灰木莲花药结构和花粉发育特征
2022-01-05潘丽琴郝建徐建民粟宏林梁福江张青青姜清彬
潘丽琴,郝建,徐建民,粟宏林,梁福江,张青青,姜清彬*
(1.西南林业大学,云南 昆明 650204;2.中国林业科学研究院热带林业实验中心,广西 凭祥 532600;3.中国林业科学研究院热带林业研究所,广东 广州 510520)
木兰科(Magnoliaceae)自从有自然分类以来,一直被认为是被子植物中最原始的类群,对研究有花植物的起源和系统发育有重要价值[1]。木兰科全世界有15 属260 余种植物,是热带亚热带地区常绿阔叶林的重要组成树种,因各树种存在较多的性状交叉和形态变异,目前,在属的界限和种的确定上仍有较大的争议[2-3]。灰木莲(Manglietia coniferaDandy),原产越南及印度尼西亚[4],因其树形优美、花大清香、干形通直、生长快、木材纹理细致易加工,我国于1960 年从越南中北部引入,因其适生性强、生长快且材性优良而在我国推广[5]。目前主要在我国华南、福建及云南等地大规模种植,是一种优良的速生用材树种,在建筑、家具、胶合板及纸浆生产中具有广阔的市场前景[6]。
灰木莲在我国引种地的花期为每年1月下旬至4月下旬,开花时满树繁花,但自然结实率几乎为零,即使偶有结实,其聚合蓇葖果的绝大部分子房败育,每果仅见十余粒甚至几粒种子[7-8]。灰木莲在我国结实率极低的问题,严重制约其大规模发展。目前,有关灰木莲的研究主要集中于栽培繁育[5-6,9-11]、早期生长适应性[12]、材性[13]及生态效应[14-15]等方面,而对其有性生殖及种实败育机制了解甚少,为探明灰木莲引种条件下的种实败育原因,拟采用石蜡切片和扫描电镜技术,对灰木莲花药结构及其小孢子发育过程进行解剖观察,以确定是否存在雄性不育的异常现象。同时,与木兰科其他近缘种的胚胎学发育资料进行比较,为进一步确立灰木莲的分类学地位和木兰科系统发育的研究积累资料和提供佐证。
1 材料
本研究试验点设在中国林业科学研究院热带林业实验中心(以下简称中国林科院热林中心),位于广西壮族自治区凭祥市,属南亚热带季风气候区,年均气温21.5℃,≥ 10℃积温7 500℃,年降水量1 220~1 400 mm,降水集中期为4月中旬至8月末,年均日照约1 260 h。选取中国林科院热林中心的伏波实验场(FB)、白云实验场(BY)和中心树木园(ZX)栽培的灰木莲人工林为研究对象,各林分造林苗均为1.5 年生的实生苗,种源来自越南尚河,无病虫侵害,健康状况良好,均有零星结实,基本情况见表1。
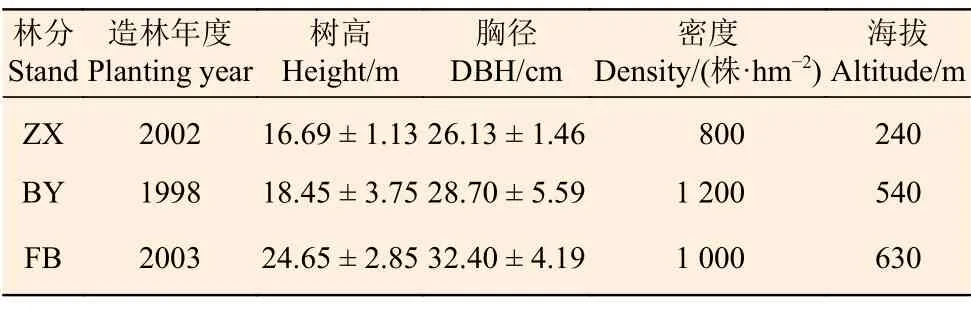
表1 观测林分基本情况Table 1 General information of the observation stands
2 方法
材料采集自中国林科院热林中心灰木莲试验林,选择8~12 年生的优良单株。2018 年10月至2020 年10月连续2 a,每隔3~6 d,采集不同发育时期花芽,体视显微镜解剖观测花药形态、结构及数量特征;剥离出花药保存于FAA 固定液(V70%乙醇:V 甲醛:V 冰乙酸=18:1:1)抽真空固定,50%乙醇润洗,经乙醇梯度脱水,浸蜡包埋后,旋转切片机进行连续切片,切片厚度6~8 μm,中性树胶封片,OLYMPUS 光学显微镜和OLYMPUS PM-10AD 成像系统观察拍照。
取新鲜花药,轻轻划破花药表皮,解剖针挑出花粉,将花药和花粉粘贴于双面导电胶布上,置于金属样品台进行喷金,在扫描电镜下观察并拍照[16]。实验数据使用Excel 软件进行统计分析。
3 结果与分析
3.1 雄蕊群数量结构及形态特征
灰木莲雄蕊群具花药平均98 枚(n=30,sd=21),花药长平均7.4 mm (n=30,sd=1.2),花药宽平均2.1 mm(n=30,sd=0.2)。雄蕊群的花药独立分离、辐射螺旋状排列于花轴的基部(图1-1),花粉囊呈丝管状平行排列于花药内侧或侧向面的两边,无花丝,尖端伸出约1~1.5 mm 的药隔(图1-3)。成熟花药具4~6 个花粉囊,多数为4 个花粉囊,偶见6 个花粉囊(图1-2)。

图1 花药结构Fig.1 The structure of anthers
具有活力的花粉粒两侧对称,异极,极面观为椭圆形或长圆形,赤道面观为舟形,具远极单沟,沟直达端部,闭合或不闭合,极轴约25.5 μm,赤道轴约62.3 μm,花粉壁光滑,极轴面具稀疏穴状纹饰(图1-4~5),失活花粉粒缩为球形,花粉壁皱缩(图1-6)。
3.2 花药壁的发育
灰木莲雄蕊原基约12月下旬至1月初开始分化,内侧花粉囊较外侧的分化发育早(图2-1)。最初的花药原基只有一层外表皮细胞和表皮内部分生组织,1月中旬,随着内部分生组织细胞的分化,在花药四个角隅处分化出孢原细胞(图2-2);孢原细胞经平周分裂形成初生壁细胞和内层初生造孢细胞(图2-3),初生壁细胞分裂后形成3~4 层花药壁细胞(图2-4);当小孢子母细胞形成时,花药壁发育完成,此时的花药壁自内向外分别为绒毡层1~2 层、中层1 层、药室内壁2 层、表皮细胞1 层(图2-5);2月上旬,小孢子母细胞进入减数分裂,中层细胞逐渐解体、退化或变成长条状或残迹,绒毡层在形成四分体时也开始部分原位变形,但中层细胞较绒毡层细胞提前被吸收(图2-6~7),绒毡层属较原始的腺质型;小孢子从四分体释放后发生变形,但随着细胞增殖扩大,残余的绒毡层和中层细胞重新恢复并被挤压为扁平状(图2-8~9);变形小孢子发育后期多数能发育正常,此时中层和绒毡层再次解体为其发育提供营养,花粉成熟时,药室内壁发育为纤维层1 层,中层和绒毡层被吸收(图2-10~11);3月初,小孢子发育成熟,花粉囊纤维层径向加厚,同侧花粉囊连接处细胞壁较薄,受机械牵拉,花粉从同侧两花粉囊连接处散出(图2-12)。
3.3 小孢子的发生及雄配子体的形成
小孢子母细胞在减数分裂前,与花药壁细胞有明显区别,细胞体积最大,核大,胞质浓厚,为行使小孢子母细胞功能储备物质(图3-1)。第1 次减数分裂末期,小孢子细胞质从外向内发生缢裂,形成二分体,但未见细胞新壁形成(图3-2);进入第2 次减数分裂时,二分体发生同时或不同时缢裂形成四分体,四个子细胞被共同的胼胝质包围(图3-3);随着胼胝质溶解,小孢子彼此分开并形成细胞新壁(图3-4)。四分体的排列方式以左右对称型和四面体型为主(图3-5~7),偶见直列型(图3-8),小孢子母细胞胞质分裂方式为修饰性同时型和连续型兼有。
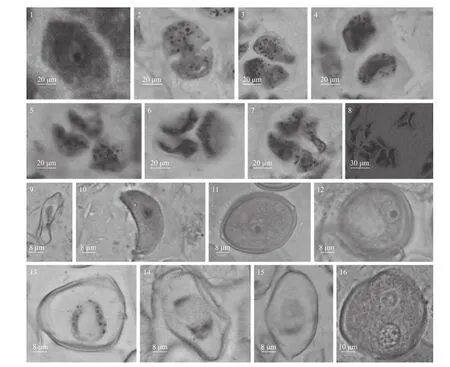
图3 小孢子的发生及雄配体子的形成Fig.3 The occurrence of microspore and the formation of male gametophyte
从四分体释放后的小孢子形态发生不同程度的变形,出现扭曲、收缩或变为肾型,细胞壁薄,细胞质液泡化(图3-9~10);变形小孢子细胞壁进一步扩张、加厚,形成形状规则、无液泡的单核小孢子,胞质变浓且分布均匀(图3-11);随后小孢子细胞质出现液泡化形成一个中央大液泡,细胞核被挤到细胞的边缘,此时小孢子为单核靠边期(图3-12);经历单核靠边期后,小孢子核物质不断积累,随即发生1 次不均等的有丝分裂,分裂过程中可观察到染色体从赤道面被拉向两极,将细胞质分成不均等的两部分而形成具营养核和生殖核的二核小孢子(图3-13~15);二核小孢子发育后期,大液泡消失,大部分细胞质和营养核形成营养细胞,小部分细胞质和生殖核形成生殖细胞,营养细胞占据花粉的绝大部分体积(图3-16),成熟小孢子形成。
3.4 花药发育过程中的败育
通过对灰木莲花药发育的解剖观察,在其发育过程中主要存在以下异常现象:(1)小孢子母细胞进入减数分裂到四分体形成时,绒毡层和中层细胞提前解体(图4-1),仅剩少量残余,不能为小孢子后期发育提供营养,经切片数量统计,此异常现象比例约占14%。(2)花粉壁收缩变形,形成心型花粉嚢,从四分体分离的小孢子因过度扭曲变形不能再正常发育(图4-2~3),因变形而败育的小孢子约占9%。(3)在小孢子母细胞分裂时期,小孢子母细胞因退化、发育异常或消失,空心花粉嚢的比例约占10%(图4-4)。
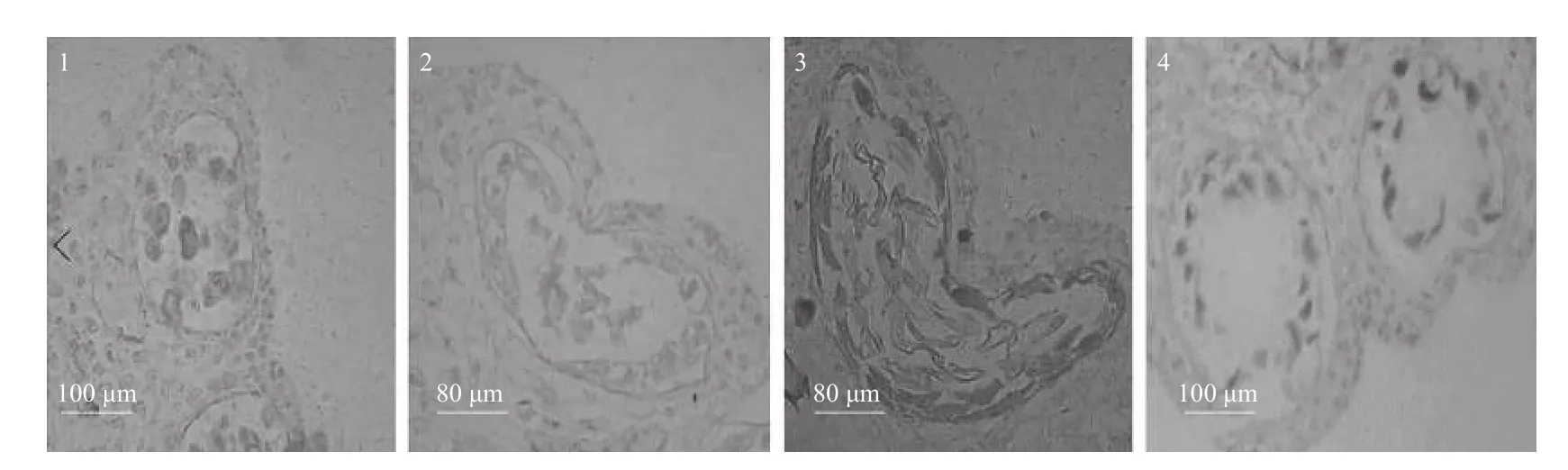
图4 雄配子体发育过程中的异常现象Fig.4 The abnormal phenomenon in the process of male gametophyte development
4 讨论
灰木莲的雄蕊含花药平均98 枚,4~6 个小孢子囊呈线型或蝴蝶型排列于扁平花药的向心面或侧向面,其花药开裂同时具侧向开裂和内向开裂2 种方式。在较为原始的双子叶植物中,整个雄蕊为具3 条维管束的扁平叶状,花粉囊则着生于近轴面的中脉与侧脉之间。据统计,进化后95%的被子植物具单脉雄蕊,这被认为是叶状雄蕊在宽度和脉迹减少的进化过程中的产物[17]。已报道的木兰科花粉嚢数均为4 个,且数量较稳定[4,18],本研究观察到灰木莲花药横切结构具4~6 个花粉嚢,其花粉嚢数目具一定的可变性,这在木兰科其他植物中尚未见报道。通常认为,花部数目多数和不定数是原始性状,少数和定数是进化性状[19],这表明灰木莲的花粉嚢数量在系统演化中保留了更为原始的性状。在植物的系统发育中,不同器官的演化速度是不同的,因而造成同一分类群在具有原始特征的同时,又兼具有许多进化特征,被称为异步进化或镶嵌进化现象,这在原始的有花科植物中表现得最明显[20]。同已报道同属的红花木莲(M.insignis)、大果木莲(M.grandis)和毛果木莲(M.hebecarpa)相比,就花药特征而言,灰木莲花药数目较少,属较进化类型,但其花粉嚢数目却比红花木莲、大果木莲和毛果木莲的原始[21-22]。
灰木莲的花药壁由表皮、药室内壁、中层和绒毡层组成,中层和绒毡层仅由1~2 层细胞组成,腺质绒毡层,成熟花粉粒为二细胞型,这和已报道其他木莲属的花药发育特征基本相同[21,23-26]。花粉壁光滑,极轴面具极稀疏穴状纹饰,为木莲属中的最原始类型,花粉极轴约25.5 μm,属木莲属中花粉极轴的最短区间,花粉赤道轴约62.3 μm,处于木莲属花粉赤道轴范围的中等区间[27]。在灰木莲的小孢子发育过程中,小孢子母细胞减数分裂具连续型和同时型2 种方式,连续型分离的小孢子以左右对称型和直线型排列方式为主,同时型分裂出的小孢子为四面体型排列,而其他木兰科树种一般仅具一种同时型的胞质分裂方式[21,24-25,28]。灰木莲小孢子在四分体时发生收缩变形,刚释放的小孢子呈扭曲、梭状或骨形,这与红花木莲、西畴含笑(Michelia coriaceaChang et B.L.Chen)和凹叶厚朴(Magnolia officinalisRehd.et Wils)的四分体变形特征相似[21,25,28-30],西畴含笑和凹叶厚朴的大部分小孢子因收缩过度而不能再进一步正常发育,但在灰木莲和红花木莲中,大多数小孢子在发生不规则的收缩变形的同时进行液泡化过程,因此,大部分小孢子仍能正常发育形成花粉粒。潘跃芝等认为,这种收缩现象可能是为了将细胞质挤到细胞的边缘,以有利于细胞核有丝分裂的进行[21,25],此现象发生的机理和作用如何?是否为更进化的类型?需进一步系统的研究。
木兰科作为原始被子植物的现存类群,花粉败育与结籽率低是木兰科濒危植物生殖失败的原因之一[31],对其濒危机制研究发现,许多属在花粉发育中存在不同环节的败育现象,如红花木莲因次生造孢细胞、花粉母细胞部分败育和二分体败育,其花粉萌发率仅为14.7%;毛果木莲存在花粉量少,且因原生质提前解体形成无细胞核不规则的败育花粉率为29%[31];香港木兰(M.championii)的小孢子败育率较高,四分体到单核花粉形成期出现异常发育[32-33];凹叶厚朴在小孢子形成时,母细胞胶合在一起不能发生减数分裂,或是分离出的小孢子因过度收缩变形不能形成正常花粉粒,或是部分花粉囊在二核花粉形成时,花药壁的绒毡层和中层细胞仍未退化完全而缺乏继续发育的营养物质[28];观光木(Tsoongiodendron odorumChun)的花粉发育败育,主要发生于从四分体分离出小孢子时,同侧花粉囊有50%的四分体退化,35%的小孢子因过度收缩发育为败育的小孢子[34]。本研究中,灰木莲小孢子发育的败育主要发生于四分体时期,绒毡层和中层细胞提前解体,花粉囊萎缩,刚释放的小孢子呈扭曲、梭状或骨形而致使部分小孢子不能发育为正常花粉粒的败育率为33%。
生殖败育是植物引种栽培条件下常发生的问题,其原因是多方面的。灰木莲在自然状态结实率几乎为零,但对其异株异花授粉的结实率可达57.67%[35],本研究所观察到的灰木莲花粉败育率为33%,这虽然在一定程度上减少了其授粉机率,但若按其花粉胚珠比1 146 来计算,每胚珠获得的花粉量概率依然很大。因此,花粉败育对其结籽率较低的影响是有限的,栽培区内是否缺少足够量的有效传粉昆虫,有待今后进一步研究证实。
5 结论
灰木莲花药结构和发育特征与其他木莲属相似,但在花粉囊结构、胞质分裂方式和小孢子四分体排列方式表现不同,小孢子发育过程的异常现象主要发生于四分体时期,为引起灰木莲花粉败育和结实率低的原因之一。
