春梢叶损失对刺梨生长和果实产量与品质的影响机理分析
2022-01-14樊卫国潘学军官纪元周禹佳
樊卫国,潘学军,陈 红,穆 瑞,官纪元,周禹佳
(1 贵州大学 国家林业和草原局刺梨工程技术研究中心,贵阳 550025;2 贵州省植物园,贵阳 550004)
干旱、冰雹、病虫害、大气污染等自然灾害和叶片的采收利用,都会造成经济植物叶片出现不同程度的损失,其损失量过大对其生长和结实会产生不利的影响。研究叶片的损失量对经济植物生长及果实产量、品质和相关生理特性的影响,可为灾后损失的评估和合理采叶量的确定提供科学依据。叶片损失对植物的影响是多方面的,不仅会改变植物的源库器官的比例,而且会造成树体养分的丢失,从而改变器官的生长、生理特性、代谢规律[1]、固碳能力及生物量的分配[2],引起植物的水力特征发生变化,严重时减缓或停滞根和新梢的生长,甚至导致植株死亡[3-6]。同时,植物在叶片损失后,也会在生长及生理上作出一些补偿反应。其中,在叶片损失后的短期内,油橄榄(Oleaeuropaea)[7]、蓝桉(Eucalyptusglobulus)[8]等植物能够提高留存叶片的光合能力,以此弥补叶片的碳同化能力和同化物质分配的不足,这种光合能力的上调主要与落叶后树体的水分关系及营养状况的改变有关[8-10],但随时间的延长,存留叶会加快衰老[7],最终导致固碳能力降低。
果树或其他经济树种发生不同数量的叶损失后,对其生长和果实的产量、品质产生的影响在不同树种间有明显的差异。金争平等对雄性叶用沙棘(Hippophaerhamnoides)摘取适量叶片后,能够促进当年的新梢生长,但摘叶量大于25%后,会对生长产生严重抑制,造成树势的早衰,因此建议采叶量应控制在25%以内[11]。 在春季初花期摘去油橄榄50%的叶片后,会导致座果率和果实产量大幅度降低[7]。5月份模拟病虫危害对油茶(Camelliaoleifera)树冠造成不同的叶量损失后,失叶率超过 25% 的油茶果实产量、茶籽的出籽率、出仁率、含油率都明显降低,翌年树体的生长也受到明显抑制[12]。而对于葡萄而言,在生长期适当摘除部分叶片,不但不会降低果实的产量,而且对果实的品质还会有明显提高的作用[13-14]。
刺梨(RosaroxburghiiTratt.)是原产中国西南地区的特有经济树种,在贵州的种植规模已达15万hm2以上。刺梨的抗旱力极弱,短期的干旱就能导致大量落叶[15]。另外,刺梨叶片易受食叶性害虫危害而造成大量失叶。刺梨果实和叶片中均含有丰富的营养和保健物质[16-18],在贵州刺梨主产区,除了对刺梨果实进行加工利用外,在春季也采收刺梨嫩叶作为食品加工的原料,传统的利用方式是用刺梨嫩鲜叶与茶青混合杀青后干制成茶,这种茶因富含谷氨酸而鲜爽宜人[17]。近年来已出现很多以刺梨嫩鲜叶为原料采用现代制茶工艺加工的产品,由于刺梨叶茶具有独特的茶香和保健功效,在当地市场上供不应求[18]。然而,目前生产上还没有叶用刺梨品种,利用现有品种进行果叶兼用可能还会持续很长时间,因此,在刺梨的果叶兼用过程中,如何确定合理的采叶量是生产中急待解决的重要问题,失叶量大小对刺梨生长及果实产量、品质和相关生理究竟有何影响也值得深入探究。本试验采用摘叶的方法,研究了刺梨叶片不同损失量对新梢生长和果实产量、品质及其相关生理特性的影响,旨在为刺梨合理采叶量的确定提供科学理论依据。
1 材料和方法
1.1 试验材料
试验于2017~2019年在贵州大学国家林草局刺梨工程技术研究中心试验基地进行,试验树为2015年1月定植的 ‘贵农5号’刺梨品种,种苗采用扦插繁殖,株距2.5 m,行距3 m,树势中庸且长势一致。
1.2 试验设计
试验设置4个失叶率处理,失叶率分别为20%、40%、60%和80%,以不摘叶植株为对照。每个处理3株,重复3次。
1.3 摘叶处理
于2017年5月初对参试刺梨树春梢进行摘叶处理。处理前先选择与试验树长势及大小一致的3个单株,分别摘除全部春梢叶片,称重后计算单株平均鲜叶重,再分别计算出20%、40%、60%、80%失叶率的单株平均鲜叶重,据此进行摘叶操作。在对不同处理的刺梨树春梢进行摘叶时,失叶率20%、40%、60%和80%处理中每5片叶分别摘除1、2、3和4片,使不同处理的失叶量分别达到单株鲜叶重的20%、40%、60%和80%。
1.4 观测指标及方法
1.4.1 新梢生长状况刺梨摘叶后的当年、翌年和第3年,定时观察不同处理各次新梢的萌芽时期及生长状况。于叶损失的当年、翌年和第3年的10月上旬刺梨新梢全部停长后,测定不同处理的春梢、夏梢和秋梢的数量,同时在不同处理的每个植株上随机选择春梢、夏梢和秋梢各50个,测定其长度及梢径。
1.4.2 座果率和果实产量在失叶当年、翌年和第3年的开花期,测定不同失叶率处理及对照植株的花朵数,果实成熟期测定单株的果实数、平均单果重和单株果实平均产量,计算各年的座果率。
1.4.3 果实品质指标在叶损失当年、翌年和第3年的果实成熟期,随机取不同处理及对照的果实各3份,每份20个果,带回实验室进行可溶性固形物、可溶性总糖、可滴定酸、维生素C及总黄酮的含量测定。测定总黄酮含量的果实在取样后立即杀酶、烘干、粉碎过40目筛备用。果实可溶性固形物含量采用数字折光仪测定,可溶性总糖含量采用蒽酮法测定,可滴定酸含量采用酸碱滴定法测定(GB 12293-1990,以苹果酸作为其换算系数),维生素C含量测定采用2,6-二氯靛酚法(GB/T 6195-86),总黄酮含量测定参照杜薇等[19-20]的方法。
1.4.4 叶片营养元素含量在失叶处理当年、翌年和第3年的6月中旬(刺梨营养诊断期),对不同处理及对照当年春梢中部存留叶和翌年、第3年春梢中部叶片进行取样后,按李港丽提供的果树叶样预处理方法[21]进行洗涤、烘干、粉碎后过60目尼龙筛,测定叶中的营养元素含量。其中,N含量测定用凯氏定氮法[21],P含量测定用钒钼黄比色法[21],K含量测定用火焰光度计法[21],Ca、Mg、Fe、Mn、Zn、Cu含量测定用电感耦合等离子发射光谱法[22],B含量测定用姜黄素比色法[21]。
1.4.5 失叶当年春梢中内源激素含量从失叶处理后的1~30 d内,每隔5 d对不同处理及对照植株春梢进行1次取样。取样部位为失叶春梢中部的带芽体梢段,不同处理及对照各取样3份,取样后立即将样品置于冰壶内带入实验室,称取0.5 g,用80%冷甲醇提取内源激素,采用酶联免疫法[23]测定其中的生长素(IAA)、赤霉素(GA1+3)、玉米素核苷(ZR)和脱落酸(ABA)含量。测定内源激素的试剂盒由中国农业大学提供。
1.4.6 失叶当年春梢存留叶片的光合指标从失叶处理后的第1天起至第30天内,每隔5 d用Li-6400 便携式光合测定系统对不同处理失叶春梢存留叶的净光合速率(Pn)和蒸腾速率(Tr)进行1次测定,每次测定的时间均在上午10:00~11:00。然后按Pn/Tr计算水分利用效率(WUE)。
1.4.7 失叶当年果实中维生素C合成积累关键酶活性在失叶当年8月初刺梨果实快速膨大期(维生素C合成积累的关键期)[24],取不同处理及对照的果实用液氮冷冻处理后置于超低温冰箱(-79 ℃)中保存,用于测定对维生素C合成、积累有极其重要影响的L-半乳糖-1,4-内酯脱氢酶(GalLDH)、单脱氢抗坏血酸还原酶(MDHAR)、脱氢抗坏血酸还原酶(DHAR)、抗坏血酸氧化酶(AAO)、抗坏血酸过氧化物酶(APX)的活性。其中,GalLDH、MDHAR、APX、AAO活性采用分光光度法测定,参照试剂盒实验说明书进行酶的提取;DHAR活性采用酶联免疫法测定,酶的提取及具体测定方法按试剂盒实验说明书进行。上述酶活性的测定试剂盒由北京索莱宝科技有限公司提供。
1.4.8 失叶当年果实中类黄酮合成积累关键酶活性在失叶当年8月中旬果实成熟期(类黄酮合成积累的关键时期),取不同处理及对照的果实用液氮冷冻处理后,置于超低温冰箱(-79 ℃)中保存后,用酶联免疫法测定果实中影响类黄酮物质合成的苯丙氨酸解氨酶(PAL)、肉桂酸-4-羟化酶(C4H)、4-香豆素辅酶A连接酶(4CL)和查尔酮合成酶(CHS)的活性,酶的提取分别参照Brödenfeldt R.等[25]和范存斐等[26]的方法。上述酶活性的测定试剂盒和总黄酮含量的标准品均由深圳子科生物科技有限公司提供。
1.5 数据整理与统计
采用Microsoft Excel 2007进行数据整理并作图, 用 DPS v7.05 软件进行差异显著性和相关性分析, 多重比较采用邓肯氏新复极差法。
2 结果与分析
2.1 春梢失叶对刺梨不同年份新梢生长的影响
2.1.1 当年新梢生长试验结果(表1)表明:失叶刺梨当年春梢的数量在不同失叶率处理间无显著差异,但失叶会促进刺梨当年的夏梢萌发,增加夏梢数量,并使秋梢数量及长度、粗度明显减小。其中,随着刺梨春梢失叶量的增大,当年夏梢的数量随之增加,而秋梢数量却随之减少;失叶率20%~80%处理的夏梢数量比对照增加了 1.55~3.43倍,秋梢的数量比对照减少了26.19~99.78个,且当年夏梢及秋梢数量在不同处理和对照间差异均达到显著水平(P<0.05)。失叶率20%和40%处理的当年夏梢长度和梢径与对照的差异不显著,而其当年秋梢的长度和梢径随失叶率的增大而显著减小。另外,试验中还观察到,失叶使当年夏芽萌发和抽生的时间提早,失叶率超过20%的3个处理,夏芽提早膨大,夏梢抽生的时间比失叶率20%的处理和对照提早了10 d以上。
2.1.2 翌年和第3年新梢生长刺梨春梢失叶量过大会严重抑制翌年和第3年新梢的生长(表1)。其中,刺梨失叶后第2年,失叶率20%处理的春梢、夏梢及秋梢的数量、梢长、梢径与对照无显著差异;但失叶率超过20%的3个处理,3次新梢的数量、长度及梢径都显著小于对照。至刺梨失叶后的第3年,失叶率60%和80%处理的3次新梢的数量、长度及梢径仍然显著低于对照和失叶率20%及 40%处理。由此可见,刺梨春梢失叶率超过20%后,对其新梢生长的抑制作用能够持续多年;失叶率40%以上处理的树势衰弱,到失叶后翌年和第3年的春梢抽生仍然明显比对照晚。
2.2 春梢失叶对刺梨当年春梢中内源激素含量的影响
图1显示,从失叶后5 d开始,春梢中IAA、 GA1+3、ZR和ABA的含量就已在不同处理间表现出明显差异,且随着失叶时间的增加处理与对照间的差异逐渐明显增大。其中,各失叶处理春梢中的IAA和ABA含量与对照相比均表现出随失叶率增大而降低的趋势;在失叶后5 d时,除失叶率20%处理的IAA含量与对照差异不显著外,其他3个处理与对照的差异均达到显著水平(P<0.05),从失叶5~30 d,所有处理春梢中的IAA含量均随失叶率增大而显著降低;各处理失叶春梢中ABA的含量变化与IAA类似,在失叶后5 d内,春梢中的ABA含量就迅速下降,在此后的时间内均表现出随失叶率增大而显著降低的趋势。同时,各处理春梢中GA1+3和ZR的含量随失叶时间的变化趋势正好与IAA和ABA含量的相反,表现出不断增加的趋势;从失叶后10 d开始, GA1+3和ZR含量增加在处理之间和处理与对照间已经有很大差异。因此,失叶春梢中内源激素含量变化可能对当年的夏梢萌发、存留叶的光合及水分生理产生重要的调控作用。
2.3 春梢失叶对刺梨当年春梢存留叶片光合特性的影响
图2显示,从4月下旬至5月下旬(失叶后30 d内),对照组(CK)春梢叶片的净光合速率(Pn)呈不断增强的趋势,这是叶片逐渐发育成熟的正常生理表现,而各失叶处理春梢存留叶的Pn则表现出先升高后降低的趋势。其中,在失叶后的15 d内,各失叶处理春梢存留叶的Pn都迅速提高,而且表现出随失叶率的增大而明显增强的趋势;到失叶20 d后,各失叶处理的Pn增强的趋势开始减弱,且失叶率80%处理的Pn比前期明显下降;失叶25 d之后,所有失叶处理的Pn都低于对照,而且随失叶率的增大Pn下降的趋势也十分明显。春梢存留叶的Pn在失叶后短期内迅速增强,可能是光合补偿的生理表征,而其Pn在失叶后期的迅速降低可能与存留叶中养分的转移导致的叶片生理衰老有关。
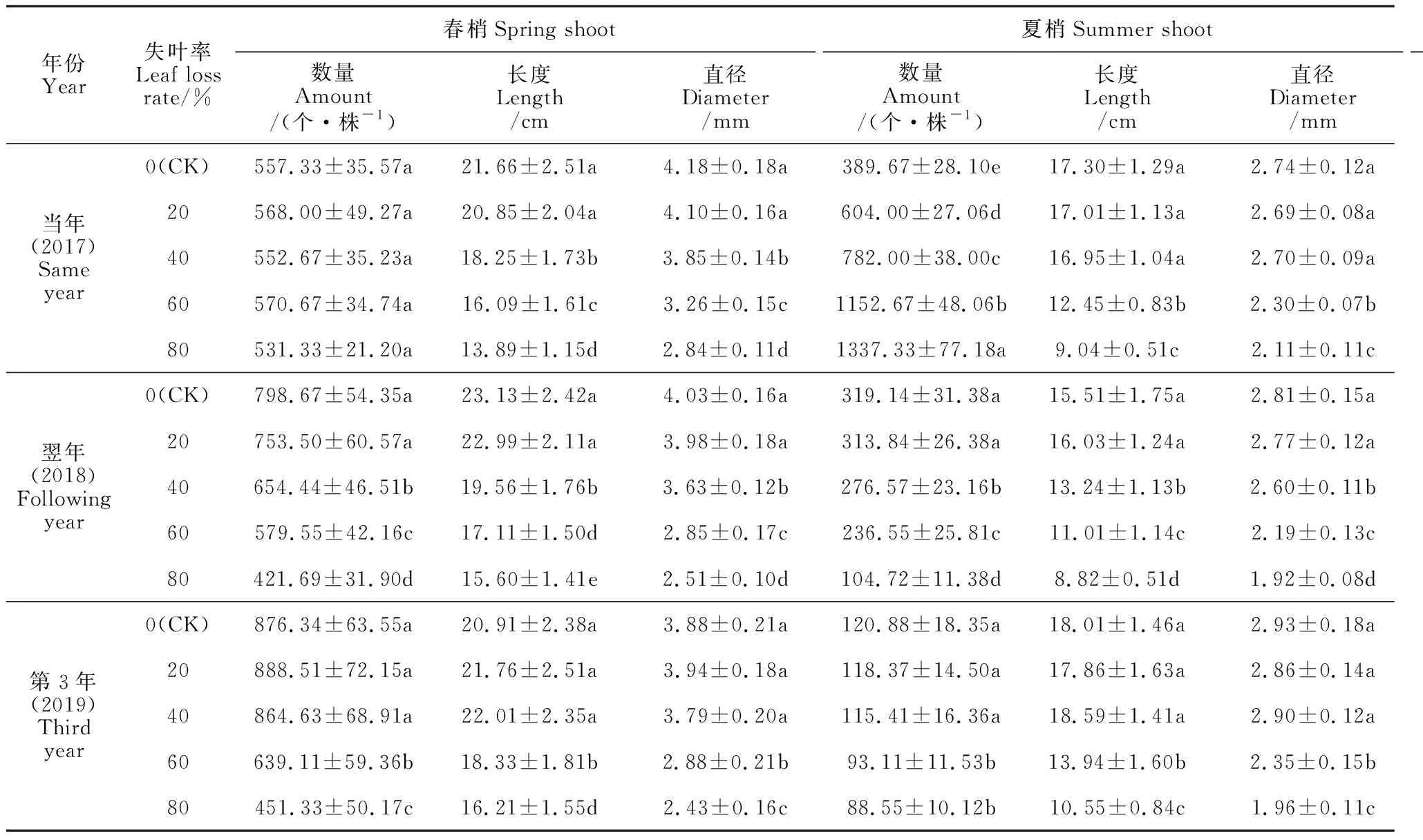
表1 春梢叶片损失对刺梨新梢生长的影响

图1 不同春梢失叶量下刺梨当年春梢内源激素含量的变化 Fig.1 The endogenous hormone contents change of R. roxburghii spring shoots in same year under different leaf loss rates
同时,图2还显示,所有失叶处理春梢存留叶的Tr都显著高于同期对照,且其Tr均随失叶率的增大而增加,不同处理间的差异也均达到显著水平(P<0.05);随着失叶天数的增加,各处理春梢存留叶的Tr均逐渐增加。这表明失叶会加快刺梨春梢存留叶的水分蒸腾,且失叶率越大、失叶时间越长这种增加的趋势越明显,这对维持失叶植株的水分平衡和生理代谢具有重要作用。
另外,水分利用效率(Pn与Tr的比值)是光合作用过程中对水分有效利用的重要生理指标。试验结果(图2)表明,各失叶处理刺梨春梢存留叶WUE均不同程度地低于同期对照,并随着失叶率的增加而逐渐降低,且在相同时期内处理间及其与对照间差异大多达到显著水平;随着失叶时间的延长,各失叶处理春梢存留叶WUE均表现出先升后降的变化趋势,且均在失叶后15 d左右达到最大值。说明失叶处理明显降低了刺梨春梢存留叶WUE,且失叶率越大,WUE降幅越大,这与失叶后春梢存留叶的水分蒸腾加剧有关。
2.4 春梢失叶对刺梨不同年份叶片营养元素含量的影响
2.4.1 常量营养元素含量表2显示,春梢失叶过多会明显降低各年份刺梨叶中的常量营养元素N、P、K、Mg的含量,且降幅随着失叶率的增加而增大。其中,在失叶率20%及以上的处理中,当年存留的春梢叶中N、P、K、Mg含量都显著低于相应对照,而Ca的含量在不同处理间的差异不显著,这可能与N、P、K、Mg元素容易转移和Ca元素不容易转移的特性有关;除Ca以外,存留春梢叶中N、P、K、Mg含量随失叶率的增大而显著减少,这种不利影响可持续至翌年甚至第3年。在失叶翌年,失叶率20%处理的春梢叶中N、P、K、Mg含量与对照已经没有显著差异,但失叶率超过20%的3个处理的N、P、K、Mg含量仍然明显低于对照,说明失叶率过大处理的这些元素含量到翌年都难以恢复到正常水平。在失叶后的第3年,失叶率20%~40%处理的春梢叶中N、P、K、Mg含量均与对照没有显著差异,而失叶率60%和80%两个处理的N、P、K、Mg含量
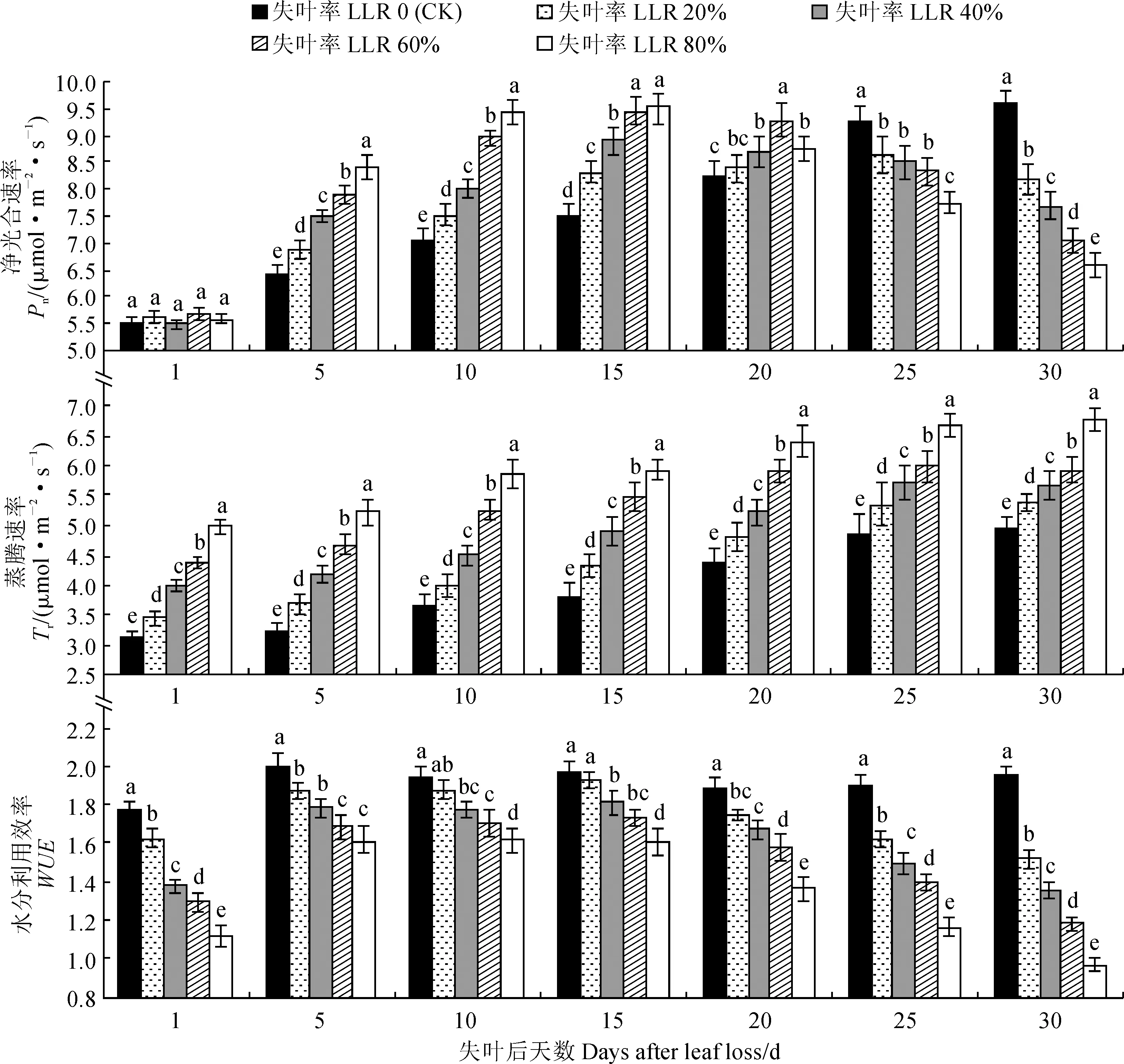
图2 不同失叶量下刺梨当年春梢存留叶Pn、Tr和WUE的变化Fig.2 The Pn, Tr and WUE change of spring shoots remaining leaves of R. roxburghii in same year under different leaf loss rates
仍然显著低于对照。另外,虽然失叶当年存留春梢叶中Ca的含量在不同处理间没有显著差异,但翌年失叶率60%和80%处理,以及第3年失叶率80%处理春梢叶中Ca含量仍然显著低于对照和失叶率20%、40%处理(P<0.05),说明刺梨春梢失叶量过大对其Ca含量的不利影响要到翌年后才表现出来。
2.4.2 微量营养元素含量从表3可知,在刺梨失叶当年春梢存留叶中,不同失叶率处理及对照间的Fe、Mn、Cu、B微量元素含量差异均不显著,但失叶率60%和80%两个处理的Zn含量显著比同期其他处理和对照显著降低,说明当年刺梨春梢存留叶中Fe、Mn、Cu、B元素含量在不同失叶率处理下均未显著影响,而其Zn含量则在失叶量过大时显著降低。在翌年春梢叶中,失叶率60%和80%两个处理的Fe、Cu含量显著低于对照和失叶率20%及40%的处理;失叶率80%的处理的Mn含量显著低于其他处理;失叶率大于20%处理的B含量显著低于失叶率20%处理和对照,说明失叶对春梢叶中Fe、Mn、Cu、B元素含量的降低作用在翌年才表现出来。到失叶后的第3年,刺梨春梢叶中Fe、Mn、Cu的含量在各处理与对照间已无显著差异,但失叶率60%、80%处理的Zn、B含量仍然明显比对照和失叶率20%、40%处理显著降低,说明失叶量过大对刺梨树体春梢叶片Zn、B损失的不利影响同样会持续多年。
2.5 春梢失叶对当年刺梨果实中维生素C和类黄酮物质合成、积累关键酶活性的影响
在植物维生素C合成的L-半乳糖途径中,L-半乳糖-1,4-内酯脱氢酶(GalLDH)直接催化半乳糖内酯形成维生素C[27-28]。维生素C可被抗坏血酸氧化酶(AAO)和抗坏血酸过氧化物酶(APX)催化氧化分解为单脱氢抗坏血酸(MDHA),再通过非酶歧化反应形成脱氢抗坏血酸(DHA),DHA在MDHAR和DHAR的作用下被还原为AsA[29-31],因此,AAO 和APX能够促进AsA的氧化分解,而MDHAR和DHAR则促进AsA的循环再生。表4结果表明:在刺梨果实维生素C大量合成积累的关键时期,不同失叶处理果实中GalLDH活性均远比AAO、APX和MDHAR、DHAR活性要强得多,说明失叶后GalLDH在刺梨果实维生素C的积累过程中起主导作用,这与刺梨具有维生素C不易被氧化从而大量积累的生理特性[32]有关;同时,随失叶率增大,刺梨果实中GalLDH活性逐渐明显减弱,且除失叶率20%以外的其他处理的GalLDH活性都比对照显著降低,而同期的AAO、APX、MDHAR、DHAR的活性则均呈现出增强的趋势,说明春梢叶损失后刺梨果实中出现了维生素C氧化分解和循环再生的过程。
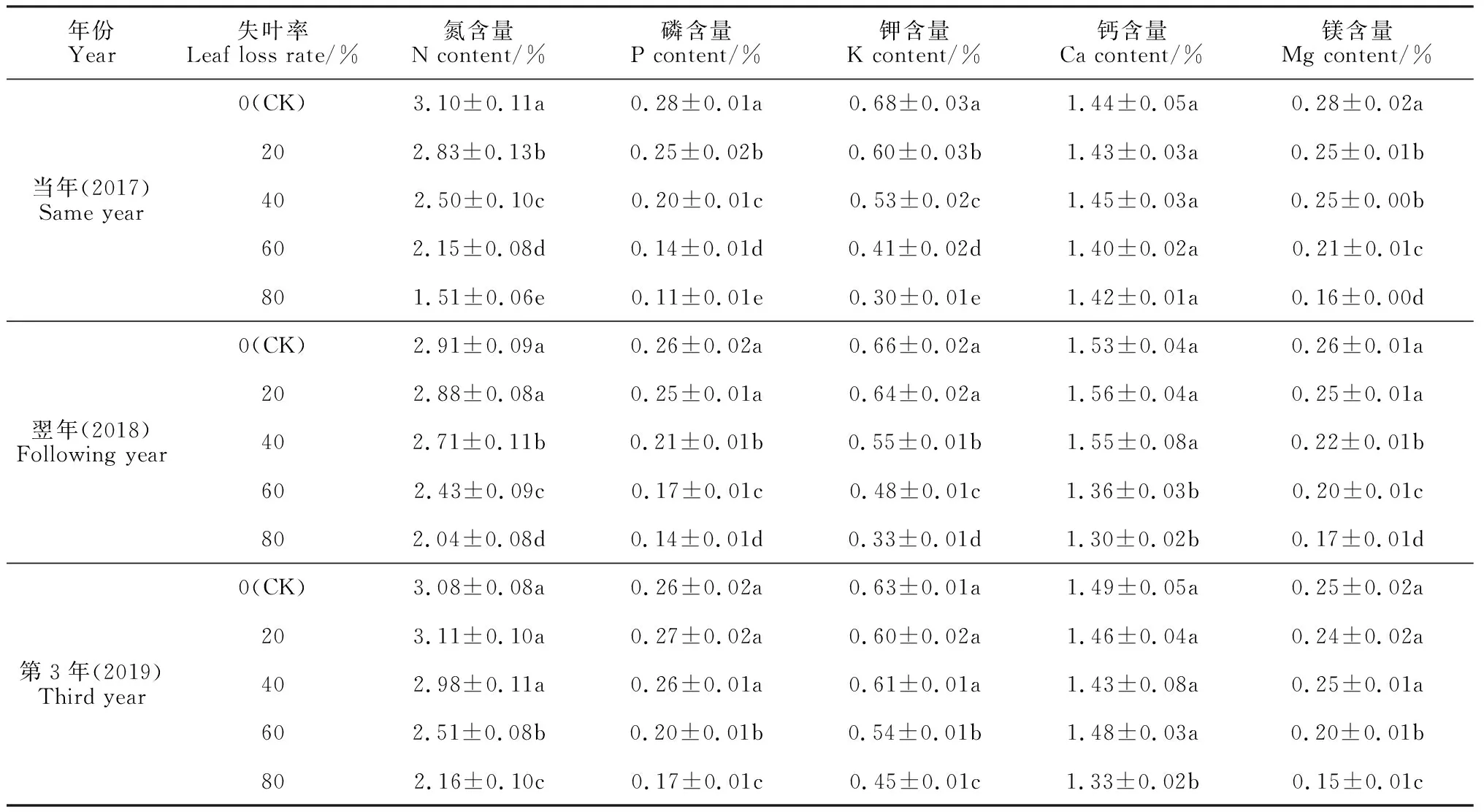
表2 失叶对不同年份刺梨春梢叶片中常量营养元素含量的影响
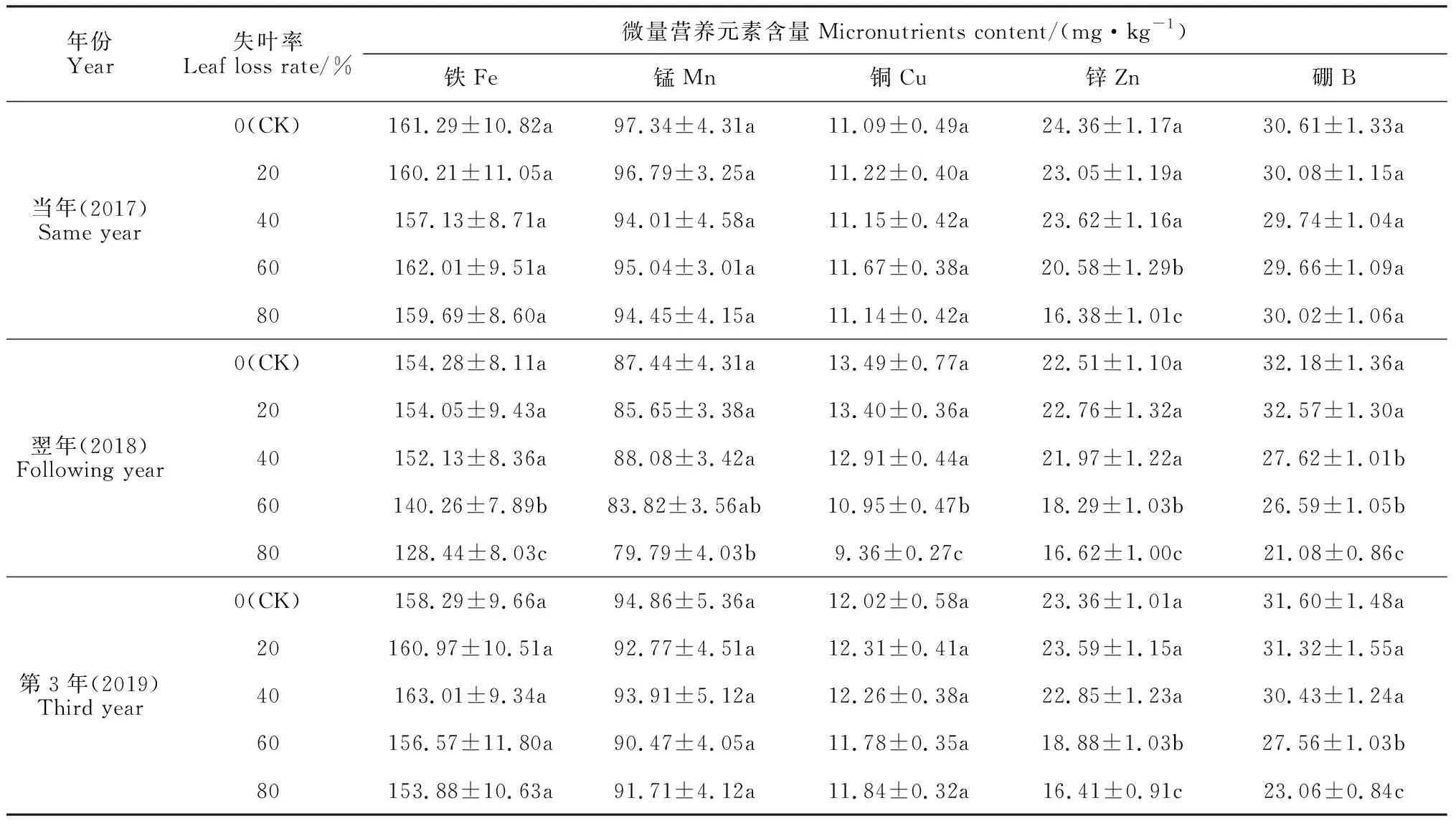
表3 失叶对不同年份刺梨春梢叶片中微量营养元素含量的影响
苯丙烷途径是植物体内类黄酮物质合成代谢的重要途径,其中,苯丙氨酸解氨酶(PAL)、肉桂酸-4-羟化酶(C4H)、4-香豆素辅酶A连接酶(4CL)和查尔酮合成酶(CHS)是类黄酮物质合成代谢的关键酶[33-35]。从表5可以看出,在春梢失叶当年,刺梨果实中PAL、C4H、4CL和CHS的活性随失叶量的增大而逐渐减弱,总黄酮的含量也随之降低;在不同失叶处理中,除失叶率20%处理的4CL活性与对照差异不显著外,上述4种类黄酮物质合成代谢关键酶的活性在处理间及其与对照间的差异均达到显著水平(P<0.05);在相同失叶处理下,CHS的活性明显最高,其次是4CL活性,而PAL、C4H活性远低于前两者。以上结果说明春梢叶损失对刺梨果实中类黄酮物质的合成代谢有抑制作用,其相关机理有待进一步研究。
2.6 春梢失叶对不同年份刺梨果实产量及品质的影响
2.6.1 果实产量春梢失叶量过大会严重降低刺梨的座果率、单果重和果实产量(表6)。其中,在春梢失叶后3年内,失叶率20%处理的刺梨座果率、单果重和单株果实产量与对照的差异均不显著;但随失叶量的增大(失叶率40%~80%),刺梨当年的座果率、平均单果重和单株果实产量逐渐显著降低,并各处理间及其与对照间均差异显著;在失叶翌年,失叶率超过20%的 3个处理的刺梨果实产量仍然显著小于对照和失叶率20%的处理;直至失叶第3年,失叶率40%处理的产量才恢复正常,而此时失叶率60%和80%两个处理的产量指标都仍然显著低于其余处理和对照。这说明春梢失叶量超过20%后,失叶对刺梨结果和产量造成的不利影响能够持续2年以上甚至更长;失叶量过大导致的树体养分丢失、树势减弱、座果率降低、果实变小是刺梨果实产量降低的根本原因。
2.6.2 果实品质刺梨春梢失叶会不同程度地降低当年、翌年和第3年果实的可溶性固形物、可溶性总糖糖、可滴定酸、AsA和总黄酮的含量;各指标的降幅随着失叶时间增加而减小,随着失叶量的增加而明显增加,且失叶量过大会严重降低刺梨果实以上品质指标(表7)。
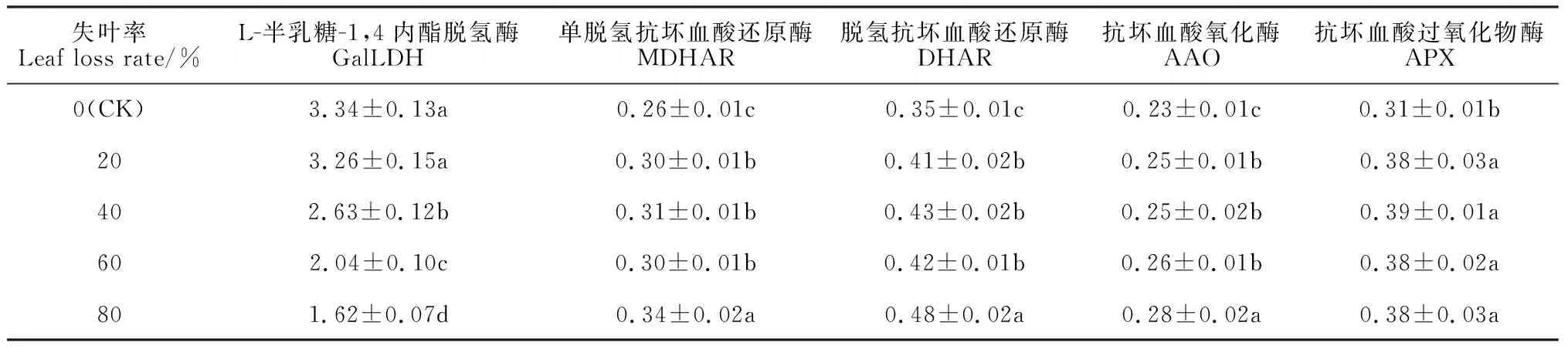
表4 叶片损失对当年刺梨果实中维生素C积累的相关酶活性的影响

表5 叶片损失对当年刺梨果实中类黄酮物质合成关键酶活性的影响
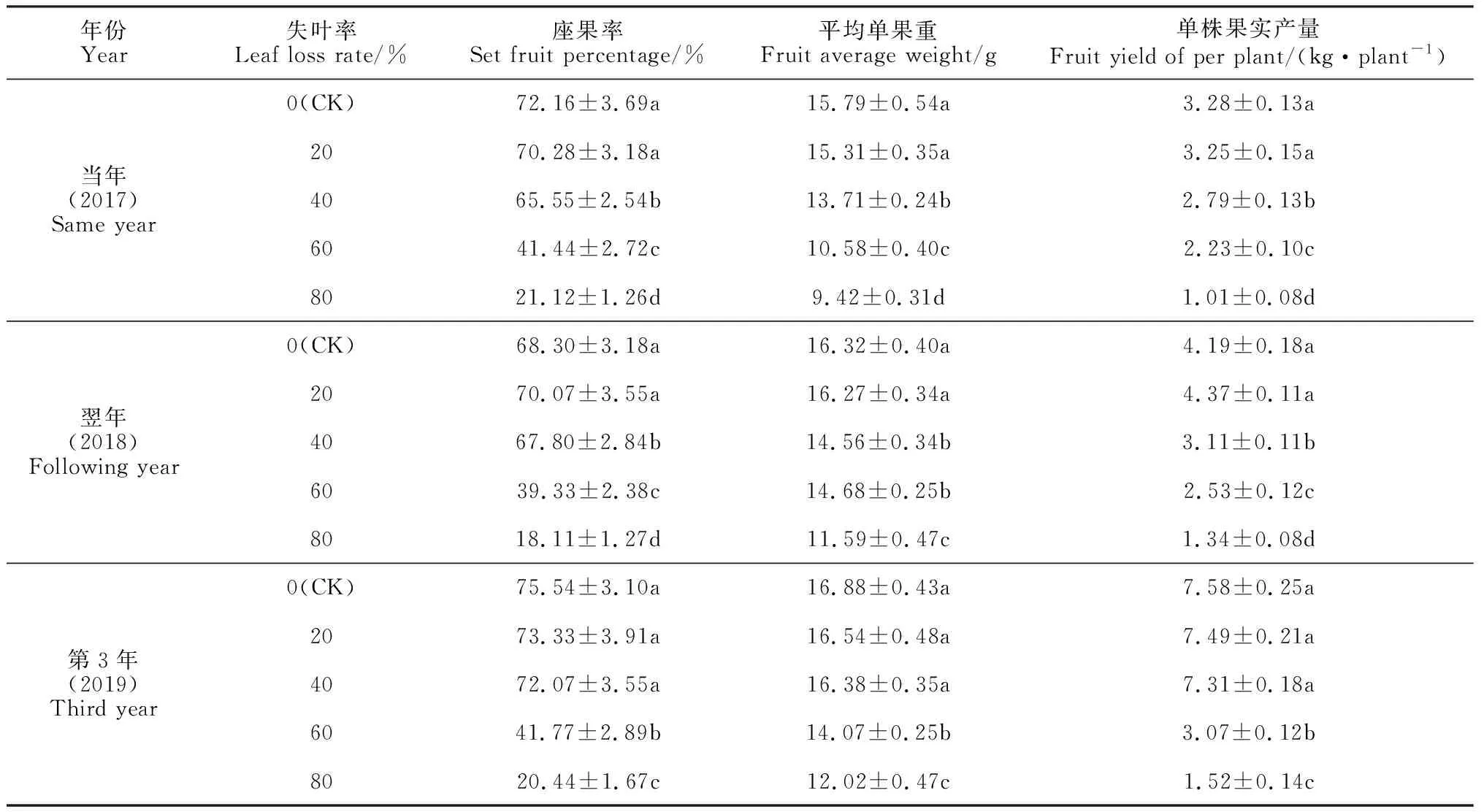
表6 春梢叶片损失对不同年份刺梨果实产量的影响
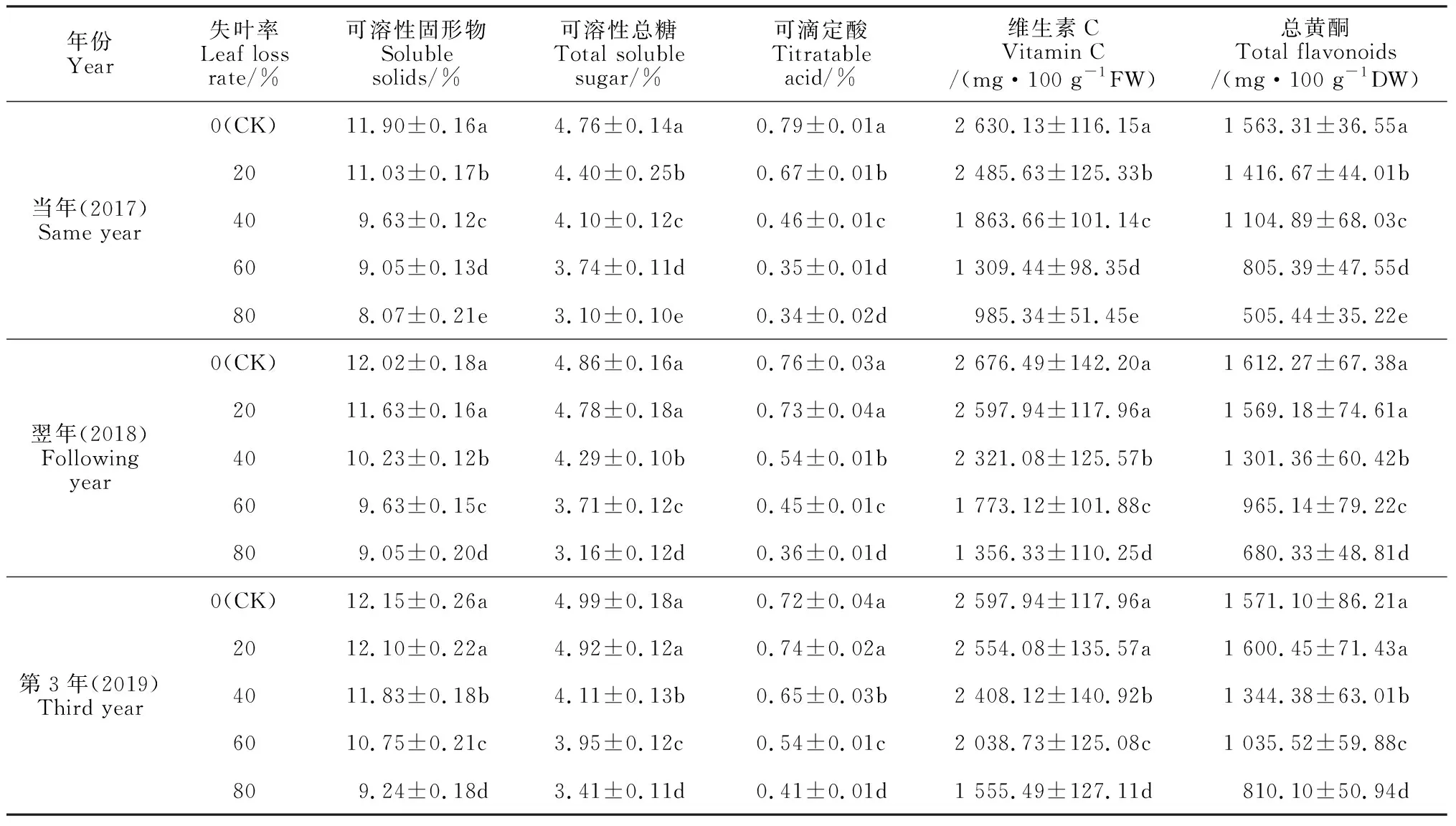
表7 春梢叶片损失对不同年份刺梨果实品质的影响
首先,随春梢失叶量的增大,刺梨当年的果实中可溶性固形物、可溶性总糖、可滴定酸、AsA和总黄酮的含量均表现出十分明显的逐渐降低趋势,且所有处理与对照间的差异均达到显著水平(P<0.05)。其中,失叶率40%~80% 处理果实中AsA含量分别比对照显著减少了29.14%~62.54%,它们的总黄酮含量分别比对照显著减少了29.32%~67.67%。可见,失叶量过大对刺梨果实品质的不利影响极其严重,这种影响可能与源库器官比例改变后引起的营养物质合成、运转、分配的相关生理变化有密切的关系。
其次,在春梢失叶后的翌年和第3年,虽然失叶率20%处理刺梨果实所有品质指标与对照间的差异已经不显著,但失叶率超过20%的3个处理的上述品质指标仍然显著低于对照,说明失叶量大于20%后,失叶对刺梨果实品质的不利影响持续时间较长。另外,在试验中还观察到,春梢失叶能够使刺梨果实着色有所提早,果实上的皮刺也有所变硬。
3 讨 论
3.1 果叶兼用刺梨的合理采叶量、间隔期和采叶后的养分管理
刺梨主产区主要是在春季采收刺梨嫩叶进行加工利用,在尚无叶用刺梨品种的情况下,对果叶兼用刺梨的采叶量应严格控制在总叶量的20%以内,否则会造成严重的树体养分损失,导致树势衰弱和严重减产,降低果实的品质。对于结果较多和树势较弱的刺梨,最好不要采叶。在采叶过程中,对营养生长旺盛的树和树冠上必须剪掉的徒长枝及基部萌蘖枝上的叶片可适当多采。由于失叶对刺梨树体中N、P、K、Mg等养分的损失影响可持续多年,即便在采叶量不超过总叶量20%的前提下,也建议对果叶兼用刺梨的采叶间隔期最好在1年以上。采叶后要及时加强刺梨的养分管理,及时增施速效N、P、K肥和Mg肥,适当补充Zn、B等微量元素,以便尽快恢复树势,避免因失叶对果实产量和品质造成的不利影响。
3.2 失叶对刺梨生长、果实产量及品质和相关生理的影响
有些植物在失叶后不仅不会限制其生长,而且还能促进生长,甚至加快生长,这种现象称之为植物的补偿反应或超补偿反应[12, 36-37],其生理机制可归纳为光合作用、同化产物及营养元素的代谢、运转和内源激素变化对生长的调控,其中,细胞分裂素在该机制中发挥了重要作用[36]。IAA、ABA浓度的降低能够解除对新梢侧芽萌发的抑制,而GA和CTK浓度的增加有利于促进新梢侧芽萌发的生长,此外,ABA浓度的降低和CTK浓度的增加对促进叶片气孔开启和增强光合作用有积极的作用[23]。在本试验中,随失叶率的增大,刺梨失叶春梢中的IAA、ABA含量降低,GA1+3和ZR含量增加,当年的夏芽萌发期提早,夏梢的数量增大,在短期内春梢存留叶的Pn及Tr增强,这是刺梨在失叶后短期内的应急生长和生理补偿反应,刺梨失叶后的生长及光合反应与失叶春梢中内源激素变化密切关联。然而,由于失叶造成刺梨树体养分的大量损失,使刺梨对失叶的生长及生理补偿反应难以长期维持,类似的结果在油橄榄[7]、沙棘[11]、杨树[38]、棉花[36]等植物上都有报道。
失叶对果树及经济树种果实产量及品质的影响不尽相同。在葡萄上,由于摘除部分叶片改善了光照条件,增强了叶片的光合产物合成和糖的输出,使葡萄果实着色度、糖度及花青素、氨基酸、总酚的含量都有所提高[13,39-41],进而改善了果实的品质,苹果[42]上也有类似的报道。然而在油茶上,失叶量在12. 5%~25%的范围内对茶籽产量、品质和翌年树势的生长影响不大;当叶损失量超过 25%,油茶的长势、茶籽产量、出油率和油脂品质严重降低,而且这种不利影响可持续多年[12]。在油橄榄上,初花期结果枝上的叶片损失50%后,座果率、单果重和果实产量明显降低[7]。维生素C和黄酮类物质含量极其丰富是刺梨果实最重要的品质特征,刺梨春梢叶损失量超过20%后,不仅会严重降低当年、翌年和第3年的果实产量,而且会明显降低果实的维生素C和类黄酮物质的含量,不利于保持刺梨特有的品质。糖类物质是维生素C和类黄酮物质合成的重要底物,刺梨果实中维生素C的积累量与GalLDH的活性、可溶性糖的含量呈极显著正相关[43],PAL、C4H、4CL和CHS都参与了类黄酮物质合成积累的调控[31-34]。刺梨大量失叶后,由于光合源器官的减少,导致固碳能力减弱,输入果实中的糖类物质减少,同时果实中GalLDH、PAL、C4H、4CL、CHS的活性也明显降低,这些都是大量失叶后刺梨果实中维生素C和类黄酮物质减少的重要生理原因。
4 结 论
刺梨春梢失叶后,在短期内失叶春梢中的IAA和ABA含量迅速降低,GA1+3和ZR含量迅速增加,失叶春梢存留叶的Pn会迅速增强,当年夏梢的数量会增加,当年秋梢和翌年及第3年的新梢生长会受到明显抑制。春梢失叶会降低刺梨叶片中的营养元素含量,导致刺梨树体养分损失,对N、P、K、Mg元素的损失影响尤其严重。随失叶率的增大,刺梨果实的产量、品质明显降低。大量失叶后刺梨果实中糖的含量减少和GalLDH、PAL、C4H、4CL、CHS的活性减弱,是导致刺梨果实中维生素C和类黄酮物质含量减少的重要生理原因。失叶量过大对刺梨生长和果实产量、品质的不利影响可持续多年。在春季对果叶兼用的刺梨进行嫩叶采收利用时,采叶量应控制在20%内,采叶的间隔期最好在1年以上。
