引物,设计的引物先在亲本CH257和‘中国春’间筛选,共筛选出亲本间多态性引物16对;用这16对引物扩增11株三雌蕊和11株单雌蕊组成的小群体,获得5对连锁的标记,分别为2DL07、2DL17、2DL22、2DL25和2DL38。
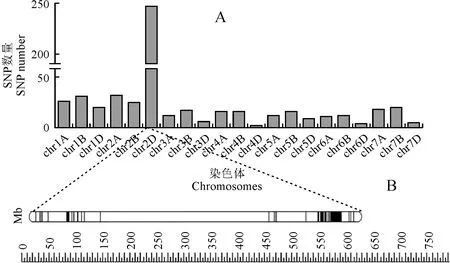
图1 SNP差异位点分布位置(A)以及SNP位点在2D染色体上的分布(B)Fig.1 The physical distribution of SNP difference sites (A) and SNP sites are distributed on the chromosome 2D (B)
利用筛选出的5个多态性标记对F2群体的312个单株进行了PCR扩增(图2),与三雌蕊亲本CH257带型相同的个体基因型记为“A”,二者的杂合带型记为“H”,与单雌蕊亲本‘中国春’带型相同的个体基因型记为“B”。读带结果(表3)显示, 5个标记的基因型数量分离比例均符合1∶2∶1。根据小群体筛选结果推测,上述5个标记与CH257中的三雌蕊控制基因Pis-CH257均存在连锁关系。
2.4 遗传图谱构建
利用5个连锁标记检测全部F2群体,根据分子标记检测结果和田间表型鉴定结果,使用Joinmap4.0软件计算连锁距离,最终将CH257中控制雌蕊发育的基因Pis-CH257定位于2DL染色体2DL22-2DL25之间,2DL22和2DL25与Pis-CH257的遗传距离分别为1.1cM和2.3cM。使用Mapchart2.3绘制遗传连锁图谱(图3,A),通过查询了小麦2D染色体的长度和5个标记的物理位置和已有报道的标记的物理位置(图3,B)绘制了Pis-CH257的物理图谱[15](图3,C)。
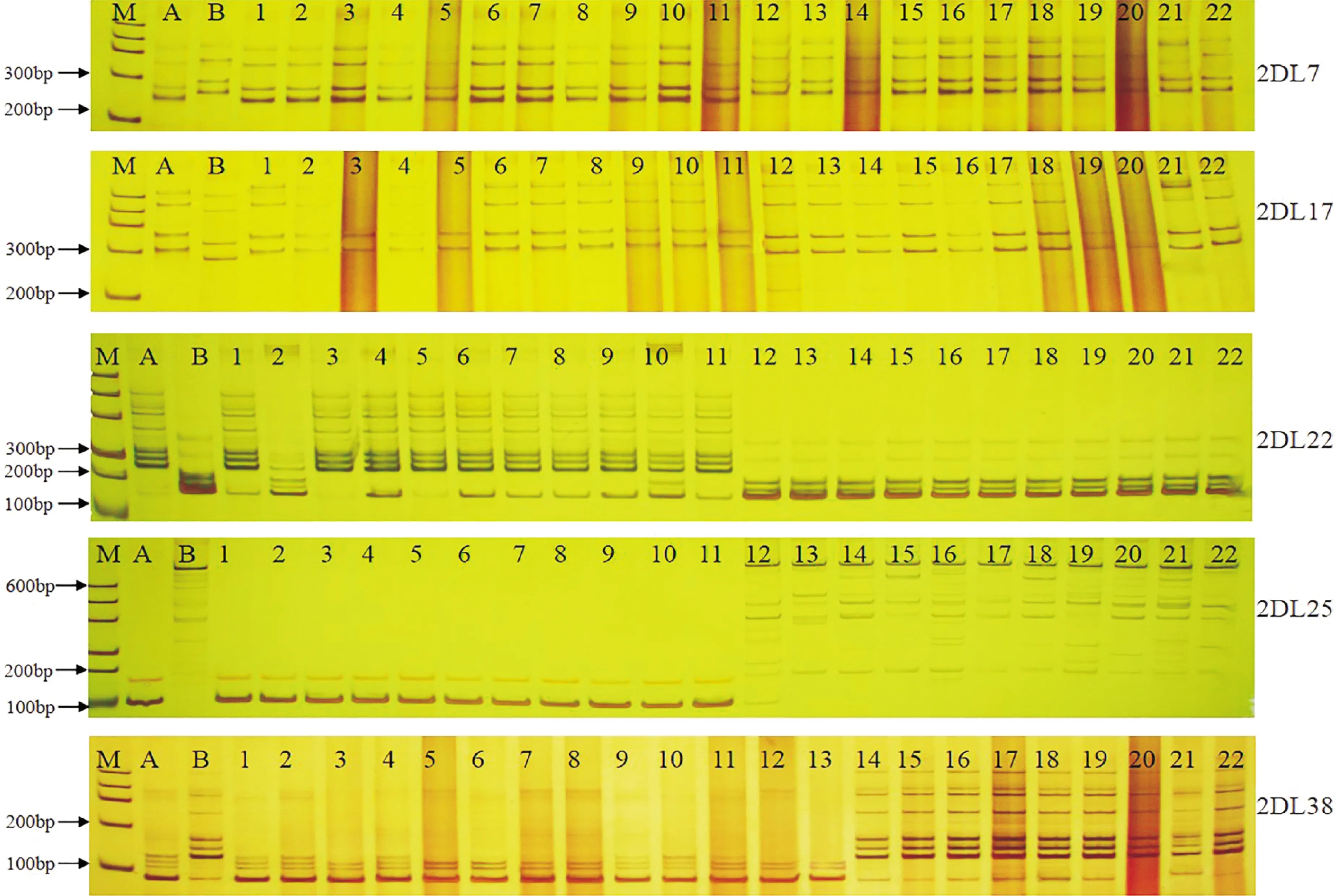
M. Marker;A.三雌蕊亲本(CH257);B.单雌蕊亲本(中国春);1~11.三雌蕊单株;12~22.单雌蕊单株图2 Pis-CH257连锁SSR标记在“CH257×中国春”F2代三雌蕊与单雌蕊小群体扩增结果M. DNA size marker; A. Three pistil parent (CH257); B. Single pistil parent (Chinese Spring); 1-11. Three pistil per plant; 12-22. Single pistil per plantFig.2 Amplification results of Pis-CH257-linked SSR markers in three pistil and single pistil small population of “CH257×CS” F2
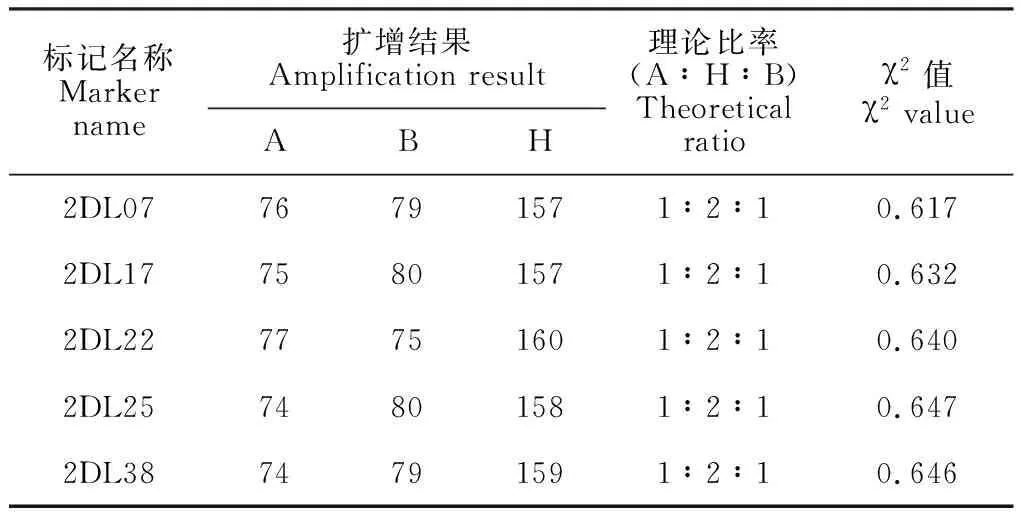
表3 Pis-CH257连锁SSR对F2群体的扩增结果
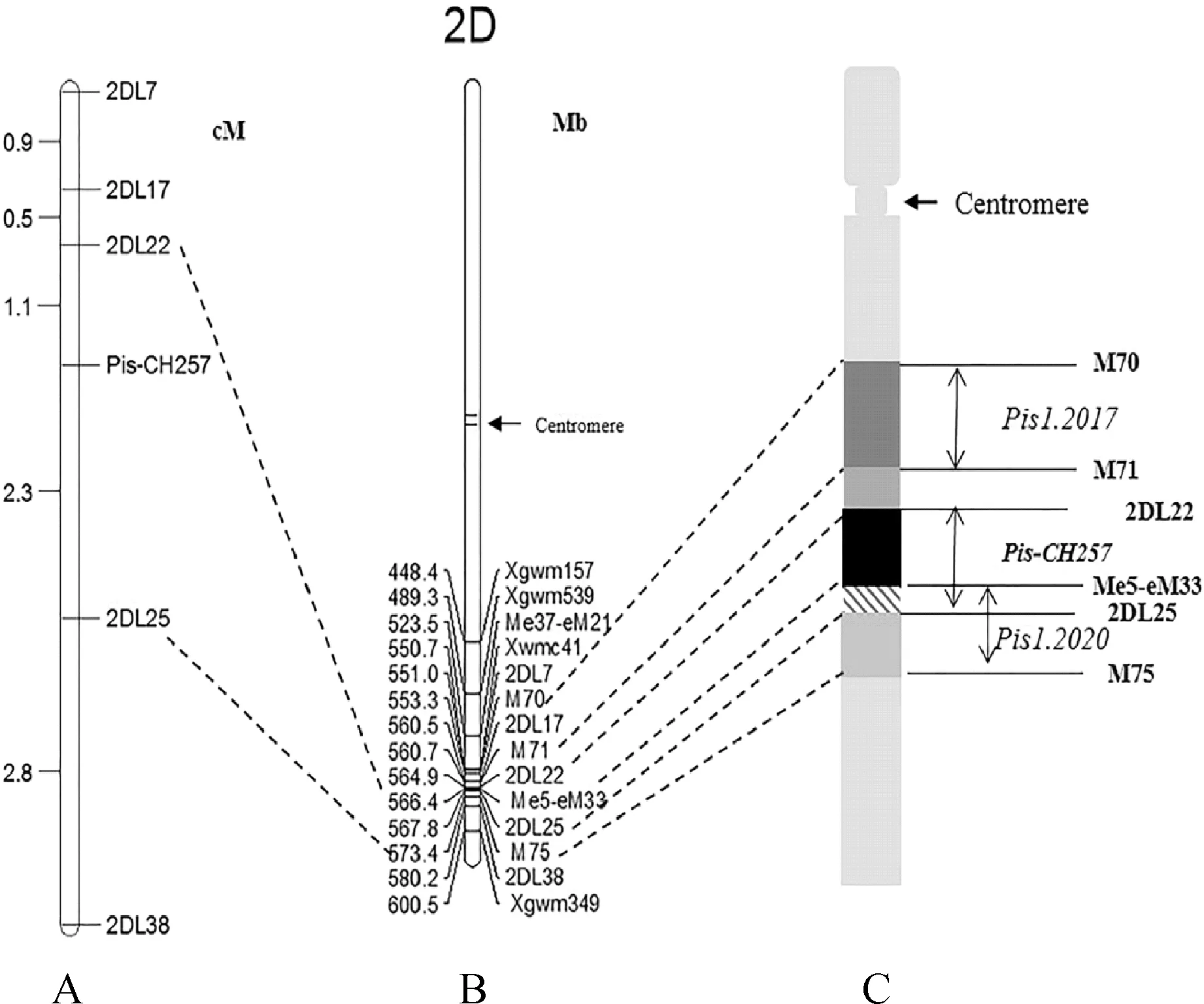
图3 Pis-CH257遗传连锁图谱(A)、Pis-CH257已有标记物理图谱(B)和已有标记比较物理图谱(C)Fig.3 Pis-CH257 genetic linkage map (A), Pis-CH257 existing marker physical map (B) and existing marker physical map (C)
3 讨 论
与其他谷类作物一样,小麦花器官的发育直接影响谷物产量,然而这种花器官异常可以为改良小麦产量提供独特的遗传资源[16]。解析小麦花发育机制可以促进杂交小麦的发展,增加小麦的产量构成[17-19]。对控制小麦花发育基因的分子特征和调控因子的鉴定,可以改变小麦育种策略和加快培育高产小麦品种进程[20-21]。一个小花中发育出3个雌蕊在小麦遗传育种研究中具有极大的利用价值,不仅能够为研究植物花器官发育的机理和遗传机制提供实验材料,而且对小麦杂种优势利用和产量的提高也具有一定的作用[21]。关于三雌蕊性状的遗传机制,马守才等[22-23]的研究认为控制三雌蕊小麦的基因有隐性和显性两种,且受细胞质影响严重;武军等[24]、Peng等[6-7]研究认为,三雌蕊基因受一对显性单基因控制;但Zhu等[25]的研究认为三雌蕊性状受单个半显性基因控制。本研究通过对“CH257/中国春”组合后代群体进行遗传分析后,发现CH257的三雌蕊性状可以稳定遗传给后代,而且在CH257中控制三雌蕊性状的基因是一个显性单基因。
SNP芯片结合传统的分离群体分组分析法(BSA),可以快速检测2个不同表现型构建的混合池间的遗传差异,发现与目标性状关联度高的SNP变异,确定其所在染色体区段,并在此基础上开发与目标基因位点连锁的特异性PCR标记,实现对目标性状控制基因的快速定位[26-27]。本研究采用90K SNP芯片结合BSA法,将CH257中控制三雌蕊性状的基因Pis-CH257初步定位在2D染色体长臂上,之后利用在该区段开发的SSR标记将其定位于两个SSR标记2DL22和2DL25之间。Yang等[28]于2017年将控制小麦三雌蕊性状基因Pis1定位在标记M70和M71之间,两个标记与Pis1的遗传距离分别为1.1 cM 和3.0 cM。Zhu等[25]于2019年开发了4个SSR标记将控制小麦三雌蕊性状的基因12TP定位在2DL染色体上,两侧标记遗传距离7.58 cM。Yu等[29]于2020年利用BSA构建了三雌蕊位点(Pis1)的部分遗传连锁图谱。这些研究都将三雌蕊基因定位在小麦2D染色体上,所有这些研究都表明在2DL染色体臂上有一个控制三雌蕊表型的基因座。将已报道三雌蕊基因连锁标记的物理图谱与本实验中所用标记的物理图谱进行了比较(图3,C)发现, Yang等[28]通过M70和M71标记定位的Pis1物理位置与Pis-CH257所在区段物理位置相邻;Yu等[29]将Pis1定位在分子标记Me5-eM33和M75之间,其物理位置与Pis-CH257区段物理位置接近,且有部分位置重合。出现这种现象的原因可能是由于遗传背景不同所导致,但二者是否是等位基因还需要进一步的实验验证。本研究缩短了三雌蕊基因所在遗传区间,但距离成功克隆三雌蕊小麦还有着不小的差距,下一步将利用来自F2的大群体筛选交换株,进行精细作图。
综上所述,本研究通过对“CH257/中国春”组合的F1、B1C1和F2群体进行遗传分析后得出,CH257中存在一个受显性基因控制的基因位点控制着小麦三雌蕊性状,通过BSA法混池后经90K SNP芯片扫描确定控制三雌蕊的基因Pis-CH257处于2D染色体长臂上。根据基因芯片扫描结果在2D染色体上开发了5个SSR标记将Pis-CH257定位于2DL染色体2DL22-2DL25之间,遗传距离分别为 1.1 cM和2.3 cM,为克隆控制小麦三雌蕊性状的基因提供了基础。
