水稻黑条矮缩病株不同分蘖内病毒含量分析
2022-01-14王海涛宛柏杰徐秋芳周益军
王海涛,宛柏杰,董 岩,王 满,徐秋芳,3,周益军
(1.江苏省农业科学院植物保护研究所/江苏省食品质量安全重点实验室-国家重点实验室培育基地,江苏 南京 210014;2.江苏沿海地区农业科学研究所,江苏 盐城 224002;3.江苏大学生命科学研究所,江苏 镇江 212013)
水稻黑条矮缩病是由呼肠孤病毒科(Reoviridae)斐济病毒属(Fijivirus)中的成员水稻黑条矮缩病毒(rice black-streaked dwarf virus,RBSDV)侵染引起的一种病害[1-3]。RBSDV除可侵染水稻外,还可以侵染小麦、玉米、大麦、燕麦和高粱等禾本科作物,是制约东亚地区粮食生产的重要植物病毒[1,3]。RBSDV基因组由10条线性双链RNA组成,可编码6个结构蛋白和7个非结构蛋白[4]。其中,由基因S10编码的P10是病毒主要的外层衣壳蛋白[5]。RBSDV主要由介体灰飞虱(Laodelphax striatellusFallén)以持久性、增殖型的方式进行传播,侵染引起水稻植株矮缩、叶片浓绿僵直、不抽穗或穗小,在叶背和茎基部有白色至褐色瘤状突起[1]。
由病原微生物侵染引起的植物体内激素的变化,可以促进或抑制病原物对植物的侵染,使植物表现出更为严重或变轻的病症[6-7]。水稻受病毒侵染后通常表现为植株矮小,但仍可进行分蘖,可能与水稻体内激素水平的改变有关。目前,关于RBSDV侵染水稻后如何调控植株内源激素相关的信号通路,进而影响其侵染的报道已较为详细,如:RBSDV的侵染可以激活水稻茉莉酸(jasmonic acid,JA)信号通路,抑制其油菜素内酯(brassinosteroids,BRs)途径,进而抑制病毒对水稻的侵染[8];RBSDV侵染水稻后引起脱落酸(abscisic acid,ABA)含量的增加,进而通过抑制JA信号途径调节水稻内活性氧水平来负调控水稻对RBSDV的防御[9-10];抑制生长素(auxin)信号,可以促进水稻对RBSDV的敏感性,导致更为严重矮化的症状[11];等等。然而目前仍有一些基础问题尚不明确,如关于水稻黑条矮缩病毒在水稻不同分蘖内的绝对含量仍不清楚。此外,病株不同分蘖上介体灰飞虱的获毒性差异亦不明确。为此,本研究拟测定水稻黑条矮缩病株中不同分蘖内及根部的病毒含量,旨在更进一步增强对RBSDV侵染水稻的认识,为研究因病毒侵染而引起的植物生理学上的变化奠定一定的理论基础。
1 材料与方法
1.1 供试材料
灰飞虱采集自江苏省海安市水稻田(119.02°E、33.62°N),经筛选后长期饲养于实验室。RBSDV水稻病株由带毒灰飞虱传毒获得,长期保存于江苏省农业科学院植保所实验室。接种水稻为迟熟中粳淮稻5号,该品种高感水稻黑条矮缩病。灰飞虱常规饲养选用迟熟中粳武育粳3号水稻苗。
1.2 试验方法
1.2.1 灰飞虱的饲毒及传毒。将2龄期无毒灰飞虱若虫在清洗干净的RBSDV水稻上饲喂2 d,后转移至健康无毒的武育粳3号水稻苗上饲养12 d,随后采用Elisa方法对灰飞虱带毒率进行检测。按每棵苗2头的有效接种虫量,将体内RBSDV度过循回期的灰飞虱接种到淮稻5号幼苗上。以无毒灰飞虱接种淮稻5号作为对照处理。接种3 d后,将水稻苗种植于田间水泥池内,记录种植日期,喷施吡虫啉,覆盖防虫网。
1.2.2 总RNA的提取。接种RBSDV 40 d后,取RBSDV侵染的水稻病株5株,根据分蘖数量,茎叶部 分 别 标 记 为RB1-L1至RB1-L5、RB2-L1至RB2-L6、RB3-L1至RB3-L5、RB4-L1至RB4-L5、RB5-L1至RB5-L5;根部分别对应标记为RB1-R、RB2-R、RB3-R、RB4-R和RB5-R。根据TaKaRa公司RNAiso Plus(9109)试剂说明书提取样品总RNA,利用NanoDrop 2000(Eppendorf)分光光度计测定RNA浓度,冻存于-80℃备用。
1.2.3 cDNA的合成及荧光定量PCR。参照TaKaRa公司反转录试剂(RR047A)使用说明书合成水稻及灰飞虱cDNA。设计RBSDV外壳蛋白P10基因部分序列PCR引物(F端引物:CATTGTCAAATCGCCCCACG,R端引物:TACCGCGCTCCAAGTTTGTT)对RBSDV毒株带毒率进行分析。以RBSDV水稻不同分蘖的cDNA为模板,利用RBSDV-P10定量特异性PCR引物[12](F端引物:GCCCCACGTTGCATCTTC,R端引物:TGTTGGGCAAAGTGCTAGTTTC),参照TaKaRa公司荧光定量试剂(RR066A)说明书,配制20μL反应体系,在IQTM5荧光定量PCR仪内进行3步法PCR反应。
1.3 数据分析
本研究所有数据均利用SPSS 19.0进行分析。多组数据间采用单因素方差分析,2组数据间采用t检验进行差异性统计分析。
2 结果与分析
2.1 水稻黑条矮缩病株的分蘖数
将接种了RBSDV的淮稻5号水稻种植于田间水泥池内,40 d后各取5株无毒灰飞虱接种的水稻及表现出典型RBSDV病害症状的水稻(图1-A),利用RBSDV特异性引物进行PCR检测。结果显示,与对照相比,在表现出典型RBSDV病害症状的水稻内可以检测到1条清晰的条带,表明RBSDV成功侵染水稻(图1-B)。

图1 RBSDV侵染水稻后的症状及PCR检测
接着,对RBSDV侵染40 d后水稻的分蘖数进行分析,每组3株,各处理取5组。t检验分析发现,RBSDV侵染后水稻分蘖数没有显著性差异(表1),表明RBSDV侵染后对水稻的分蘖数无影响。

表1 相同生育期内健康水稻和RBSDV水稻分蘖数
2.2 标准曲线的制定
为明确水稻黑条矮缩病株中不同分蘖内病毒的绝对含量,本研究首先将RBSDV外壳蛋白P10基因的开放阅读框(open reading frame,ORF)克隆至pEASY-Blunt Zero载体(全式金公司),筛选后将阳性克隆送至生物公司测序,取测序正确、无突变的质粒(质量浓度为358 ng/μL),将该质粒稀释为6个浓度梯度(358×10-3、358×10-4、358×10-5、358×10-6、358×10-7、358×10-8ng/μL),以不同浓度的质粒为模板进行荧光定量PCR,获得不同浓度对应的Ct值(表2)。以质粒浓度为横坐标,Ct值为纵坐标,获得标准曲线:y=-3.374 9x+33.622,其中相关系数R2=0.995,且当x=0时,Ct值为33.622,表明该标准曲线较合理,可以满足后续试验(图2)。根据以下公式计算不同浓度质粒对应的绝对含量(表2):
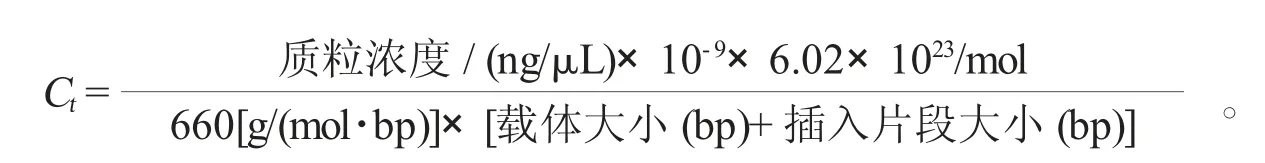

表2 不同稀释倍数对应的Ct值

图2 根据稀释倍数和Ct值制定的标准曲线
2.3 水稻黑条矮缩病株中不同分蘖内病毒的绝对定量
选取RBSDV病株不同分蘖的部分叶部和茎部(标记为L)、根部(标记为R)(图3),根据荧光定量PCR得到的Ct值,结合绝对定量计算公式和标准曲线公式,计算出各病株不同分蘖内及根部病毒的绝对含量(表3)。

图3 RBSDV水稻病株分蘖数标记及取样部位
利用单因素方差分析,对各病株平均病毒含量进行两两比较发现,接种RBSDV 40 d后,5棵RBSDV病株的平均病毒含量无显著性差异(3.83×105±1.48×105、4.23×105±9.15×104、4.17×105±1.50×105、6.28×105±1.89×105、3.41×105±1.23×105,均在105个copy/μg RNA),该结果表明同批次RBSDV的接种效率基本一致,对后续开展饲毒试验的影响较小。5棵病株不同分蘖内的病毒绝对滴度在104~106个copy/μgRNA,表明病毒在不同分蘖内存在一定的侵染差异,其中一级分蘖(L3)内的病毒含量(1.59×105±2.56×104copy/μgRNA)显著低于整株病毒含量的平均值(4.38×105±6.39×104copy/μg RNA)(P=0.000 78),该结果表明病毒更容易在侧分蘖内积累,导致分蘖更矮。此外,从表3中可以得出,RBSDV侵染水稻后,利用独立样本t检验分析发现,病毒在水稻根部的绝对含量(7.83×105±1.36×105copy/μgRNA)显著高于茎叶部的病毒含量(3.71×105±5.39×104copy/μgRNA)(P=0.035)。

表3 水稻黑条矮缩病株不同分蘖内病毒的绝对含量
3 讨论与结论
截至目前,我国先后报道了11种水稻病毒病[13],它们在不同时期对我国的粮食生产带来了巨大的威胁。其中,有10种病毒由介体飞虱或叶蝉以持久增殖型的方式进行传播,6种来自于呼肠孤病毒科,2种来自于弹状病毒科,2种来自于白细病毒科。在这11种水稻病毒中,有8种侵染水稻后出现典型的矮化症状。研究表明,病毒侵染后引起植物内源激素发生变化,进而促进病毒侵染,导致植株矮化,如水稻矮缩病毒(rice dwarf virus,RDV)的结构蛋白P2与赤霉素合成关键蛋白贝壳杉烯氧化酶互作,抑制赤霉素的合成从而导致植物矮缩[14]。部分病毒侵染后引起水稻分蘖增多,如由水稻草矮病毒(rice grassy stunt virus,RGSV)引起的水稻草矮病,病株分蘖急剧增多,呈杂草状。有研究报道RGSV侵染后引起的分蘖增多可能与独脚金内酯(strigolactones,SL)和生长素(indoleacetic acid,IAA)等激素变化相关[15-16],然而目前关于RGSV分蘖增多的具体的机制仍不明确。本研究发现,与无毒灰飞虱接种后的水稻相比,RBSDV侵染淮稻5号40 d后,水稻分蘖数没有明显增多或减少,推测RBSDV侵染水稻后未引起SL的变化。
接种RBSDV 40 d后,水稻植株明显矮缩。通过计算发现,随机5棵RBSDV病株内病毒含量基本一致,表明同一批次接种RBSDV的效率相对一致,该结果对后续开展水稻黑条矮缩病的抗性鉴定、介体灰飞虱与病毒的互作研究具有一定的指导意义。病株不同分蘖内病毒的绝对滴度存在一定差异,其中一级分蘖内的病毒含量显著低于整株的病毒含量,病毒在水稻不同分蘖内积累的机制值得深入探究。水稻分蘖的产生过程包括分蘖芽的形成以及分蘖芽的伸长[17],RBSDV如何侵染分蘖芽,以及在何时侵染新生分蘖组织尚不清楚。水稻分蘖芽顶端分生组织是否具有干细胞重要调节因子WUSCHEL(WUS)[18],进而抑制病毒对分生组织的侵染,也值得进一步深入研究。
笔者研究发现,RBSDV侵染水稻后,病毒在根部的绝对含量高于整株的平均病毒含量,推测RBSDV从茎叶部转移至根部后,更容易在水稻根部复制和增殖,然而这其中的机制尚不明确。中国小麦花叶病毒(Chinese wheat mosaic virus,CWMV)仅可在相对低温时成功侵染烟草,而在24℃却不能侵染;Andika等研究发现,沉默烟草根部特异表达的RNA依赖的RNA聚合酶(NbRDR6)后,仅可以在根部检测到CWMV,且NbRDR6可以抑制CWMV的积累以及抑制病毒源小RNA的产生[19]。RBSDV侵染水稻后根部的某些基因是否被诱导,进而有利于病毒的侵染,值得进一步探究。植物根部通常位于地表下面,负责吸收土壤中的水分和无机盐,具有支持、繁殖、贮存合成有机物的作用,大量病毒在根细胞内积累势必会引起根发生一系列的生理变化。然而RBSDV如何影响水稻根部正常的生理功能,进而影响水稻的生育周期,目前仍不清楚。本研究揭示了RBSDV在水稻不同分蘖及根部的绝对滴度,可以加深人们对RBSDV侵染水稻的认识,有望为研究病毒与寄主植物的互作提供一定的理论依据。
