四倍体水稻蛋白质含量和谷蛋白基因表达研究
2022-01-10隆艳喜罗雁茹竺正航廖芷依
隆艳喜,罗雁茹,竺正航,廖芷依,王 兰
(华南农业大学 农学院,广东省植物分子育种重点实验室,广东 广州 510642)
水稻是我国最主要的粮食作物之一,其播种面积约占总种植面积的30%[1]。随着生产的发展、人民生活水平的提高,以及农产品对外贸易的需要,人们不只是关注水稻的产量,水稻的品质也成为关注的热点[2-3]。水稻的品质包括淀粉含量、蛋白质和氨基酸的含量及组成等[4]。水稻胚乳中的淀粉是通过几种酶的协同活性合成的,水稻基因组至少携带8个功能性α-淀粉酶基因[5]。淀粉通常由直链淀粉和支链淀粉组成,直链淀粉含量决定了淀粉的回生和糊化[6]。有研究发现,OsNAC20和OsNAC26双突变可以降低淀粉和贮藏蛋白含量,导致直链淀粉含量较低[7-8]。水稻胚乳蛋白位于淀粉体附近,影响淀粉的消化率[9]。水稻种子中的蛋白质主要以贮藏蛋白的形式存在于胚乳中,根据各贮藏蛋白的溶解性不同,分为清蛋白、球蛋白、醇溶蛋白、谷蛋白等4个组分。清蛋白、球蛋白主要储存于糊粉层和胚中,醇溶蛋白出现在PB-Ⅰ中,而谷蛋白储存于胚乳PB-Ⅱ中[10]。PB-Ⅰ和PB-Ⅱ有不同的起源;PB-Ⅰ起源于内质网网膜,PB-Ⅱ起源于液泡;这说明水稻醇溶蛋白和谷蛋白的合成场所和形成过程是不同的[11]。在胚乳蛋白中谷蛋白占70%以上,它对稻米品质的优劣及营养价值起着重要作用。就其氨基酸结构而言,大米蛋白质是所有其他谷物蛋白质中最优选的蛋白质。谷蛋白富含赖氨酸,其含量在谷物中位居第二,是谷物中的限制性氨基酸[12]。
同源四倍体水稻是由二倍体水稻经过染色体组加倍获得的种质,具有生长势强、生物学产量高、籽粒大、米质好和抗逆性强等特点,被列为21世纪水稻育种新战略之一[2]。同源四倍体水稻胚乳蛋白各组分含量与相应的二倍体相比,同源四倍体水稻胚乳蛋白含量大部分呈增加趋势,各组分分布与其二倍体水稻相一致,即趋同于二倍化[13-14]。因此,基因组多倍化对同源四倍体水稻重复基因组进化最主要的影响是在蛋白质表达量上的差异,这种差异对水稻育种有着积极的意义[11]。由于同源四倍体水稻减数分裂时期同源染色体联会及分离没有规律性,导致同源四倍体水稻育性偏低、结实率下降[15]。但在胚乳发育过程中,同源四倍体水稻及其二倍体原种胚乳发育的基本过程无差异,发育进程存在差异[16]。同源四倍体的胚乳蛋白随着基因剂量的增加而增加,主要表现在谷蛋白上,胚乳蛋白亚基无变化,一些胚乳差异蛋白也被找到[13-14,17]。四倍体杂种是由2个同源四倍体水稻杂交而来,具有更强的生物学优势与杂种优势[18]。肖波等[19]研究发现,同源四倍体水稻广陆矮4号与L202-4x杂交,其自交后代连续多年结实率都保持在80%左右。Guo 等[20]通过对同源四倍体水稻及其杂种F1不同组织的转录组差异进行研究,找到了影响四倍体水稻育性和杂种优势的相关基因[21]。对多倍化水稻DNA甲基化变异与基因组的研究表明,二倍体水稻和四倍体水稻的大部分基因的表达水平没有差异[22]。迄今,现有的对胚乳蛋白的研究多局限在低结实率的同源四倍体水稻上,在生产上的应用价值不高,而对于高产四倍体杂种的胚乳蛋白的研究还处于空白。
华南农业大学农学院植物种质创新与细胞生物学实验室对矮脚南特四倍体(AJNT-4x)连续自交52代,选育出结实率比较高的同源四倍体水稻;同时对同源四倍体水稻Jackson-4x(T45)和96025-4x(T44)杂交,再连续自交25代,选育出结实率接近二倍体水稻的新型四倍体水稻华多1号(H1)[23]。本研究选用高结实率的同源四倍体水稻AJNT-4x与新型四倍体水稻H1为研究对象,对四倍体水稻的氨基酸、直链淀粉、蛋白含量以及谷蛋白相关基因表达等进行深入系统的研究,旨在了解不同染色体组成与品质的关系,为四倍体水稻优质育种提供理论依据。
1 材料和方法
1.1 试验材料
供试水稻材料为新型四倍体华多1号(H1)、美香占2号(MXZ2)、象牙香占(XYXZ)、矮脚南特二倍体(AJNT-2x),自交选育52代的矮脚南特同源四倍体(AJNT-4x)。于2019年晚季种植于华南农业大学实验农场,正常田间管理。从水稻幼穗发育期开始,对试验材料进行标记和观察。用记号笔标记当天开花的材料,连续标记一个月。摘取标记开花后5,6,7,8,9,10,11,12,15,18,20,21,24,27,30 d的材料,将其迅速装入冰盒,然后切胚,液氮速冻,放置-80 ℃冰箱保存备用。
1.2 农艺性状调查
试验材料于2019年晚季种植在华南农业大学教学农场。田间试验采用随机区组,株行距18 cm×21 cm,单株种植,每小区种植100株。田间管理同一般大田,力求均匀一致。成熟期各小区去除边行,测量20株的结实率、粒长、粒宽、千粒质量,均为3次重复[24]。
1.3 品质分析及氨基酸测定
参照郑明等[25]的方法,用近红外谷物分析仪InfratecTM1241对试验材料H1、MXZ2、XYXZ、AJNT-2x、AJNT-4x进行品质分析,每份样品重复取样3次,每次取样扫描3次,记录数据。用德国Sykam全自动氨基酸分析仪S-433D对AJNT-2x、H1、AJNT-4x花后15,20,30 d的17种氨基酸含量进行测定,参考周梦怡等[26]的方法。
1.4 胚乳蛋白的逐级提取和总蛋白提取
参照Kumamaru等[4]的水稻全蛋白和胚乳蛋白的提取方法,略做修改。将准备好的AJNT-2x、AJNT-4x及H1花后5,6,7,8,9,10,11,12,15,18,20,21,24,27,30 d切胚的胚乳分别磨成粉,准确称取100 mg干粉于1.5 mL离心管中,加入1.0 mL试剂(50 mmol/L Tris-HCl(pH 值6.8)+0.5 mol/L NaCl),剧烈振荡5 min,使样品悬浮混匀。在低温摇床4 ℃振荡6 h后,于4 ℃ 10 000 r/min离心10 min;重复上述步骤2次,合并3次上清即为清蛋白和球蛋白组分。在沉淀中加入1.0 mL试剂B(60%正丙醇),剧烈振荡5 min,使样品充分混匀。在低温摇床4 ℃振荡6 h后,于4 ℃ 10 000 r/min离心10 min;重复上述步骤2次,合并3次上清即为醇溶蛋白组分。在沉淀中加入1.0 mL试剂C(1%乳酸),剧烈振荡5 min,使样品充分混溶。在低温摇床4 ℃振荡6 h后,于4 ℃ 10 000 r/min离心10 min;重复上述步骤2次,合并3次上清即为谷蛋白成分。
1.5 蛋白含量测定
将逐级提取的蛋白样品进行1∶3稀释(样品∶磷酸缓冲盐溶液(1×PBS)),4种蛋白的含量采用BCA法进行测定,具体测定方法采用BCA 法蛋白质浓度测定试剂盒(生工生物公司)说明书进行操作,使用BioTek公司的Epoch2微孔板分光光度计酶标仪测定。
1.6 不同时期RNA的提取
选取H1开花后第5,7,9,10,12,15,18,20,24,30天的胚乳籽粒,对谷蛋白基因GluA-1、GluA-2、GluA-3、GluB-1、GluB-2、GluB-4和GluB-6、GluC-1、GluD的RNA表达水平进行研究(表1)。籽粒总RNA的提取参照杜何为等[27]方法。
1.7 谷蛋白基因的qRT-PCR分析
反转录用翌圣生物公司第一链cDNA合成试剂盒Hifair®Ⅱ1st Strand cDNA Synthesis Kit(gDNA digester plus),荧光定量用翌圣生物公司Hieff®qPCR SYBR Green Master Mix(High Rox Plus),扩增程序为:预变性95 ℃ 5 min;变性95 ℃ 10 s,退火/延伸60 ℃ 10 s,40个循环。
1.8 数据处理
用Graph pad整理试验数据并作图,利用SPSS软件进行数据分析,分析各组显著性差异。
2 结果与分析
2.1 AJNT-4x与H1的遗传组成与产量相关性状分析
AJNT-4x为AJNT-2x加倍并连续自交52代选育而成的高产同源四倍体水稻。华多1号(H1)为Jackson-4x(T45)和96025-4x(T44)杂交,再连续自交25代选育而成的高产新型四倍体水稻杂种。由表2可以看出:AJNT-4x的粒长明显比AJNT-2x的增长且差异显著,粒宽、粒厚与AJNT-2x的也存在显著差异;AJNT-4x的千粒质量比AJNT-2x的明显增加,且差异显著;AJNT-4x的结实率为49.94%,比自交第一代同源四倍体水稻的结实率明显提高,但仍然不及其原种AJNT-2x(71.18%);H1的粒长、粒宽、粒厚、千粒质量均不及AJNT-4x,但其结实率为74.29%,与AJNT-4x差异显著,与AJNT-2x接近,达到二倍体水平。
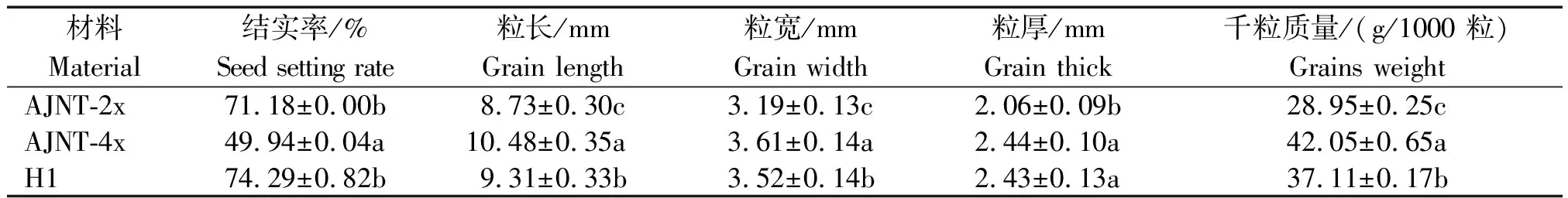
表2 AJNT-4x与H1的粒形性状分析Tab.2 The grain shape traits analyses for AJNT-4x and H1
2.2 AJNT-4x与H1的营养品质和食用品质分析
以国家级优质品种美香占2号(MXZ2)、象牙香占(XYXZ),以及矮脚南特二倍体(AJNT-2x)为对照,对AJNT-4x与H1的蛋白质含量与直链淀粉含量进行测定。结果表明:AJNT-4x糙米蛋白含量最高,为182.70 mg/g,优于国家优质品种MXZ2、XYXZ及其二倍体原种AJNT-2x,且差异显著;H1糙米蛋白含量比国家优质品种MXZ2、XYXZ略高,但不及AJNT-2x且差异显著。AJNT-4x的精米蛋白含量为115.70 mg/g,优于国家优质品种MXZ2、XYXZ,远远高于二倍体原种AJNT-2x且差异显著;H1的精米蛋白含量最低,仅71.30 mg/g,不及国家优质品种MXZ2、XYXZ(表3)。在直链淀粉含量方面,AJNT-4x精米直链淀粉含量为265.30 mg/g,优于二倍体原种AJNT-2x,远远高于国家优质品种MXZ2、XYXZ,且差异均显著;H1的精米直链淀粉含量高于国家优质品种MXZ2、XYXZ,但不及AJNT-2x(表3)。在碱硝值方面,AJNT-4x与H1的碱硝值相接近,分别为5.00,5.08,均低于国家优质品种XYXZ、MXZ2及AJNT-2x;在胶稠度方面,AJNT-4x的胶稠度为44.7 mm,远远低于国家优质品种MXZ2、XYXZ、及二倍体原种AJNT-2x,差异显著;H1的胶稠度为112.7 mm,介于国家优质品种MXZ2与XYXZ之间(表3)。

表3 AJNT-4x与国家优质品种MXZ2、XYXZ、二倍体原种AJNT-2x及新型四倍体H1 的品质分析Tab.3 Quality analysis for AJNT-4x,and national quality varieties of MXZ2,XYXZ,and AJNT-2x,and neo-tetraploid H1
这些结果说明,AJNT-4x的蛋白含量很高,但其胶稠度很低,其营养品质很高,高于国家优质品种,但食用品质却很差;H1的蛋白含量最低,但其胶稠度却很高,其营养品质偏低,但食用品质接近国家优质品种,口感好。
2.3 AJNT-4x与H1氨基酸积累的动态分析
比较分析AJNT-2x、AJNT-4x和H1花后15,20,30 d的17种氨基酸含量及人体必需氨基酸含量,结果表明:花后15 d,AJNT-4x与AJNT-2x、H1的17种氨基酸含量总量基本一致,但随着胚乳的发育,AJNT-4x和AJNT-2x 的总氨基酸含量逐渐升高,花后20 d达到最大值,且差异显著,AJNT-4x为91.70 mg/g,AJNT-2x为79.0 mg/g,然后略有下降并趋向稳定;而H1在花后15 d达到最大值,为65.60 mg/g,然后略有下降并趋向稳定,花后30 d为61.20 mg/g,与AJNT-2x(72.70 mg/g)差异显著(图1-A)。在花后15 d,H1的17种氨基酸总含量达到最大值,而AJNT-4x和AJNT-2x继续积累,直到授粉后20 d达到最大值。H1的17种氨基酸含量在发育过程中的变化趋势与总氨基酸含量的变化趋势基本一致(图1-B)。在氨基酸积累的动态过程中,人体必需氨基酸(苏氨酸、缬氨酸、蛋氨酸、异亮氨酸、赖氨酸)总量的变化趋势与17种氨基酸总量的变化趋势一致。花后30 d,AJNT-4x的17种氨基酸总量与人体必需氨基酸总量高于AJNT-2x与H1,H1的最低。由此推知,氨基酸含量积累过早停止导致H1在成熟期的总氨基酸含量下降,甚至低于AJNT-2x。

A.AJNT-2x、AJNT-4x和H1花后15,20,30 d的总氨基酸含量分析;B.H1花后15,20,30 d的17种氨基酸含量分析。不同字母表示在P =0.05水平的差异显著。A.Analysis for the total amino acid content of AJNT-2x,AJNT-4x and H1 on 15,20,30 days after flowering;B.Analysis for the seventeen amino acids′contents of H1 on 15,20,30 days after flowering. Different letters indicate significant differences at P =0.05.
2.4 胚乳蛋白含量积累的动态分析
花后5 d开始连续取材,直到花后30 d籽粒成熟,比较分析AJNT-4x、AJNT-2x及H1 4种胚乳蛋白含量积累的动态变化。结果表明:AJNT-4x与AJNT-2x、H1的醇溶蛋白与谷蛋白在胚乳整个发育过程中的积累趋势基本一致,都是在花后5 d已经积累。花后30 d,H1的清-球蛋白、醇溶蛋白含量都介于AJNT-2x与AJNT-4x之间,AJNT-4x的清-球蛋白、醇溶蛋白含量最高;而H1的谷蛋白含量(9.69 mg/g)却低于AJNT-2x(14.62 mg/g)与AJNT-4x(17.66 mg/g),差异显著,导致H1胚乳总蛋白含量(28.09 mg/g)低于AJNT-2x(31.04 mg/g)和AJNT-4x(37.76 mg/g)(图2)。说明H1总蛋白含量下降主要由胚乳谷蛋白含量下降引起。
在胚乳整个发育过程中,AJNT-4x与AJNT-2x、H1的清蛋白和球蛋白的波动一直比较平缓(图2-B)。AJNT-4x与AJNT-2x、H1的醇溶蛋白积累存在一个快速增长期,H1与AJNT-2x 都是从花后12 d开始快速积累,到花后15 d结束(H1为2.01 mg/g,AJNT-2x为1.77 mg/g),而AJNT-4x从花后12 d开始快速积累,到花后18 d结束(3.10 mg/g),含量远远高于H1与AJNT-2x,差异显著(图2-C)。AJNT-4x与AJNT-2x、H1的谷蛋白积累,从花后6~21 d,都存在快速增长期,但3种材料谷蛋白积累增长的时间和幅度略有差异。AJNT-4x、AJNT-2x存在3个快速增长期,AJNT-4x为花后6~7 d,9~11 d,18~21 d,AJNT-2x为花后7~8 d,9~12 d,20~21 d为快速增长期;H1从花后8~15 d持续缓慢增长,花后15~18 d为快速增长期(图2-D)。由于H1的谷蛋白积累稍为平缓,快速积累的时间比较短,导致成熟期H1的谷蛋白含量最少;AJNT-4x快速积累的时间稍早,导致AJNT-4x的蛋白含量比AJNT-2x高。由此推知,AJNT-4x的谷蛋白快速积累的时间比AJNT-2x、H1早,且快速积累的时间长,导致AJNT-4x的蛋白含量最高。
2.5 不同时期谷蛋白相关基因的表达分析
选取3个GluA、4个GluB、1个GluC和1个GluD共9个谷蛋白亚家族基因(表1),对AJNT-4x与AJNT-2x、H1在胚乳不同发育时期的表达进行研究。结果表明:花后5 d,AJNT-4x与AJNT-2x、H1的9个谷蛋白亚家族基因都有表达;随着胚乳的发育,各谷蛋白基因的表达量逐渐升高、再逐渐下降并趋于稳定,表达趋势基本一致,但不同的亚家族基因在相同的时间表达的丰度有差异。AJNT-4x中GluC-1表达最活跃,花后20 d达到峰值3 810.37,远远高于其他几个基因的表达峰值;成熟期花后30 d,AJNT-4x的9个基因的表达量均高于AJNT-2x和H1,尤其是GluC-1,在AJNT-4x中的表达量为1 404.12,远远高于AJNT-2x和H1。H1的3个谷蛋白基因GluA-1、GluA-2、和GluA-3的表达量的最大值远远低于AJNT-2x和AJNT-4x的最大值;达到最大值的时间也早于AJNT-2x和AJNT-4x,H1为花后12 d达到最大值,而AJNT-2x和AJNT-4x均为花后18 d达到最大值 (图3)。
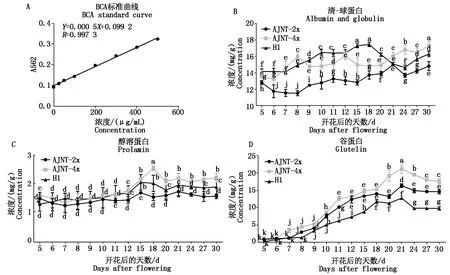
A.BCA法的标曲;B.清-球蛋白;C.醇溶蛋白;D.谷蛋白。以AJNT-2x为对照。每种蛋白不同的字母表示在P=0.05水平的差异显著。A.The standard curve of the BCA method;B.Albumin and globulin;C.Prolamin;D.Glutelin. AJNT-2x was used as control.Different letters in each protein indicate significant differences at P=0.05.
对于GluB基因而言,在AJNT-4x与AJNT-2x、H1 3个材料中,不同的基因在相同的时间表达丰度不同。GluB-1和GluB-4在H1中表达很活跃,GluB-1

每个基因不同的字母表示在P=0.05水平的差异显著。Different letters in each gene indicate significant differences at P=0.05.
在AJNT-4x与AJNT-2x、H1中分别为花后18,18,12 d达到最大值,在H1中表达峰值远远超过AJNT-2x和AJNT-4x,差异显著;而GluB-4在花后20 d,H1中的表达量也远远超过AJNT-2x和AJNT-4x,差异显著。GluB-2和GluB-6在胚乳不同的发育时期,在H1中的表达量均低于AJNT-2x和AJNT-4x,且在花后15 d达到最大值,而在AJNT-2x中为花后20 d达到最大值,在AJNT-4x中为花后18 d达到最大值。
相对于GluA、GluB、GluC而言,GluD的基因唯一,因此家族内不存在个别基因的差异。在表达高峰期,GluD在H1中的表达量远远低于AJNT-2x和AJNT-4x,差异显著。在3个材料中,GluD均在花后18 d达到最大值。
在胚乳发育过程中,不同的谷蛋白基因在相同的材料中达到峰值的时间不一样,相同的谷蛋白基因在不同的材料中达到峰值的时间也不一样。对于AJNT-4x而言,GluA-1、GluA-2和GluA-3均为花后18 d达到最大值,GluB-1、GluB-2和GluB-6均为花后18 d达到最大值,GluB-4为花后15 d达到最大值,GluC-1为花后20 d达到最大值,GluD为花后18 d达到最大值;对于H1而言,GluA-1、GluA-2和GluA-3均为花后12 d达到最大值,而GluB-1为花后12 d,GluB-2和GluB-6均为花后15 d达到最大值,GluB-4为花后20 d达到最大值,GluC-1为花后12 d达到最大值,GluD为花后18 d达到最大值。这些基因在不同的时期表达丰度的不同,导致谷蛋白含量的动态变化,并出现多个峰值。在不同的发育时期所检测的9个谷蛋白基因中,7个基因在H1中的表达峰值均远远低于AJNT-2x和AJNT-4x,导致H1谷蛋白含量下降;而它们在AJNT-4x中表达峰值均高于AJNT-2x和H1,导致AJNT-4x的蛋白含量最高。
3 讨论与结论
3.1 不同遗传组成与品质的关系
随着生物技术的发展,近几年国内外育种家利用多倍体杂交及连续自交多代选育技术,成功培育出一批高育性的四倍体水稻,其产量接近于二倍体水稻,克服了四倍体水稻育性低的问题[28-29]。但产量增加的同时,其品质有没有受到影响,值得探究。王成瑗等[30]通过对吉林省优质型、高产型和优质高产型共12个水稻品种的产量与稻米品质进行研究,发现稻米品质与产量呈负相关,产量提高后,除稻米的加工品质提高外,其他品质,如外观品质、营养品质及食用品质均下降。叶定池等[31]通过对产量构成因子每穗总粒数、实粒数、千粒质量等进行研究,发现大部分的产量构成因子都明显地影响稻米的品质,如实粒数与蛋白质含量呈显著的负相关。本研究对AJNT-4x及H1的品质进行深入系统的研究,发现AJNT-4x的结实率不及其二倍体原种AJNT-2x,仅49.94%,由此推知其产量明显下降,但其糙米与精米蛋白质含量分别为182.70,115.7 mg/g,远远高于其二倍体原种AJNT-2x,营养品质大大提高,而其胶稠度只有44.70 mm,远远低于二倍体原种,食用品质明显下降。而H1的结实率达到74.29%,接近二倍体水稻,由此可知其产量明显提高,但其糙米与精米的营养品质明显下降,远远低于AJNT-4x及AJNT-2x,而其食用品质远远高于AJNT-4x及AJNT-2x,接近国家优质品种。所以不能笼统地评价产量与品质的关系,因为品质分营养品质、加工品质、外观品质与食用品质等多方面,应该从不同的层次去评价产量与品质的关系。
营养品质包括蛋白质含量、氨基酸含量等,不同的研究材料,稻米蛋白质含量与产量的关系也不一样。目前存在2种观点,一种认为稻米产量与蛋白质含量呈负相关,即产量高蛋白质含量低;另一种认为稻米的产量与蛋白质含量呈正相关,即产量高蛋白质含量也高。本研究通过比较研究发现:H1的产量虽然提高了,接近AJNT-2x,但其精米中的蛋白质含量却下降了,低于AJNT-2x;AJNT-4x虽然产量不高,但其蛋白含量却提高了,远远高于H1及其二倍体原种AJNT-2x。由此推知,产量与蛋白质含量不存在正相关或负相关,产量或品质由材料本身的遗传组成决定。
3.2 胚乳蛋白的组成与含量,及谷蛋白相关基因的表达对稻米品质的影响
水稻的贮藏蛋白质主要存在于胚乳中,根据溶解性的不同,分为清蛋白、球蛋白、醇溶蛋白和谷蛋白。清蛋白与球蛋白主要集中在米糠与精糠中,在稻米加工碾磨过程中大部分丢失。精米中的蛋白质主要为醇溶蛋白和谷蛋白,所以这2种蛋白决定了稻米的食用品质和营养品质[32]。醇溶蛋白不能被肠胃消化,导致稻米的食味品质下降,而谷蛋白容易被人体消化和吸收,且富含赖氨酸、精氨酸等人体必需氨基酸,谷蛋白在水稻的品质改良中具有重大意义[12]。
花后30 d,与AJNT-2x、AJNT-4x相比,H1的胚乳清-球蛋白、醇溶蛋白含量位于AJNT-2x、AJNT-4x之间,而谷蛋白含量却远远低于AJNT-2x、AJNT-4x。谷蛋白含量的显著下降,导致H1的蛋白含量下降。也就是说H1蛋白含量下降是由于谷蛋白含量下降引起的。通过对胚乳的整个发育过程进行谷蛋白积累的动态分析,H1谷蛋白快速积累的时间比AJNT-2x、AJNT-4x均短,H1谷蛋白积累比较平缓,最终导致谷蛋白含量偏低。
谷蛋白是多基因家族,根据序列的相似性,分成GluA、GluB、GluC和GluD 4个亚家族[33]。总共15个基因,其中12个已被注释到这4个亚家族中,3个基因未命名但属于谷蛋白家族。谷蛋白基因的表达具有时空特异性,不同的材料在籽粒中表达的时期也有不同。孙红正等[34]对稻米贮藏蛋白的生物信息学分析,认为GluA-1与GluA-2在花后10 d的籽粒中表达量最高。Zhao等[35]的研究表明,GluA-3在种子发育前期开始表达,并于花后10~16 d达到峰值,其他谷蛋白成员在花后6 d才能检测到相应的转录本,花后14~16 d达到峰值。牛洪斌等[36]认为GluB-6在花后10~15 d达到峰值。本研究对AJNT-4x与AJNT-2x、H1的9个谷蛋白基因在胚乳的不同的发育时期进行研究,发现各谷蛋白基因在花后5 d均有表达。随着胚乳的发育,各基因的表达量均逐渐升高,达到峰值后,再逐渐下降并保持稳定。但各基因达到峰值的时间略有差异,与前人的研究相似。GluA和GluB、GluB-4、GluC亚族的8个基因均在花后12~20 d达到峰值,GluD在花后18 d达到峰值。谷蛋白基因的表达时空特异性,导致谷蛋白的含量在胚乳发育过程中出现多个峰值。李文静和孙艳香[37]对GluC-1的启动子进行研究,认为GluC-1是胚乳特异表达基因。本研究谷蛋白各基因的表达中,GluC-1在各时期的表达量最高,可能与GluC-1的胚乳特异启动子有关。在成熟期花后30 d,所研究的9个谷蛋白基因中,H1中谷蛋白基因的表达均远远低于AJNT-2x、AJNT-4x,这也是导致H1胚乳谷蛋白质含量下降的原因之一,最终导致稻米蛋白含量下降,影响稻米的营养品质。
3.3 氨基酸组成及含量对稻米品质的影响
稻米营养品质的高低,不仅取决于蛋白质含量,还与氨基酸的组成与含量,尤其是必需氨基酸含量有关[38]。人体摄取蛋白质主要是以氨基酸的形式进行吸收,必需氨基酸是人体自身不能合成或合成速度不能满足人体需求,必须从食物中摄取的,所以必需氨基酸是评价稻米营养品质的重要指标。张顺等[39]通过对28个精白米样品的必需氨基酸、总氨基酸及蛋白质含量进行分析,发现蛋白质含量与总氨基酸含量、必需氨基酸呈显著的正相关,总氨基酸含量与必需氨基酸含量也呈显著的正相关。本研究表明,必需氨基酸含量与17种氨基酸含量呈正相关,它们在胚乳发育过程中的变化趋势基本一致。花后15 d,H1必需氨基酸积累达到峰值,而AJNT-2x与AJNT-4x的必需氨基酸继续积累,花后20 d达到峰值。AJNT-4x的氨基酸总量与必需氨基酸总量高于AJNT-2x,是由于基因剂量的增加引起的;但H1的含量低于AJNT-4x,是由于H1的必需氨基酸积累过早,最终导致H1的总蛋白含量下降。牛洪斌等[40]认为,谷蛋白B亚族成员的赖氨酸等人体必需氨基酸含量普遍比A亚族的高,对增加稻米中GluB亚族成员含量有利于提高赖氨酸含量,改良稻米的品质。在本研究中,花后30 d,H1的GluB亚族成员4个中有3个成员(GluB-1、GluB-2、GluB-4)的表达均高于AJNT-2x,且与其他谷蛋白基因相比,H1与AJNT-4x的表达差距也最小。所以H1的蛋白含量虽与AJNT-2x、AJNT-4x差异显著,远远低于AJNT-2x、AJNT-4x,但H1的赖氨酸含量却与AJNT-2x、AJNT-4x相接近。说明虽然H1的营养品质下降,但不影响它的营养价值。
3.4 结论
通过对同源四倍体水稻AJNT-4x及其原种AJNT-2x、国家优质品种MXZ2、XYXZ,以及新型四倍体H1进行蛋白含量、直链淀粉含量、碱硝值、胶稠度等品质分析,发现AJNT-4x精米与糙米的蛋白含量远远高于二倍体原种AJNT-2x、新型四倍体水稻H1、和国家优质品种MXZ2、XYXZ,AJNT-4x的营养品质明显提高,但其胶稠度却最低,食用品质最差;H1精米蛋白含量下降,低于国家优质品质,且远远低于AJNT-4x,营养品质下降,但其胶稠度接近国家优质品种,且远远高于AJNT-4x,食用品质接近国家优质品种。以AJNT-2x为对照,对AJNT-4x和H1的胚乳蛋白质含量、人体必需氨基酸及谷蛋白基因的表达进行研究,深入探讨四倍体水稻的营养品质差异。在胚乳发育过程中,AJNT-4x的清-球蛋白、醇溶蛋白及谷蛋白含量最高,而H1的清-球蛋白、醇溶蛋白介于AJNT-2x与AJNT-4x之间,H1的谷蛋白含量最低,H1蛋白含量下降是由谷蛋白含量下降引起的。在胚乳整个发育过程中,H1谷蛋白的含量变化一直比较平缓,且谷蛋白快速积累的时间比较短。不同的谷蛋白基因在胚乳发育的不同时期表达量不一样,同一谷蛋白基因在不同的材料中表达的峰值不一样。所研究的9个谷蛋白基因中,花后30 d,只有3个基因(GluB-1、GluB-2和GluB-4)在H1中表达量高于AJNT-2x,所有的基因在AJNT-4x中表达量均最高。AJNT-4x的17种氨基酸含量与人体必需氨基酸含量最高,H1最低。主要是由于在胚乳发育过程中,H1的氨基酸积累在花后15 d达到峰值,而AJNT-2x与AJNT-4x在花后20 d达到峰值。基因剂量的增加导致AJNT-4x的氨基酸总量与必需氨酸总量高于AJNT-2x,但H1的氨基酸积累过早停止,导致它的氨基酸总量与必需氨基酸含量低于AJNT-4x,甚至低于AJNT-2x,最终导致H1的总蛋白含量最低。研究结果为水稻多倍体优质育种提供理论依据。
