High level of extra-pair paternity in the socially monogamous Marsh Tits (Poecile palustris)
2022-01-07JuanWangYanhuiWeiLeiZhangYitingJiangKekeLiandDongmeiWan
Juan Wang, Yanhui Wei, Lei Zhang, Yiting Jiang, Keke Li and Dongmei Wan
Abstract
Keywords: Extra-pair paternity, Genetic compatibility hypothesis, Marsh Tits, Microsatellite
Background
Previous work based on molecular evidence shows that most socially monogamous birds follow a genetic polyandrous mating system (Griffith et al. 2002; Brouwer and Griffith 2019). By August 1st, 2017, nearly 76% of socially monogamous passerine species (255 species; 371 publications) exhibit cases of extra-pair paternity (EPP) (Brouwer and Griffith 2019). For instance, 75% of the nests have EPP and over 25.3% of offspring are not sired by the social father in Coal Tit (Parus ater
) (Lubjuhn et al. 1999).Study in African Blue Tits (Cyanistes teneriffae
) found that 39% of the nests have EPP and 15% of offspring are extra-pair young (Garcia-Del-Rey et al. 2012). Although the EPP rates of a seemingly large population have been reported, this only represents under 4% of the avian biodiversity. Even when considering the passerines, only 47% of families have been sampled (Brouwer and Griffith 2019). Furthermore, sampling is unevenly distributed with a larger representation of the birds from Western Europe and the eastern part of North America than the Asian area (Brouwer and Griffith 2019). Uneven sampling results in a less comprehensive insight into birds’ EPP behaviors. More studies on representative bird species from a broader range of regions are needed (Brouwer and Griffith 2019; Ding et al. 2021).Even though the advantage of EPP for females is debated, one potential benefit that females may obtain is an indirect genetic benefit by gaining compatible genes from extra-pair mate (Mitton et al. 1993; Brown 1997).Trivers (1972) suggests that females may accrue fitness benefits from choosing genetically dissimilar sires for their offspring. These genetically dissasortative mating preferences have been referred to as genetic compatibility preferences (Mays et al. 2008). The extra-pair fertilization of socially monogamous birds has provided an opportunity to understand the effects of genetic complementarity on mate choice. Under genetic compatibility hypothesis,females should prefer to choose males with lower genetic similarity than their social mates in extra-pair mating.This way, the extra-pair offspring will have higher genetic heterozygosis and higher adaptability than their withinpair half-siblings (Foerster et al. 2003). Yet the benefits on EPP or offspring fitness varies considerably across species. While in Red-backed Fairy-wrens (Malurus melanocephalus
), higher genetic heterozygosis and higher rate of extra-pair male siring have been observed on the young of females paired to genetically similar males (Varian-Ramos and Webster 2012), while in Varied Tits (Poecile varius
), relatedness and heterozygosis hardly showed correlation with the EPP (Ju et al. 2014). More field trials on mate choice are needed to test the genetic complementarity hypothesis.Marsh Tits (Poecile palustris
) are socially monogamous passerine birds, largely confined to mature temperate woodland (Hinsley et al. 2007). They are secondary cavity-nesting species and both sexes provide parental care during the nestling period. Here we used microsatellite molecular markers to test the paternity relationship and revealed the genetic mating system in the Marsh Tits population at the Xianrendong National Nature Reserve,Liaoning Province, China. In addition, we carried out an analysis on mate choice by comparing the genetic similarity between a female and her social partner. We also investigated female mating preferences by comparing the heterozygosis of maternal half-siblings to reveal the role of genetic compatibility on EPP of Marsh Tits.Methods
Study area and data collection
The study was done in the Xianrendong National Nature Reserve, Liaoning Province, China (122°53′24″–123°03′30″ E, 39°54′00″–40°03′00″ N) from 2012 to 2019.Within the study area, about 400 nest boxes (Length ×Width × Height = 16 m × 14 m × 34 cm) were installed on trees 2–2.5 m high from the ground. In March, adult Marsh Tits were caught with mist nets, and about 20 μL blood samples were taken from the brachial vein. From March to July, the nest boxes were visited every 1–3 days.When Marsh Tits bred in the nest boxes, the clutch size and nestling size were monitored. Parental birds were captured with nest-box traps when the nestlings were 6–8 days old (Zhang et al. 2019), and the nestlings were captured at 9–10 days old. Blood samples (about 20 μL)from all individuals were taken from the brachial vein and stored in ethanol. After being tagged by a numbered aluminum ring and a plastic color ring, all birds were released at the capture sites. GPS information of the sites were also recorded.
Laboratory work
DNA was extracted from the blood samples by TIANamp Genomic DNA Kit [Cat. DP304-03, Tiangen Biotech(Beijing) Co., Ltd.]. All samples were genotyped at nine microsatellite loci including two triplex (R34, R49), five tetraplex (R16, R24, R33, R47, R69), and two five-conjoined (R19, R75) (see in Additional file 1: Table S1). PCR reactions contained 0.5 μL of the DNA (30 ng/μL), 10 μL of the Premix Taq (5 U/μL), 0.5 μL of Primer R (10 pmol/μL), 0.5 μL of Primer F (10 pmol/μL), and 8.5 μL of ddHO. PCR amplifications were conducted under the following conditions: an initial denaturation step of 5 min at 94 °C followed by 30 cycles of 30 s at 94 °C, 30 s at 59 °C, 30 s at 72 °C, and final extension of 10 min at 72 °C.PCR products were separated by capillary electrophoresis on ABI3730XL Genetic Analyzer (Applied Biosystem)with ROX-500 as the internal lane size standards. Data were collected and analyzed using GeneMapper Software(Version 4.0, Applied Biosystem). Loci that could not be used to match offspring and their parents were re-genotyped to exclude genotyping error.
Paternity analysis and identification of extra-pair sires
Paternity assignments were performed in Cervus 3.0 software using maximum likelihood methods (Kalinowski and Taper 2006). Polymorphism parameters including allele number (N
), expected heterozygosity (H
),observed heterozygosity (H
), polymorphic information content (PIC), the cumulative exclusion probability of a false parent without genotype information of any true parent (Excl 1), the cumulative exclusion probability with the genotype of a known parent (Excl 2), Hardy–Weinberg equilibrium (HWE), and null allele frequencies(F
) were calculated. The PIC of the nine microsatellite loci were all greater than 0.5, indicating that all the nine microsatellites were highly polymorphic (see Additional file 1: Table S1). Parentage analysis revealed that all chicks in nests were genetic offspring of their associated social mothers, but not necessarily the offspring of the social fathers. The combined probability of exclusion for their genetic fathers of all loci was > 99.99% (see Additional file 1: Table S1).We searched for the most likely genetic father using inferred parental genotypes based on its genetic chicks and their associated social mother requested for assignments at 95% confidence level. Some nestlings were considered as extra-pair offspring with unknown fathers,because no male showed a good match with them.
Statistical analyses
Standardized heterozygosity (proportion of heterozygous loci/mean heterozygosity of typed loci) was chosen as individual heterozygosity (Ju et al. 2014). Pairwise relatedness (r
) were calculated as the genetic similarity between individuals, and the score of relatedness was proportion of gene sharing among individuals (a score of− 1 represents maximally difference between individuals,1 indicates clones, and 0 represents the average relatedness value between all males and females).Kolmogorove-Smirnov tests were used to determine whether the data was normally distributed or not. When the data was normally distributed, independent samplet
tests were used to test the genetic similarity between the social pairs with and without EPP, together with the standardized heterozygosity between the cuckolded social fathers and the males whose nestling has no EPP,and between the extra-pair offspring and their withinpair maternal half-siblings. Pairedt
tests were performed to test the difference in standardized heterozygosity between the cuckolding males and the males cuckolded them, and also the genetic similarity between the social mates and the extra-pair mates. If not, the data would be analyzed using non-parameter tests. Statistical analyses were performed in SPSS 21.0. All tests were two tailed with significance levels ofP
< 0.05. The values were expressed as the mean ± SE throughout.Results
Occurrence of EPP
A total of 87 males, 111 females, and 325 offspring from 44 nests were captured from 2012 to 2019. Forty-nine offspring (15.08%) were the results of extra-pair copulation (EPC), and 20 nests (45.45%) had at least one extra-pair chick. Among the 20 nests with EPP, the average number of extra-pair offspring was 2.45 ± 1.56 SE with the extra-pair offspring ratio varying from 11.11 to 71.43%. There was no case of full paternity loss. The average nestling size in the EPP nests (7.35 ± 1.42 SE) has with no significant difference from the nests without EPP(7.42 ± 1.38 SE; independent-samplet
test:F
= 0.00,P
= 0.99).Distance of extra‑pair sires
We successfully identified nine extra-pair males and calculated the distance from their social nests to the nest in which he sired young (Fig. 1). The distance varied from 40 to 2928 m with an average of 1066.22 ± 1048.35 m(n
= 9). Four of the nine nestlings were sired by neighbors (40–213 m), while the remaining five by extra-pair males nested far away from their young (689–2928 m).Genetic characteristics
We found no significant difference in standardized heterozygosity between the cuckolder and the cuckolded males (pairedt
test: correlation = 1.84,P
= 0.08),between the extra-pair young and within-pair maternal half-siblings (independent-samplet
test:F
= 0.02,P
= 0.14), or between the cuckolded social fathers and the males whose mate has no extra-pair copulation (EPC)(independent-samplet
test:F
= 0.30,P
= 0.62).No divergence was found in the genetic similarity between social mates with and without EPP (independent-samplet
test:F
= 0.377,P
= 0.53). There was no variation in relatedness from social mates to extra-pair mates (pairedt
test: correlation = 0.24,P
= 0.41) both from EPP nests.Discussion
Microsatellite loci, also known as simple sequence repeats, are important genetic markers revealing kinship, a comparative analysis of which is viable through genome-wide analyses (Huang et al. 2016). As these markers are highly variable, they are powerful tools to assign offspring to parents (Flanagan and Jones 2019).Through analysis on the microsatellite markers and maximum likelihood, we found that socially monogamous Marsh Tits participated in effective EPC: about 45.45% of the nests had at least one extra-pair offspring; 15.08% of the offspring were not genetically related to the putative father. The EPP behaviors of Marsh Tits were more frequently than the average socially monogamous birds(11.1% extra-pair offspring and 18.7% EPP nests) and many other passerines, such as Willow Tits (Parus montanus
, 6.7% extra-pair offspring and 21.4% EPP nests;Lampila et al. 2011), Great Tits (Parus major
, 5.4–8.6%extra-pair offspring and 27.8–44.2% mixed paternity nests; Lubjuhn and Strohbach 1999). Nine extra-pair fathers were successfully identified, four of whom were neighbor males while the others were not.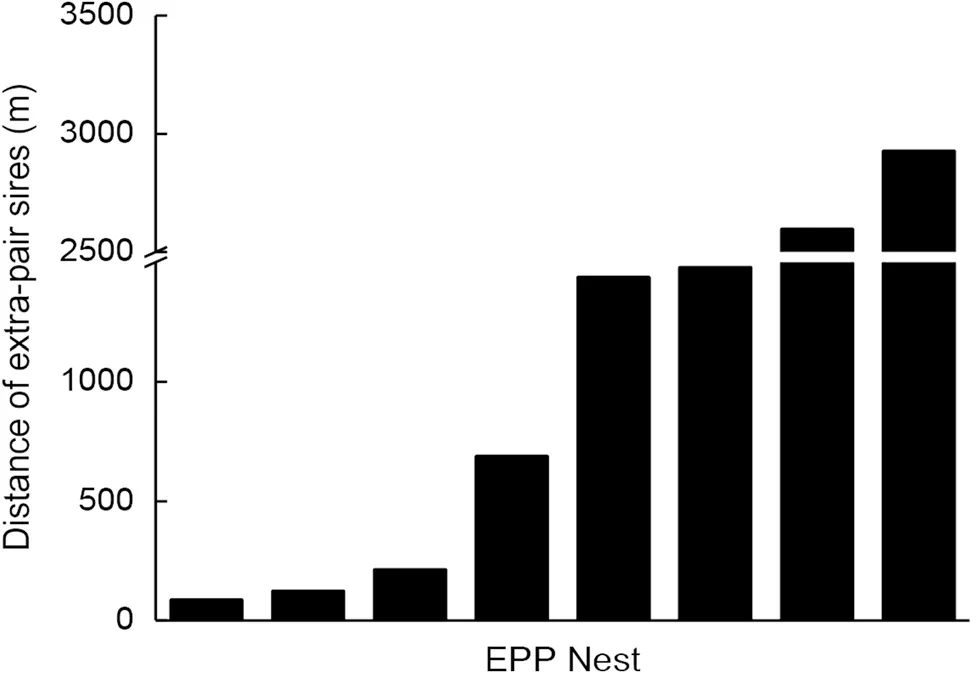
Fig. 1 Distances between the social nest of the extra-pair sire and the nest in which he sired young
Inbreeding may lead to gene incompatibly as a result of the loss of heterozygosity and the accumulation of deleterious homozygous recessive alleles (Charlesworth and Charlesworth 1987, 1999). Genetic compatibility hypothesis argues that females paired with genetically similar males would be more likely to have extra-pair copulations. In three shorebird species [Kentish Plovers(Charadrius alexandrinus
), Western Sandpipers (Calidris mauri
), and Common Sandpipers (Actitis hypoleuca
)],a strong positive correlation was found between the genetic similarity of social mates and EPP occurrence.For extra-pair copulation, females seek less related males than they do for social mates (Blomqvist et al. 2002).However, in Marsh Tits, genetic similarity hardly showed correlation with the EPP happening with no significant difference in genetic similarity between the social mates with and without EPP, or between the social and the extra-pair mates. This indicates that the genetic similarity of parents has no effect on the EPP happening in Marsh Tits. From the perspective of parents, genetic compatibility hypothesis is not supported by our findings.Heterozygosis plays important roles in EPP happening.The genetic compatibility hypothesis suggests that EPP increased offspring fitness due to increased heterozygosis (Brown 1997, 1999). Previous studies showed that in Reed Bunting (Emberiza schoeniclus
), the extra-pair young were more heterozygous, heavier, and larger than their maternal half-siblings, and the extra-pair sire carried more heterozygous alleles (Suter et al. 2007). In Tree Swallows (Tachycineta bicolor
), 47% of offspring were the results of extra-pair fertilization, and the extra pair offspring were more heterozygous than their maternal half-siblings (Stapleton et al. 2007). However, our studies found no difference in the heterozygosis between the social fathers and the extra-pair sires, or between the extra-pair offspring and their half-siblings. EPP brought no heterozygosis benefits to the extra-pair offspring.From the perspective of offspring, the genetic compatibility hypothesis was not supported either.In addition, hypotheses about the direct benefits of EPCs suggest that females may obtain direct benefits by EPCs to fertilization insurance (Schmoll and Kleven 2016), or increased foraging opportunities (Gray 1997).
The “good-genes” hypothesis suggests that females paired to relatively “poor-quality” males would seek EPCs from males of better quality to obtain indirect benefits. Prum(2017) suggested that the mate selection in nature can be irrational, unpredictable, and dynamic for some bird species. A future study on the direct benefits or other indirect benefits of EPP for female Marsh Tits will be worthwhile in order to explain why females engage in EPP in this passerine species.
Conclusions
In conclusion, the present study showed that Marsh Tits have an unexpectedly high rate of EPCs and female Marsh Tits might not obtain indirect genetic compatibility benefits through extra-pair paternity. Our study complements the understudied bird’s EPP in Asian area and contributes to the comprehensive insight of birds’ EPP behaviors.
Supplementary Information
The online version contains supplementary material available at https:// doi.org/ 10. 1186/ s40657- 021- 00304-2.
Additional file 1: Genetic variation data for Marsh Tits.
Acknowledgements
We would like to thank Liaoning Xianrendong National Nature Reserve for their support.
Authors’ contributions
JW conceived the experiments, analyzed the data, wrote the manuscript, and reviewed drafts of the paper; YW performed the experiments, analyzed the data; LZ conceived and designed the experiments, wrote the manuscript; YJ reviewed drafts of the paper; KL performed the experiments; DW conceived and designed the experiments, and reviewed drafts of the paper. All authors read and approved the final manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Grant No. 31872231) to DW, and the Youth Foundation of Liaoning University (Grant No. A251901012) to JW.
Declarations
Ethics approval and consent to participate
The experiments comply with the current laws of China. Fieldwork was carried out under the permission of Liaoning Xianrendong National Nature Reserve.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Received: 16 September 2021 Accepted: 18 November 2021
杂志排行

Avian Research的其它文章
- Taxonomic revision of the Savanna Nightjar (Caprimulgus affinis) complex based on vocalizations reveals three species
- Taxonomic status of grey-headed Yellow Wagtails breeding in western China
- The composition of mixed-species flocks of birds in and around Chitwan National Park,Nepal
- True grit: ingestion of small stone particles by hummingbirds in West Mexico
- Stopover behavior of Red-eyed Vireos (Vireo olivaceus) during fall migration on the coast of the Yucatan Peninsula
- Phylogeography and morphometric variation in the Cinnamon Hummingbird complex: Amazilia rutila (Aves: Trochilidae)