喜盐鸢尾花部特征与交配系统研究
2022-01-08刘乐乐曹效东徐正茹张君芳吴永华
刘乐乐, 曹效东, 徐正茹, 张君芳, 吴永华*
(1.兰州市园林科学研究所, 甘肃 兰州 730070; 2.西北师范大学生命科学院, 甘肃 兰州 730070)
鸢尾属(Iris)植物全世界约有260~300种,生长于多种生境,主要分布于北半球温带地区[1],随着对其不断地开发,逐渐被应用于环境绿化、切花生产、医疗、日用化工等多个领域[2]。我国有丰富的鸢尾属植物资源,共64种,主要分布于西南、东北与西北地区[3]。西北地区鸢尾属植物有18种[3,4],很多种处于未开发状态。喜盐鸢尾(Irishalophila)是鸢尾科鸢尾属琴瓣鸢尾亚属多年生草本植物[4],分布于我国西北的甘肃、新疆地区,生境为草甸草原、山坡荒地、砾质坡地及盐碱地。目前对喜盐鸢尾的研究主要有生理抗性[5-6]2012,化学成分分析[7-8]2014。遗传多样性[9-11],种子休眠与萌发特性与种间杂交障碍[11-13];逆转运蛋白序列分析[14],花色形成关键基因的克隆及表达分析等[15]。
植物的繁育系统是指有性生殖过程中直接影响后代遗传组成的特征和行为系统,其中包括花部综合特征,花的各器官寿命、花的开放样式、传粉者种类和频率、自交亲和程度和交配系统等,这些与传粉者以及传粉行为一起决定了植物后代的遗传组成和适合度,其中花部综合特征与交配系统是核心[16-17]。鸢尾属植物由于具有丰富的色彩与多样性的花部结构,是研究进化生物学与物种形成的模式植物[18-20],目前国内对鸢尾属花部特征与交配系统的研究越来越多[1],有马蔺(I.lactea)[21]、鸢尾(I.tectorum)[22]、小花鸢尾(I.speculatrix)[23]、溪荪(I.sanguinea)[24]、膜苞鸢尾(I.bulleyana)[25]等,其交配系统都为异交,除膜苞鸢尾自交不亲和,其他鸢尾属植物均是部分自交亲和,虫媒传粉,主要的传粉昆虫有西方蜜蜂(Apismellifera)、中华蜜蜂(A.ceranacerana)、黄胸木蜂(Xylocopaappendiculata)与长木蜂(X.tranquebarorum)等,为这些植物的保护与利用提供了理论基础;但是对喜盐鸢尾花部特征与交配系统方面的研究却鲜有报道。喜盐鸢尾具有观赏性佳,抗性强的优良特性,在西北地区园林绿化、生态修复等方面有较大的应用价值[12],对其花部特征与交配系统的研究,不仅可以进一步加深对喜盐鸢尾繁殖生物学的认识,了解其对生态环境的适应性,还可以为其人工栽培,品种改良、杂交选育与合理的开发利用提供基础资料。
1 材料与方法
1.1 研究地点、材料与试验设计
以兰州市园林科学研究所鸢尾属植物种质资源圃的喜盐鸢尾居群为研究对象,该居群2012年由新疆引入,种植8年,面积约100 m2,共有喜盐鸢尾300余株,主要为淡黄色花。喜盐鸢尾为蝎尾状聚伞花序,在花茎分枝处有3枚草质绿色苞片,内有两朵花,小花从顶部苞片内第2朵花开始往上下两端逐步开放;喜盐鸢尾的花被片呈浅黄色、黄色或蓝紫色;花两性,有3个单元构成,每个单元包括1枚外花被、1枚雄蕊、1枚雌蕊与1枚内花被;总共有6枚花被片,成两轮排列,辐射对称,子房纺锤状圆柱形,子房中轴胎座,具3室。
研究地点的地理坐标为北纬36°7′1″,东经103°42′21″,海拔1 542 m,属于温带大陆性气候,年平均气温为9.3℃,年平均日照为2 446 h,无霜期180 d,年平均降水量325 mm,年蒸发量1 650 mm[26]。2020年4-5月,随机选择无病虫害,生长健壮的喜盐鸢尾植株进行观察与测试。
居群中设置4个区组:开花物候及花部特征观察的时候,每个区组观测花朵5个,共观测20朵花;在花粉活力与柱头可授性检测时,8个观测日期每个区组采集样品2个,共检测64朵花;在测量花粉/胚珠比时,每个区组采集样品5个,共测量20朵花;测量杂交指数中,每个区组选择10朵花,共测量40朵花;测量人工授粉试验中,一共有9个处理(包括1个对照),根据实际开花的情况,每个区组每个处理选择4~6个即将开放且发育正常的花蕾,一共177朵花。
1.2 开花物候及花部特征观察
观测居群花期,以居群25%个体开花时为始花期(First flowering date);50%的个体达到开花时视为群体开花高峰日期(Peak flowering date);95%的个体开花结束为终花期(Last flowering date)[27-28]。记录单株从第一朵花开放到最后一朵花闭合的天数;对每个花蕾进行标记,观察单朵花的开花动态,记录从花瓣开始展开到花瓣卷曲闭合的天数。用游标卡尺测量当天开花的花直径、外花被长与宽、内花被长与宽、花丝长、花药长、花柱长与宽;开花前1 d套袋,开花当天用直径0.2 mm硬质毛细玻璃管吸取并测定泌蜜量;用糖度计(WYT-A,0%~80%)测量花蜜的糖度。
1.3 花粉形态、活力与柱头可授性检测
1.3.1花粉形态检测 在植株盛花期收取自然开裂花药中的花粉,自然干燥,将花粉用导电胶粘到样品台上,放在离子溅射镀膜仪(JMB-3500VA)处理120 s,之后在Hitachi-S3400 N型扫描电子显微镜下观察。随机选取30粒花粉,选取有代表性视野进行拍照,对花粉粒的极轴长、赤道轴长以及表面纹饰进行测量与观察。
1.3.2花粉活力与柱头可授性检测 在2020年5月12日,在4个区组中分别选取处于不同开花日期(开花前2 d至开花后5 d)的花朵,一共采集8 d,每天采集8个,共计64朵花,采集花粉与柱头,测量花粉活力与柱头可授性。花粉经MTT溶液染色5 min后观察花粉粒的着色状况,根据花粉粒着色状况判断花粉活力,花粉粒变为蓝紫色表示有活力[29]。每朵花重复观察3个视野,并拍照。根据上述实验结果计算花粉活力,计算公式为:

采用联苯胺-过氧化氢法检测柱头可授性[30],将柱头浸入联苯胺-过氧化氢溶液[V(质量分数1%联苯胺)∶V(体积分数3%过氧化氢)∶V(蒸馏水)=4∶11∶22]中,当柱头被染为蓝色并伴有大量的气泡产生,说明柱头具有可授性,反之则无。
1.4 花粉/胚珠比(P/O)与杂交指数(OCI)的检测
在4个区组中分别选取次日开放(全部花冠从绿色苞片伸出),且长度基本一致的花蕾,每个花蕾随机选取一枚雄蕊,用HCl(1 mol·L-1)60℃水解1 h去除药壁,每个雄蕊的花粉粒悬浮液定容至10 mL,用微量移液器吸取10 μL悬浮液于显微镜下观察,统计每个雄蕊花粉量,取平均值X,则每个雄蕊的花粉数目1 000X。取子房在体视显微镜下用解剖刀剖开,统计胚珠数,重复20次,计算平均胚珠数。用平均每朵花的花粉量除以平均胚珠数得到花粉/胚珠比(P/O)。按Cruden[31]的标准,P/O为2.7~5.4时,其繁育系统为闭花受精;P/O为18.1~39.0时,繁育系统为专性自交;P/O为31.9~396.0时,繁育系统为兼性自交;P/O为244.7~2 588.0时,繁育系统为兼性异交;P/O为2 108.0~195 525.0时,繁育系统为专性异交。
按照Dafni的方法计算杂交指数(OCI)[32],并判断喜盐鸢尾的繁育系统类型。对选取的40朵花进行测量,花直径小于等于1.0 mm为0;1.0~2.0 mm为1,2.1~6.0 mm为2;大于6.0 mm为3。花药开裂时间与柱头可授期:雌蕊与雄蕊同时成熟或雌蕊先熟,为“0”;雄蕊先熟,为“1”;柱头与花药等高为“0”,柱头与花药在空间上分离,为“1”。上述3个指标相加值为OIC值。根据OIC值评判植物交配系统的评判标准为:0分,闭花受精;1分,专性自交;2分,兼性自交;3分,自交亲和,异交,需要传粉者;大于等于4分,部分自交亲和,异交,需要传粉者。
1.5 授粉试验
在2020年4月28日至5月2日进行人工授粉试验,不同区组的花蕾分别挂牌并做好标记。在开花前1 d(全部花冠从绿色苞片伸出)17∶00至19∶00分别对9组花蕾进行不同的授粉试验:T1组花蕾直接套袋,检测是否自交亲和;T2组花蕾去雄后套袋,检测是否无融合生殖;T3组花蕾去雄后不套袋,检测自然条件下异交在交配系统的比例;T4组花蕾去雄后套袋,次日9∶00至11∶00人工异株授粉,与T3结果结合检测自然条件下是否存在花粉限制;T5组花蕾套袋,次日9∶00至11∶00人工同株同花授粉,与T1结果结合检测雌雄异位、异熟在避免自交中有多大作用;T6组花蕾去雄后套袋,次日9∶00至11∶00人工同株异花授粉,检测同株异花授粉的亲和性;T7组花蕾剥去3枚外轮花被片不套袋,自然授粉,检测外轮花被片对交配成功的影响;T8组花蕾剥去全部花被片后不套袋,自然授粉,检测全部花被片对交配成功的影响;CK组花蕾自然授粉,以此组为对照组。在授粉后第14 d统计各组的坐果率、种子数和结籽率,计算公式分别为:



1.6 访花昆虫行为观察
在盛花期,9∶00~17∶00点,利用样方法观测访花昆虫,每2 h观察一次,每次观察10 min,记录样方内所有访花昆虫的种类、数量、访花时间与访花方式,并用照片记录,连续观察3 d。根据昆虫在访花过程中是否先后触碰到柱头的可授面与花药中散开的花粉达到异花传粉的效果,来判断其是否为潜在的传粉者。
1.7 统计分析
采用SPSS19.0统计分析软件对本实验的数据进行相关的统计分析。通过相关性分析(Correlation analysis)与线性回归分析开花前后的天数分别与花粉活力和柱头可授性比例之间的关系;用T检验来比较两个柱头之间的距离与外花被最窄处的宽度是否存在显著差异;用广义线性混合模型(GLMM)来分析授粉试验不同处理之间种子数与结籽率是否存在显著差异;统计数据用平均值±标准差表示。
2 结果与分析
2.1 花部特征
田间观察的结果表明:喜盐鸢尾花冠筒长约9.02 mm,呈圆柱形,分泌花蜜(图1A),花蜜分泌量为1.07 μL,糖度为27%。花直径79.70 mm,外花被3枚,淡黄色,长42.40 mm,宽12.91 mm,先端舒展为宽卵圆形;开放时自然下垂,至1/3处外花被两侧向上翻卷,呈提琴形;与上方两个柱头之间的接触点为外花被的最窄处,其中两个柱头之间的距离(4.06 mm)显著的大于外花被最窄处的宽度(2.49 mm,P<0.01);中脉处有黄色蜜导(图1A)。内花被3枚,淡黄色,倒披针形,基部边缘为黄色,长47.83 mm,宽11.99 mm(图1B)。花柱分枝扁平,3个,淡黄色,长231.90 mm,宽8.23 mm,顶端两个三角形裂片,两个柱头位于裂片的基部(图1B)。雄蕊长26.96 mm,花丝细长,花药黄色,向外开裂;花药长17.42 mm,位于两个花柱之间,柱头下方(图1B),花粉橙黄色(图1B)。子房高39.14 mm,纺锤状圆柱形,子房中轴胎座,具3室。
2.2 开花物候
喜盐鸢尾在5月初现蕾,居群始花期为5月6日至5月13日,盛花期为5月14日至5月20日,末花期为5月21日至5月27日,全部花期约为22 d,其中盛花期为7 d,单花花期3~5 d,单花序花期6~11 d,单株花期11~15 d。花朵在1 d中的开花时间不固定,白天全天都有开放。单花开放时,花冠先由绿色的苞片中伸出(图1C),接下来3个花单元分离(图1D),然后外花被与雄蕊、柱头整体与内花被上下分离(图1E),之后外花被顶端部分下垂(图1F),花朵开放后第2 d,3个花单元各自卷曲在一起(图1G),花期最后所有的花被萎缩卷曲在一起(图1H)。喜盐鸢尾在开花前期,雄蕊长于雌蕊柱头的顶端,雄蕊先成熟(图1I);开放后第2 d,雄蕊向上弯曲,顶端与柱头同样长,花粉散出具有活性(图1 J);第3 d,雄蕊中间继续向上弯曲,弧度与花柱一致,柱头向下弯曲,具有可授性(图1K);第4 d,花粉散落完,外花被萎缩卷曲(图1L)。

图1 喜盐鸢尾花朵开放动态
2.3 花粉形态、活力与柱头可授性
喜盐鸢尾的花粉均呈单粒,两侧对称,极面观为一裂圆形,赤道面观为椭圆形,花粉有一个萌发沟,极轴长约73.93 μm,赤道长约41.62 μm,极赤比(P/E)约为1.78(图2A);外壁为网状纹饰(图2B)。

图2 喜盐鸢尾花粉形态、外壁纹饰及花粉活力检测
根据测量,喜盐鸢尾的花粉花朵开放的前2 d,还没有从花药里散出时就已经有活力(图2C);结果显示在开花当天花粉活力最强,为(82.66±9.27)%,随着花朵开放的时间变长,花粉活力逐渐降低,到开花第5 d时,花粉活力为最弱,为(9.57±1.91)%;花粉活力随着天数的增加先升高后降低,与开花前后的天数呈现出显著的负相关关系(P<0.05,图3A)。柱头在开花前没有活力,开花0 d只有7.69%的柱头检测有可授性,开花1 d有25%的柱头有可授性,开花2 d时100%的柱头有可授性,之后柱头可授性降低,开花5 d柱头可授性比例为66.67%;柱头可授性比例随着天数的增加先升高后降低,与开花前后的天数呈现出显著的正相关关系(P<0.05,图3B)。
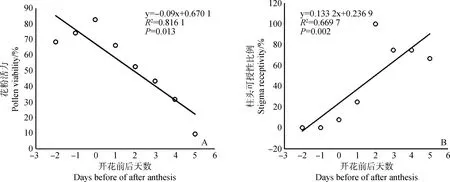
图3 喜盐鸢尾开花过程中花粉活力(A)与柱头可授性比例(B)变化
2.4 花粉胚珠(P/O)比与杂交指数(OCI)
喜盐鸢尾单花平均总花粉数为32 3750,平均总胚珠数为148.82,花粉胚珠比为2 175.47,根据花粉胚珠比判定交配系统为专性异交。单花直径为79.70 mm,大于6.0 mm,为“3”;雄蕊先熟为“1”;柱头与花药在空间上分离,为“1”,三者相加,喜盐鸢尾的杂交指数为5,根据杂交指数判定其交配系统为异交,部分自交亲和,需要传粉者。
2.5 授粉试验结果
表1显示喜盐鸢尾直接套袋(T1)结籽率与种子数显著低于对照组(P<0.05);人工同株同花授粉(T5)坐果率为100%,种子数与结籽率与对照组不存在显著差异;去雄后套袋(T2)坐果率与结籽率均为0.00%;去雄不套袋(T3)种子数与结籽率与对照组差异不显著;人工异株授粉(T4)与人工同株异花授粉(T6)种子数与结籽率显著高于对照组(P<0.05);剥去3枚外轮花被(T7)与剥去全部花被片(T8)的种子数与结籽率显著低于对照组(P<0.05),两者座果率(35%)比对照组(100%)低;在未完全成熟的绿色种荚(7个)中有昆虫幼虫啃食种子,占总数的3.95%。
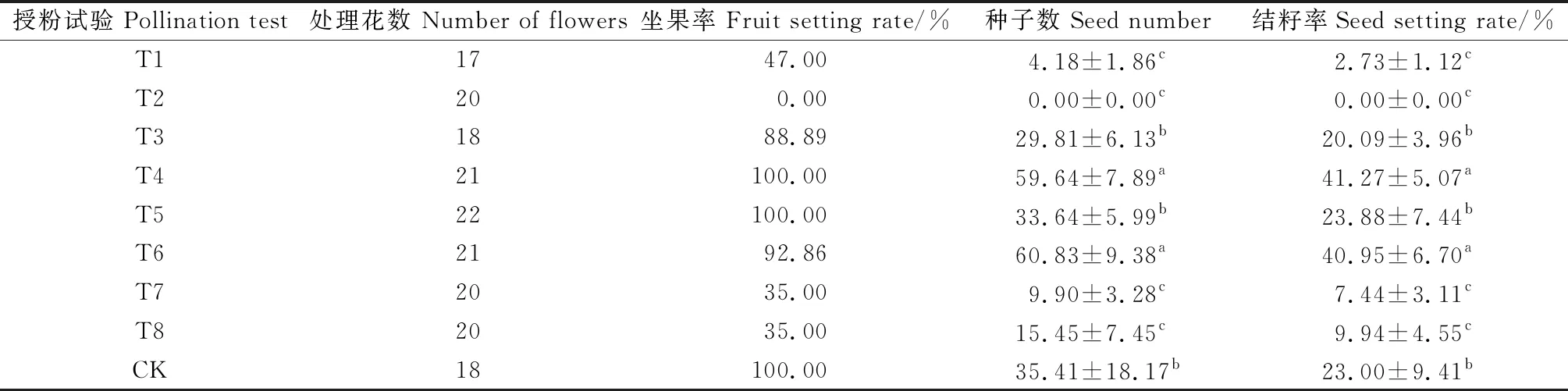
表1 喜盐鸢尾授粉试验结果
以上结果表明:喜盐鸢尾自交亲和,生殖过程以异交为主,需要传粉者,不存在无融合生殖,自然状态下存在花粉限制,花被片有助于提高种子数、结籽率与坐果率,有虫食现象。
2.6 访花昆虫的种类及访花行为
喜盐鸢尾的主要访花昆虫有:条蜂(Anthophorasp.)、意大利蜜蜂(A.melliferaligustica)与中华蜜蜂,三者的访花时间分别为13 s,34.67 s,13 s。虽然意大利蜜蜂的访花时间长于其它两者,但根据访花行为判断条蜂为潜在的传粉昆虫。访花过程中,条蜂的身体可以进入柱头与外花被之间的通道,头部首先与柱头的可授面接触,传递异花花粉,继续进入花冠筒基部吸食花蜜,此时花粉附着在条蜂的头部与身体,最后从通道出来,与柱头的不可授面接触,避免自花传粉(图3A-图3E)。中华蜜蜂(图3F)与意大利蜜蜂(图3G)为主要的访花昆虫,采集花蜜时,不接触花药与柱头,直接从外花被与柱头基部或者花冠与苞片的缝隙中伸入口器吸取花冠筒中的花蜜;采集花粉时,这两种昆虫不直接进入外花被与柱头之间传粉通道,只是从两者侧面的缝隙中探入身体采集花粉;因此判断两者并不是喜盐鸢尾的有效传粉昆虫。
在观测中还发现有淡脉隧蜂(Lasioglossumsp.)(图3H)、蚂蚁(图3I)、食蚜蝇、甲虫等访花昆虫,但数量较少;淡脉隧蜂的访花时间长达382 s,其体型较少,钻入未完全开放的花朵中盗取花粉;蚂蚁除了采食花蜜,也会采食花瓣;还观测到少量的蜘蛛,可能是访花昆虫的捕食者。

图4 喜盐鸢尾主要访花昆虫
3 讨论与结论
3.1 喜盐鸢尾的花部特征与传粉昆虫
鸢尾属中不同物种花冠的紧凑性,即瓣化的柱头与外花被之间传粉通道的大小与其传粉的泛化水平关联。花瓣结构较为松散的物种传粉昆虫种类丰富,例如变色鸢尾(I.versicolor)[33];花瓣结构较为紧凑的物种有效传粉昆虫较少,例如玉蝉花(I.ensata)[34],传粉通道两侧紧闭,有效传粉者为三条熊蜂与西方蜜蜂。喜盐鸢尾的传粉通道入口闭合,两侧有较窄缝隙,花冠紧凑;通过传粉通道采食花蜜与花粉的有效传粉昆虫只有条蜂;体型较大的意大利蜜蜂、中华蜜蜂与体型较小的蚂蚁、淡脉隧蜂,不通过传粉通道直接采集花粉或花蜜,会降低喜盐鸢尾接受有效传粉的几率。因此,通过其花部特征与传粉昆虫判断喜盐鸢尾具有特化的传粉系统。
花蜜与花粉是植物吸引传粉昆虫的两种主要报酬[35],也是植物重要的花部特征之一。喜盐鸢尾分泌与储存花蜜的器官为位于花朵基部的花冠筒,与其它无附属物亚属的鸢尾属植物一样,例如黄菖蒲、西伯利亚鸢尾、I.douglasiana等[36]。在对中国产鸢尾属部分物种系统发生学的研究中,喜盐鸢尾同样被嵌入到无附属物亚属的无附属物组[4]。喜盐鸢尾的花蜜含糖量为27%,与I.douglasiana花蜜含糖量(24%~28%)类似[36]。对鸢尾科植物射干的研究表明,其花蜜在1 d中不同时段的泌蜜量与含糖量不同,既与昆虫的访花活动相适应,也与环境因素有关(温度、湿度)[37]。本研究观察到喜盐鸢尾花未开放时就有花蜜从苞片中溢出,但没有对花蜜量与含糖量动态检测;其花粉颜色为橙黄色,与蜜导颜色一致,表面有网状的纹饰,有助于粘附到传粉昆虫的身体。访花昆虫中条蜂以采食花蜜为主,其余花粉与花蜜都采食。
喜盐鸢尾去除外花被或全部花被片后,种子数与结籽率都小于对照组,坐果率降低,这可能与外花被的黄色蜜导有关。蜜导在虫媒花中普遍存在,是一类对传粉者具有特殊吸引和指示作用的花图案或花结构,例如无叶美冠兰(Eulophiazollingeri)通过挥发香甜气味与唇瓣上黄色蜜导可以诱导绿彩带蜂进入花朵中觅食[38],但是蜜导是否会影响植物的雌性繁殖适合度还不确定,例如蓝翠雀花(Delphiniumcaeruleum)的蜜导黄色髯毛并不影响坐果率与结籽数[39]。因此喜盐鸢尾外花被的蜜导能否通过吸引更多昆虫访花从而提高雌性适合度还需要进一步的验证。
3.2 喜盐鸢尾的交配系统
根据花粉胚珠比判断喜盐鸢尾的交配系统为专性异交;根据杂交指数判断为异交型,部分自交亲和,需要传粉者;授粉试验的结果表明喜盐鸢尾以异交为主,部分自交亲和,需要传粉者,不存在无融合生殖。结合以上,判断其交配系统为兼性异交。
雌雄同体植物的雌性功能与雄性功能在时间上的异熟与在空间上的异位有助于促进植物异花授粉,从而避免近交衰退[40]。在时间上,喜盐鸢尾的雄蕊先熟,花粉在开花前2 d时已经有活力,在开花当天的活力最高,开花之后花粉活力逐渐降低;雌蕊后熟,柱头在开花前不具有可授性,开花后柱头可授性逐渐上升,在第2 d达到顶峰,之后雌蕊可授性的比例逐渐下降。在空间上,喜盐鸢尾雌雄异位,雌蕊成熟后,柱头向下弯曲90°,伸展在外花被的外侧;花药被向内翻卷的外花包裹,而无附属物鸢尾属植物的花药并不会被向内翻卷的外花被包裹。因此,喜盐鸢尾提琴状的外花被使其与无附属物鸢尾相比雌雄异位的机制有所不同。这种雌雄异位机制可能与喜盐鸢尾的花粉数量(323 750)较多有关,是同属的鸢尾(22 732)[10]、小花鸢尾(34 293)[23]、溪荪(51 176)[24]、膜苞鸢尾(61 080)[25]的花粉数量的6~14倍。较少的花粉散落的时间可能较短,例如溪荪的花粉2 h就能完全散尽[24];喜盐鸢尾花粉数量多,完全散尽所需时间可能更长,需要更有效的雌雄异位机制来防止自交。人工同株同花授粉的坐果率、种子数与结籽率均高于直接套袋,说明这种雌雄功能的时空隔离能够减少自花授粉,从而有利于促进异花授粉。
本研究中人工异花授粉的种子数与结籽率均高于对照组,说明喜盐鸢尾在自然条件存在花粉限制,这可能是因为有效传粉者不足有关。与大多数鸢尾属植物一样,喜盐鸢尾具有较强的克隆生长能力,即通过根状茎进行无性繁殖,作为一种生殖补偿机制来保证繁衍[10,40]。
引入地与原生境差异可能会对自然居群繁殖成功构成影响[41]。瑞士兰科手参属植物Gymnadeniaodoratissima的交互移栽试验表明低海拔的低地居群移栽到高海拔后繁殖成功率没有变化,但是高海拔的山地居群移栽到低海拔后繁殖成功率显著下降,同样海拔高度居群间移栽不会引起繁殖成功的变化,这可能是因为山地居群比低地居群的传粉者更加多样化[42]。南非石蒜科纳丽花属植物Nerinehumilis的交互移栽试验表明本地表型与引入表型相比产生更多的种子[43],这是因为本地表型能接受更多的花粉,单次访花产生更多的种子。本研究的喜盐鸢尾是引种成功的苗圃居群,其交配成功与原生境居群相比是否有差异以及原因有待进一步的研究。
4 结论
喜盐鸢尾5—6月开花,部分雌雄异熟,雌雄异位,交配系统以异交为主,部分自交亲和,不存在无融合生殖,需要传粉者。
