沙门氏菌生物被膜的结构、调控因子与控制措施研究进展
2022-01-10闫玉卿董鹏程张一敏毛衍伟梁荣蓉朱立贤
闫玉卿,董鹏程,张一敏,毛衍伟,梁荣蓉,朱立贤 ,罗 欣,2
(1. 山东农业大学食品科学与工程学院,山东泰安271018;2. 江苏省肉类生产与加工质量安全控制协同创新中心,江苏南京210095)
沙门氏菌(Salmonellaspp.)是导致食源性疾病的重要致病菌之一,在肉类食品中的污染率较高[1]。据欧盟食品安全局(EFSA)统计,沙门氏菌近年来一直是欧盟的第二大常见的人畜共患病致病菌,并且大多数来源于动物性食品[2-3]。据估计,沙门氏菌每年在全球会造成9 380万人感染和15.5万人死亡[4]。2009—2015年,美国由沙门氏菌引起的疾病暴发了896次、23 662例,在由单一确诊病因的暴发疾病中占到30%,位居第二[5]。2014年,欧盟报告的确诊人类沙门氏菌病的病例就高达88 175例,导致了9 830人住院治疗和65人死亡[6]。2008—2015年,我国食品中毒事件中,因微生物导致的食物中毒事件占62.02%,而在所有的致病菌中,沙门氏菌占比为23%,居于首位[7]。综上所述,沙门氏菌是一种容易引起高爆发率疾病的食源性致病菌,威胁着人类的健康。
生物被膜(biofilm)是一种复杂的多糖蛋白复合物,它由在生物或非生物表面附着的细菌以及细菌分泌的胞外聚合物(EPS)组成[8]。胞外聚合物包括多糖、蛋白质、脂质以及细胞外DNA等,它们使细菌相互黏附并附着在物体表面[9]。沙门氏菌能够在不同器物的接触表面形成生物被膜,如玻璃、不锈钢、聚乙烯等;鼠伤寒沙门氏菌还可以在肉汤的气-液界面形成生物被膜[10]。生物被膜一旦形成,对各种杀菌剂和环境变化的敏感性大大降低,与浮游菌相比,生物被膜对抗菌素的抗性更强,这使得清除食品加工设施上的生物被膜成为一个巨大的挑战[11],传统的杀菌措施不能够有效地将其去除,从而造成持续污染[12]。有研究统计,生物被膜对农产品加工、乳制品、水产、畜禽加工和即食食品等工业都会产生一些不利影响[13]。尤其在肉类工业中,致病菌和腐败菌形成的生物被膜会成为加工产品的持续污染源,导致严重的卫生问题和经济损失[14]。
1 沙门氏菌生物被膜的主要结构组成与调节因子
1.1 生物被膜的主要结构成分
1.1.1 蛋白类成分
卷曲菌毛(curli fimbriae)是一种由csgBAC-csgDEFG操纵子编码的富含蛋白质的丝状物质,具有很强的黏附性,在细菌的运动和定殖等过程中非常重要。卷曲菌毛在生物被膜的形成过程中可以促进细菌在表面的初始黏附以及细菌间的相互作用。有学者通过构建卷曲菌毛相关基因csgA与csgD的缺失菌株,研究它们在沙门氏菌生物被膜形成过程中的作用,结果发现这两个基因缺失的沙门氏菌完全不能形成生物被膜,从而印证了卷曲菌毛在生物被膜形成过程中起着重要的作用[15]。此外,卷曲菌毛还会影响沙门氏菌的表型,研究发现具有完整卷曲菌毛的沙门氏菌在含有刚果红的培养基上呈现典型的RDAR(红色、干燥和粗糙)形态,然而未能产生完整的卷曲菌毛会导致刚果红培养基琼脂平板上出现PDAR(粉红色、干燥和粗糙)形态[16]。除卷曲菌毛外,沙门氏菌表面另一种常见的菌毛——Ⅰ型菌毛(type Ⅰ fimbriae)在细菌黏附和生物被膜形成过程中同样起到重要作用。Ⅰ型菌毛由fim基因簇(fimA~H)编码,其主要结构亚基FimA由基因fimA编码,由fimH基因编码的结构亚基FimH可与宿主细胞上含甘露糖的糖蛋白受体结合而介导细菌黏附[17]。
与菌毛不同,鞭毛是一种长螺旋状可旋转的多蛋白组装体,作为细菌的运动器官位于细菌细胞表面,使细菌能在液体培养基中游动,并群集于固体表面,是细菌单一泳动和群集泳动中不可或缺的组件。鞭毛组件的主要调节蛋白包括FlhD、FlhC、FliA、FliZ、FlgM、FliD和FliT等,而控制鞭毛基因表达的启动子分为三类:1类启动子(PflhDC启动子)编码2种蛋白质FlhD和FlhC;2类启动子控制构成钩基体(HBB)、丝帽蛋白(FliD)和调节蛋白(FliA、FlgM、FliZ和FliT)等结构元件的表达;3类启动子由FliA激活,正向调控自身表达以及编码FlgM、FliZ、FliD和FliT的基因表达[18]。鞭毛素为鞭毛的主要结构亚单位,沙门氏菌中鞭毛素由单一的fliC基因编码。有学者研究发现鞭毛素编码基因fliC缺失后,肠炎沙门氏菌成膜能力下降一半左右[19]。
生物膜相关蛋白(BAPs)是细菌在黏附过程中的重要蛋白,也存在于成熟的生物被膜中。BapA是肠炎沙门氏菌形成生物被膜所必需的第二大蛋白,由位于bapA基因下游的Ⅰ型蛋白分泌系统(BapBCD)分泌。Latasa等[20]研究发现,基因bapA的缺失会导致生物被膜形成能力的丧失,而bapA的过表达增加了生物被膜的形成能力;他们还发现卷曲菌毛的过量产生可以弥补bapA突变株的生物被膜缺陷。
沙门氏菌常通过一般分泌途径(GSP)来分泌蛋白,如菌毛。沙门氏菌利用GSP同源物分泌可溶性蛋白(如纤维素酶、果胶酶、蛋白酶等),它们可以分解大分子物质供细菌利用。此外,沙门氏菌还可利用GSP分泌转运蛋白,GspD是一种完整的外膜蛋白,它是由12种单体组成的多聚体复合物,可以转运大分子。GSP蛋白在分泌功能上具有高度特异性,不同细菌来源的蛋白不能相互替代[21-22]。
1.1.2 胞外多糖
纤维素(cellulose)是一种由bcsABZC-bcsEFG基因编码的β-1,4-D葡萄糖聚合物,它是胞外多糖的重要组成部分,可以支持长距离的细胞间相互作用,从而产生黏性质地,在生物被膜形成过程中起重要作用。Solano等[23]进行的bcsC和bcsE基因的非极性突变以及互补实验表明,这2个操纵子均在肠炎沙门氏菌和鼠伤寒沙门氏菌中负责纤维素的生物合成。真核细胞的细菌黏附和侵袭试验以及缺乏纤维素的突变体的体内毒力研究表明,纤维素的产生与肠炎沙门氏菌的毒力无关。但是,缺乏纤维素的突变体对氯处理更敏感,这表明纤维素的产生和生物被膜的形成可能是肠炎沙门氏菌在表面环境中存活的重要因素。
脂多糖(lipopolysaccharide)是介导沙门氏菌生物被膜形成的关键组分,rfaG基因编码的葡糖基转移酶和rfaJ基因编码的1,2-葡糖基转移酶都参与脂多糖生物合成[24]。Kim等[25]研究发现:rfbA基因也会影响脂多糖,当生物被膜在肉汤、禽肉以及各种材料上形成时,rfbA突变会使脂多糖发生异常。此外,rpoN基因在细菌应激反应中起着至关重要的作用,它负责胞外多糖的产生,被认为与生物被膜的形成有关,该基因受到sigma因子调控[26]。荚膜异多糖酸是一种荚膜细胞外多糖,它有助于沙门氏菌在上皮细胞上形成三维结构[27]。然而这种胞外多糖对于非生物表面沙门氏菌生物被膜的形成不是必需的。
生物被膜中的各个组分之间既相互作用又相互影响,共同维持生物被膜的完整性和稳定性。Gibson等[28]研究发现,正常的卷曲菌毛产量会使纤维素的产量降低,而卷曲菌毛与纤维素之间的产量平衡在一定程度上又受到脂多糖的影响。胞外DNA(eDNA)是胞外聚合物中的核酸成分,有促进生物被膜形成、参与遗传信息传递以及抗菌等作用[29-31]。eDNA的释放由糖基酶和外膜蛋白磷脂酶A的活性触发[32],而eDNA又与胞外多糖、胞外蛋白以及一些代谢产物结合,可保持EPS结构完整性。
1.2 沙门氏菌生物被膜中的主要调节因子
1.2.1 CsgD
CsgD是一种由csgD基因编码的转录调节蛋白,作为沙门氏菌生物被膜形成的主要控制和整合单元,可调节特定沙门氏菌生物被膜相关基质化合物的表达。Grantcharova等[33]使用csgD基因突变菌株在玻璃上进行生物被膜实验,表明CsgD是生物被膜成熟所必需的,且在微菌落形成中是必不可少的[33]。CsgD可以正向调控卷曲菌毛和纤维素的合成,如图1所示。
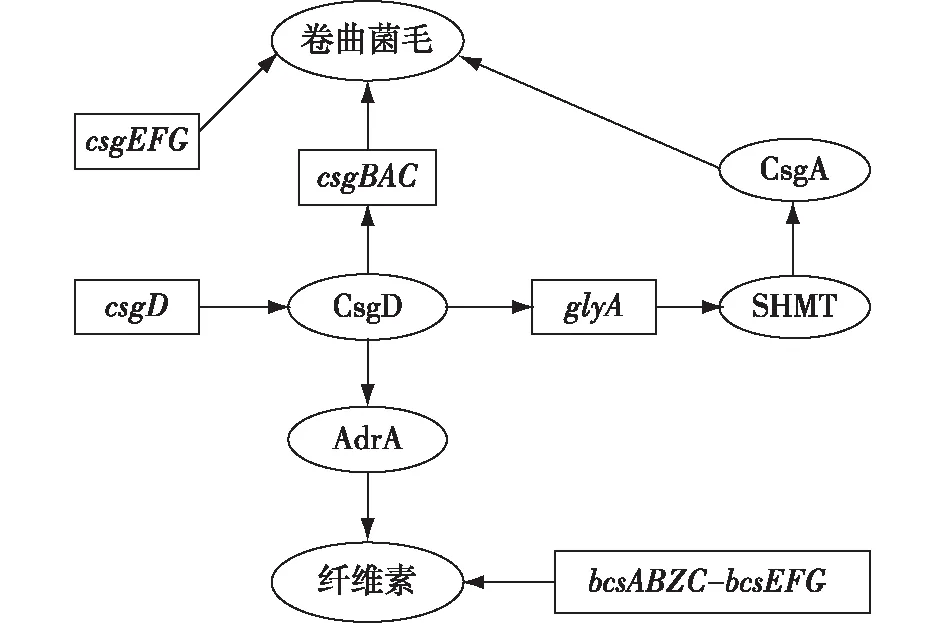
图1 CsgD对卷曲菌毛和纤维素的调控Fig.1 Regulation of CsgD on curli fimbriae and cellulose
CsgD通过调节csgBAC编码的结构性卷曲菌毛亚基的转录来控制卷曲菌毛的合成,CsgA作为编码卷曲菌毛的主要子单元也受到CsgD的调控。CsgD上调了glyA基因的表达,glyA编码的丝氨酸羟甲基转移酶(SHMT)负责甘氨酸的生物合成。CsgA的N端是一个富含甘氨酸的氨基酸区,因此SHMT活性的上调促进了CsgA的合成,从而促进生物被膜的形成。此外,csgC与csgEFG的基因产物对卷曲菌毛的产生具有辅助作用。纤维素的生物合成也受到CsgD的正向调控。CsgD刺激AdrA(膜结合的具有双鸟苷酸环化酶活性的GGDEF结构域蛋白,该蛋白参与环二鸟苷酸(c-di-GMP)的合成)的转录,AdrA通过与纤维素合成操纵子bcsABZC-bcsEFG的基因产物相互作用促进纤维素的合成。此外,AdrA产生的环状核苷酸是纤维素合成的活化剂[34]。
CsgD还能够调节O-Ag荚膜,这种荚膜由操纵子yihU-yshA和yihVW合成并转运,是EPS的另一种重要组分。CsgD通过抑制调节因子YihW来激活yihU-yshA,并与其他细胞外基质化合物协同作用,对yih操纵子进行差异调节[28]。综上所述,CsgD可被视为生物被膜控制点,调节所有主要沙门氏菌生物被膜成分的表达,并控制浮游菌和生物被膜之间的过渡。
1.2.2 c-di-GMP
c-di-GMP是一种重要的细菌第二信使,它能对多种细胞外信号作出反应,还可以控制动物和植物病原菌的毒力,参与某些细菌的细胞周期控制[35]。c-di-GMP的产生和降解分别由鸟苷酸环化酶(GGDEF结构域蛋白)和磷酸二酯酶(EAL或HD-GYP结构域蛋白)完成。AdrA是第一个被识别的对沙门氏菌生物被膜形成起重要作用的GGDEF结构域蛋白,在纤维素生物合成中具有调节作用。当adrA基因过表达时,环境中的c-di-GMP水平会升高。Simm等[36]使用CR/CF分析和细胞c-di-GMP定量分析,研究显示AdrA通过c-di-GMP的转录后,会对负责纤维素生物合成的bcs基因发挥作用。然而当编码EAL结构域蛋白的yhjH基因过表达时,环境中的c-di-GMP水平降低;且研究发现STM3611和STM1827(EAL结构域蛋白)参与细胞c-di-GMP水平的下调,从而导致纤维素产量降低,生物被膜形成减少。
通常,高c-di-GMP水平会刺激各种黏附素和生物被膜相关的胞外多糖的产生,并降低鞭毛的表达和活性[37]。Kader等[38]研究发现,c-di-GMP浓度升高,除了激活纤维素的生物合成外,还通过增强CsgD和CsgA的表达来增强卷曲菌毛的产生。同样地,当环境中CsgD和CsgA的表达量较高时,c-di-GMP的浓度也会提高,再次激活CsgD的表达,最终形成一个正反馈循环。当c-di-GMP与鞭毛的转录调节因子FleQ结合时,会使该蛋白的构象发生变化,从而无法调节下游的鞭毛基因。此外,Ha等[39]研究发现,当c-di-GMP水平升高时,YcgR蛋白会与鞭毛的转子蛋白FliG以及定子蛋白MotA相互作用,使得运动能力降低。
1.2.3 群体感应信号分子
群体感应(QS)是细菌通过分泌化学信号并对化学信号作出反应而相互交流的过程,群体感应在毒性、抗生素耐药性、生物被膜形成和运动性方面起到重要作用。群体感应系统由自诱导物、群体感应信号分子和调控蛋白组成。
在沙门氏菌中,QS机制常通过自诱导因子(AI)介导,即AI-1、AI-2。革兰氏阴性菌使用的是AI-1通信系统,其群体感应系统的信号分子N-酰基高丝氨酸内酯(AHL)由LuxI型酶(AHL合成酶)在一定环境下合成。AHL可以通过细菌膜自由扩散,通过特定的通道到达细胞外,并在环境中积累。当环境中的AHL浓度达到一定阈值时,AHL自动进入细胞并与细胞内LuxR结合,引起细菌细胞特异性功能基因的表达[40]。Chorianopoulos等[41]研究发现,在牙槽菌培养的无细胞上清液中存在AHL,对沙门氏菌生物被膜的发育有负面影响,但是沙门氏菌的这个系统是不完整的,因为它不含有编码AI-1合成酶的同源LuxI的基因,因此细菌无法产生自己的AHL信号。然而,沙门氏菌存在一种称为SdiA的蛋白质,这是一种与LuxR同源的转录调节因子,可以响应由其他微生物产生的、能够自由通过细胞膜扩散的AHL[42]。
AI-2是革兰氏阳性菌和革兰氏阴性菌中都存在的群体感应信号,可以调控生物被膜的形成、黏附能力等。随着细菌数量的增加,AI-2在细胞外膜中逐渐积累,最后通过在Lsr操纵子中合成的Lsr转运体将AI-2导入细胞内膜[8]。AI-2的合成、分泌与信号传递受到luxS基因调控的操纵子调控。AI-2的前体为4,5-二羟基-2,3-戊二酮(DPD),它是S-腺苷蛋氨酸(SAM)代谢的副产物,由luxS基因编码的LuxS酶是SAM分解代谢过程中的关键酶[43]。周文渊等[44]研究发现,肉桂醛可以抑制沙门氏菌的AI-2的活力,从而减少生物被膜的生成。
2 沙门氏菌生物被膜形成的影响因素
2.1 菌株特性
沙门氏菌生物被膜的形成能力与其菌株自身的特性有关,例如血清型的不同、基因的表达状况不同以及胞外聚合结构的表达等。Wang等[45]在研究从牛肉中分离的肠道沙门氏菌菌株的生物被膜形成能力时发现,不同的菌株之间生物被膜的形成能力具有高度的菌株特异性,并且与卷曲菌毛和纤维素的产生密切相关。Yin等[46]在研究沙门氏菌的生物被膜时发现,来源于中国肉牛屠宰厂的8个血清型、70多株沙门氏菌中,不同血清型的沙门氏菌形成生物被膜的能力不同,其中山夫登堡沙门氏菌(Salmonellasenftenberg)和金斯敦沙门氏菌(Salmonellakingston)形成生物被膜的能力较强,而同种血清型中不同的菌株生物被膜的形成能力也存在差异。此外,不同菌株之间泳动能力、自聚性能等的差异也会影响生物被膜的形成。
2.2 接触面特征
当沙门氏菌在材料表面黏附时,接触面的特征是影响生物被膜形成的重要因素,其中包括材料的不同、接触表面粗糙度以及表面疏水性等。王虎虎等[47]研究沙门氏菌生物被膜在不同食品接触面的转移率时发现,生物被膜在不同材料上的转移率差异显著,其中,生物被膜在不锈钢、玻璃以及聚乙烯表面的转移率较大。生物被膜在不同材料上的转移率与材料的表面疏水性、粗糙度以及电荷量等有关。Dhowlaghar等[48]使用扫描电子显微镜观察食物接触面上沙门氏菌生物被膜的形成状况,结果发现,与不锈钢、聚乙烯和聚氨酯表面相比,生物被膜在buna-N橡胶表面的含量最少,而此种橡胶的表面疏水性强、表面能量低、粗糙度强。Nguyen等[49]研究发现伤寒沙门氏菌可以附着在不锈钢和丙烯酸材料的表面,但在不锈钢表面的附着性更强。这是因为不锈钢是亲水的,与疏水表面相比,细菌更容易附着在这些表面。Benhamed等[50]研究发现,疏水材料比亲水材料更不易附着细菌。
2.3 环境因素
环境因素是影响生物被膜形成的又一个重要因素,其中包括环境的温度、pH、生存环境中的营养成分以及离子含量等。温度是影响生物被膜形成的重要因素,温度对生物被膜形成的影响依赖于生物被膜形成阶段:在指数阶段,随着温度的升高,生物被膜的数量增加;而一旦达到稳态,则出现相反的趋势;此时随着温度的升高,生物被膜的形成速率增加,而生物被膜的形成总量随着温度的升高而减小[51]。
Iliadis等[52]在研究低营养条件下温度、pH和NaCl浓度对肠炎沙门氏菌和鼠伤寒沙门氏菌的影响时发现,NaCl含量的增加限制了生物被膜的生长,在低NaCl浓度下,pH从4.5增加到7.4对生物被膜的形成起到了有利作用;同时也验证了肠炎沙门氏菌生物被膜形成的最适温度为34.5 ℃。O′Leary等[53]研究酸性环境对伤寒沙门氏菌DT104分离株生物被膜形成和基因表达的影响时发现,与培养基pH为7的环境相比,分离株1481在培养基pH为5时生物被膜形成的相关基因csgD和mlrA以及毒力相关基因hilA和invA表达上调。此外,营养物质是生物被膜形成过程中的关键物质,Paz-méndez等[54]研究发现,与牛奶汁、番茄汁和鸡肉汁相比,在营养成分更高的大豆胰蛋白酶培养基中的沙门氏菌成膜率更高。
2.4 其他微生物的影响
在实际的生产过程中,食品器具等的表面微生物种类繁杂,生物被膜常常以混合生物被膜的形式存在,沙门氏菌生物被膜的形成受到环境中其他微生物的影响。Gómez等[55]发现乳酸乳球菌、弯曲乳杆菌等在48 h内会对沙门氏菌的生物被膜完全抑制。这些乳酸菌主要通过与潜在的病原体混合或产生抗微生物物质和抑制细菌黏附的生物表面活性剂来抑制生物被膜[56]。Petrova等[57]证实了从益生菌株鼠李糖乳杆菌GG中分离到的凝集素样分子对包括沙门氏菌在内的多种病原体形成的生物被膜具有明显的抑制作用。
竞争与合作是混合生物被膜菌落中主要的细菌相互作用形式。能分泌生物表面活性剂、多糖或酶(如核酸酶和蛋白酶)的细菌,可减弱细菌与表面和细菌与细菌之间的相互作用,并减少细菌定殖和生物被膜的形成,或增加细胞的分散。同时,生物被膜中的细菌相互作用也可以促进细菌定殖和生物被膜的形成。研究发现,双歧杆菌对沙门氏菌具有生物拮抗作用,其通过分泌酸降低pH从而降低病原菌的竞争,或通过对营养物质和定殖位点的竞争来影响沙门氏菌的生长[58]。Pang等[59]将肠炎沙门氏菌与铜绿假单胞菌共培养,结果发现,混合生物被膜中沙门氏菌的数量明显低于单菌种生物被膜,这是由于铜绿假单胞菌的拮抗作用。此外,混合生物被膜对氯的耐受能力增加,荧光显微镜图像显示,混合生物被膜中存在较多的多糖复合物,这可能是肠炎沙门氏菌耐氯能力增强的原因之一。Chen等[60]研究发现,沙门氏菌与大肠杆菌O157:H7共培养后,2种菌相互拮抗且对加入混合生物被膜中的乙酰丙酸更敏感。
多种微生物间的交互作用使得生物被膜的形成比单一菌种更复杂,在不同的阶段,生物被膜间的进化是动态的。目前其他微生物与沙门氏菌共存时对沙门氏菌生物被膜的影响尚不明确,且沙门氏菌与其他微生物共存对消毒剂的敏感性或耐受性也会受到影响,但相关机制尚不清楚,该方面的研究是今后关注的方向之一。
3 沙门氏菌生物被膜的控制
3.1 消毒剂
消毒剂在食品加工厂中常用来抑制和清除食品器械表面的生物被膜,其主要通过杀死细菌、减少活菌数来控制生物被膜的形成。屠宰加工过程中常见的消毒剂有含氯消毒剂、H2O2、过氧乙酸等[11]。氯、乙醇和乳酸等消毒剂对于沙门氏菌生物被膜都有抑制作用,一般来说,这些消毒剂的抗菌活性取决于其本身浓度。在高浓度时,氯会腐蚀设备,乙醇易燃;而低浓度下,细菌可能会存活并产生诱导耐酸响应,产生更强的耐酸性;当乙醇类消毒剂与氯、乳酸联合使用时,在较低浓度下就能达到较高的抗菌活性[61]。
将消毒剂与其他影响生物被膜的物质联合使用往往会得到更好的抗生物被膜效果,Vestby等[62]用合成溴化呋喃酮对常规消毒剂(如次氯酸盐和苯扎氯铵)在消毒前进行预处理,结果显著提高了消毒剂的致死效果,因为溴化呋喃酮减少了沙门氏菌生物被膜的形成,生物被膜的结构发生变化而更易接受后续的抗菌处理。Lamas等[63]研究发现,将防腐剂添加到培养基中会对一些与生物被膜形成相关的基因产生影响:在含亚硫酸钠(SS)、亚硝酸钠(SN)、乙酸钠与柠檬酸混合物(SA)的培养基中,基因csgD显著下调。而csgD编码的生物被膜主要调节因子CsgD负责编码卷曲菌毛以及通过adrA和c-di-GMP调控纤维素的合成;在SS、SA培养基中,群体感应相关基因sdiA显著下调;鼠伤寒沙门氏菌中的毒力基因invA的转录也在3种培养基中下调。Kim等[64]使用热处理、紫外线、H2O2和乳酸等几种在食品工业中广泛应用的干预措施来干扰伤寒沙门氏菌在非生物表面的初始附着时发现,在食品加工过程中,受伤的细胞有很多机会存活并附着在非生物表面,热、紫外线和H2O2只能损伤细胞但对其附着性能影响不显著,而乳酸处理能有效降低附着在非生物表面的细菌数量,从而有效地延缓其初始附着和生物被膜的形成。因此可以考虑,将物理方法与消毒剂两者有机结合起来,会获得更好的抑制效果。
1990年7月,在美国巴尔的摩召开的第一届国际纳米科学与纳米技术大会上“纳米技术”(Nanoscaletechnology)这一新概念被提了出来。而所谓纳米药物就是指通过一定的微细加工方式对某些具有药用价值或其他的物质直接进行操作,形成新的具有纳米尺度的物质或载体[1]。纳米药物主要包括纳米晶、纳米乳、纳米球、纳米囊、脂质体和胶束等[2]。像脂质体、胶束等纳米药物相关技术已经成熟,而且已有上市产品,如力朴素等。
3.2 群体感应抑制剂
AI-2是革兰氏细菌中常见的一种自诱导物,沙门氏菌可以通过AI-2调节毒力因子的产生和生物被膜的形成。LsrA是一种ATP结合蛋白,在转运AI-2方面起着重要作用。展青霉素(patulin)具有AI-2的结构类似物,是一种群体感应抑制剂。
Vijayababu等[65]研究发现,30 μg/mL的patulin对伤寒沙门氏菌生物被膜的形成有明显的抑制作用,patulin与LsrA蛋白的关键残基通过氢键相互作用,阻断了AI-2的转运。贾坤等[66]研究发现,铜绿假单胞菌产生的AHL信号分子对鼠伤寒沙门氏菌生物被膜具有抑制作用,且随着AHL添加量升高,抑制作用增强。c-di-GMP是存在于沙门氏菌中的一种信号分子,它能够调控生物被膜的形成。Li等[67]研究发现,邻氨基苯甲酸盐对肠炎沙门氏菌生物被膜具有抑制作用,它通过降低细胞内c-di-GMP水平并增强泳动能力来抑制生物被膜的形成。
3.3 生物控制措施
噬菌体是一种能侵染细菌并影响生物被膜形成的病毒,它能够直接攻击生物被膜内的细菌,而不攻击胞外聚合物,是一种高效的生物学抑制方法。此外,噬菌体侵染EPS边缘的一些细菌,导致噬菌体越来越多。生物被膜上细菌的减少导致EPS物质的消减,最终将生物被膜完全去除[68]。Ferreira等[69]研究发现,使用溶菌噬菌体对不锈钢上的肠炎沙门氏菌生物被膜进行降解时,在第8天使用107 PFU/mL的噬菌体处理生物被膜35 min清除效果最好。
酶是对特定化学分子具有催化活性的蛋白质,它可以有效且靶向去除生物被膜。单一的酶不能有效地破坏生物被膜结构,但是当其与表面活性剂结合时可以提高酶的去除效率[70]。生物表面活性剂是一种可以降低液体和固体表面张力的化合物,它们穿透固体基质与生物被膜之间的界面并吸附在界面上,通过降低界面张力使得细菌表面和固体表面之间的吸引力作用减少,使得生物被膜更易去除[71]。
此外,利用沙门氏菌与其他细菌的拮抗作用也能实现对沙门氏菌生物被膜的控制。乳酸菌等益生菌可以通过影响毒力基因的表达、生物被膜的形成和群体感应来抑制病原体。由革兰氏阴性菌和革兰氏阳性菌产生的细菌素是核糖体合成的抗菌肽,对其他细菌具有抗菌活性[72]。Vandeplas等[73]研究发现,细菌素和产生细菌素的细菌对沙门氏菌具有一定的控制能力。
3.4 其他控制措施
一些从植物中提取的活性成分、新型化学材料等也能有效抑制生物被膜。精油是从植物中提取的芳香族油性液体,以其抗菌活性闻名。Oh等[74]在实验中将牛至、百里香酚、香芹酚精油加入培养沙门氏菌生物被膜的96孔板中,最后发现,3种精油对沙门氏菌生物被膜的形成均有抑制作用,且香芹酚精油和百里香酚精油的抑制效果更好。
银纳米粒子有较好的抑菌效果,但是银纳米粒子本身是不稳定的。Ag@AgCl是把银纳米粒子和其他材料杂化在一起的一种更加稳定的新型材料,将其用来处理鼠伤寒沙门氏菌生物被膜,获得了很好的抑制效果,Ag@AgCl是通过抑菌和降低细菌黏附性能来抑制生物被膜的形成[75]。
血根碱是一种天然的植物源性抗菌物质,王静慧等[76]研究发现,沙门氏菌生物被膜所产的胞外多糖在血根碱处理后有明显的减少,从而证明了血根碱可通过抑制沙门氏菌产生胞外多糖来抑制生物被膜的形成。唐玉霞等[77]研究发现,牛蒡叶的34%乙醇洗脱成分对沙门氏菌生物被膜具有抑制作用,其中的绿原酸和槲皮素等活性成分对沙门氏菌的细胞疏水性和泳动能力产生影响,从而抑制了生物被膜的形成。
4 结论
在食品工业中,食品及其加工器械上的生物被膜增加了病原微生物传播的风险,导致食品被污染,进而威胁消费者的健康。随着对致病微生物的深入研究,对沙门氏菌生物被膜的形成过程及其机制、影响因素、控制清除措施的研究也更全面。
沙门氏菌生物被膜的形成过程非常复杂,生物被膜的形成不仅取决于菌体特性和遗传基础,还取决于环境因素,包括温度、pH、表面特性、营养成分和背景菌群等。
目前对生物被膜的抑制主要使用抗生素和消毒剂,然而生物被膜对其具有极高的耐药性,因此寻找新的和有效的策略来控制它变得至关重要。随着对生物被膜形成的分子机制、群体感应信号通路的研究越来越深入,从分子层面通过抑制细菌毒力、限制耐药性、抑制生物被膜调节因子等相对应的干预措施来抑制沙门氏菌生物被膜将成为今后的研究热点。
